
不同糖对拟南芥种子萌发和sscd1突变体细胞死亡的影响
2016-10-10 08:40支添添任春梅
安徽农业科学 2016年23期
黄 弈, 支添添, 任春梅,2*
(1.湖南农业大学生物科学技术学院,湖南长沙 410128;2.作物基因工程湖南省重点实验室,湖南长沙 410128)
不同糖对拟南芥种子萌发和sscd1突变体细胞死亡的影响
黄 弈1, 支添添1, 任春梅1,2*
(1.湖南农业大学生物科学技术学院,湖南长沙 410128;2.作物基因工程湖南省重点实验室,湖南长沙 410128)
[目的]比较不同糖对拟南芥种子萌发以及sscd1突变体细胞死亡的影响。[方法]以拟南芥Col-0野生型和sscd1突变体为材料,在培养基中外源添加蔗糖、葡萄糖、果糖和麦芽糖,比较不同糖对拟南芥种子萌发和sscd1突变体细胞死亡的影响。[结果]在120 mmol/L浓度范围内,蔗糖、葡萄糖和果糖对拟南芥种子的萌发无显著影响,而麦芽糖对种子萌发有延迟作用,且糖浓度越高延迟作用越显著。120 mmol/L不同糖对sscd1突变体细胞死亡的抑制效果有显著差异,抑制效果由高到低依次为麦芽糖、蔗糖、葡萄糖和果糖。[结论]该研究为深入研究糖代谢途径调控酪氨酸降解途径的机理提供理论依据。
糖;拟南芥;sscd1突变体;细胞死亡
笔者在前期研究中通过EMS诱导法分离了一种在短日照条件下(8 h光照/16 h黑暗)发生细胞死亡的拟南芥突变体sscd1(short-day sensitive cell death1)[1]。这种突变体在短日照条件下的MS培养基上萌芽,叶片会出现先萎蔫后白化的现象。采用台盼蓝染色的方法研究萎蔫及白化苗的死亡情况,结果证实,突变体sscd1萎蔫处发生细胞死亡[2]。通过基因定位和序列分析发现sscd1基因编码酪氨酸降解途径的最后一个酶——延胡索酰乙酰乙酸酶(fumarylacetoacetate hydrolase,FAH)。酪氨酸降解途径在动物中是必不可少的, FAH缺失会导致动物先天性致死。然而,酪氨酸降解途径在植物中的作用仍有待阐明[3-6]。在培养基中外源添加糖之后,sscd1突变体细胞死亡的表型有所恢复[7]。糖类是植物体内重要的碳源和能源,它不仅参与能量代谢,还调控植物的生长发育和生理过程,包括在种子萌发、幼苗生根、胚胎发育和衰老等发育过程中都起到了重要作用[8-9]。笔者尝试添加同一浓度不同糖如蔗糖、葡萄糖、果糖和麦芽糖,观察拟南芥种子的萌发以及sscd1突变体细胞死亡情况,比较不同糖对拟南芥种子萌发以及sscd1突变体细胞死亡的影响,为深入研究糖代谢途径调控酪氨酸降解途径的机理提供理论依据。
1 材料与方法
1.1试验材料拟南芥(Arabidopsisthaliana)野生型Col-0和sscd1突变体,由作物基因工程湖南省重点实验室植物信号转导课题组保存。
1.2试剂与仪器蔗糖、葡萄糖、果糖、麦芽糖和琼脂粉购自国药公司。智能人工气候箱:PRX-450B。
1.3培养方法用消毒液(20% bleach+0.1% Triton100)将Col-0野生型和sscd1突变体种子浸泡8~10 min后,用无菌水清洗3~4次,分别铺种在含有0、30、60、120 mmol/L蔗糖(Suc)、葡萄糖(Glc)、果糖(Fru)、麦芽糖(Mal)的MS固体培养基上,4 ℃下春化3 d后转入8 h光照/16 h黑暗的人工气候箱,培养温度(23±2)℃,光照强度80~120 μmol/(m2·s),相对湿度65%。
1.4测定指标与方法①以种子露出白色胚芽为萌发标志,对2、4、6 d的萌发情况进行统计。②以叶片完全白化为细胞死亡标志,对12 d的细胞死亡情况进行统计。
2 结果与分析
2.1不同糖对拟南芥种子萌发的影响以MS培养基为对照,分别在外源添加0、30、60和120 mmol/L的蔗糖、葡萄糖、果糖和麦芽糖的培养基上铺种拟南芥Col-0野生型和sscd1突变体,转入短日照条件下生长,分别对2、4、6 d的种子萌发率进行统计,结果见图1。由图1可知,在外源添加蔗糖、葡萄糖和果糖的培养基中,拟南芥Col-0野生型和sscd1突变体种子在第2天几乎完全萌发,且在120 mmol/L糖浓度范围内的萌发率差别不大;而在30 mmol/L麦芽糖作用下,相对于其他3种糖,2 d时种子的萌发率有一定程度的降低,且在120 mmol/L浓度范围内,麦芽糖浓度越高,萌发率越低,但生长6 d后,种子几乎完全萌发。这表明在120 mmol/L浓度范围内,蔗糖、葡萄糖和果糖对拟南芥种子的萌发并无显著影响;而麦芽糖对拟南芥Col-0野生型和sscd1突变体种子的萌发均有延迟作用,且随着麦芽糖浓度的升高,这种延迟作用更加显著。
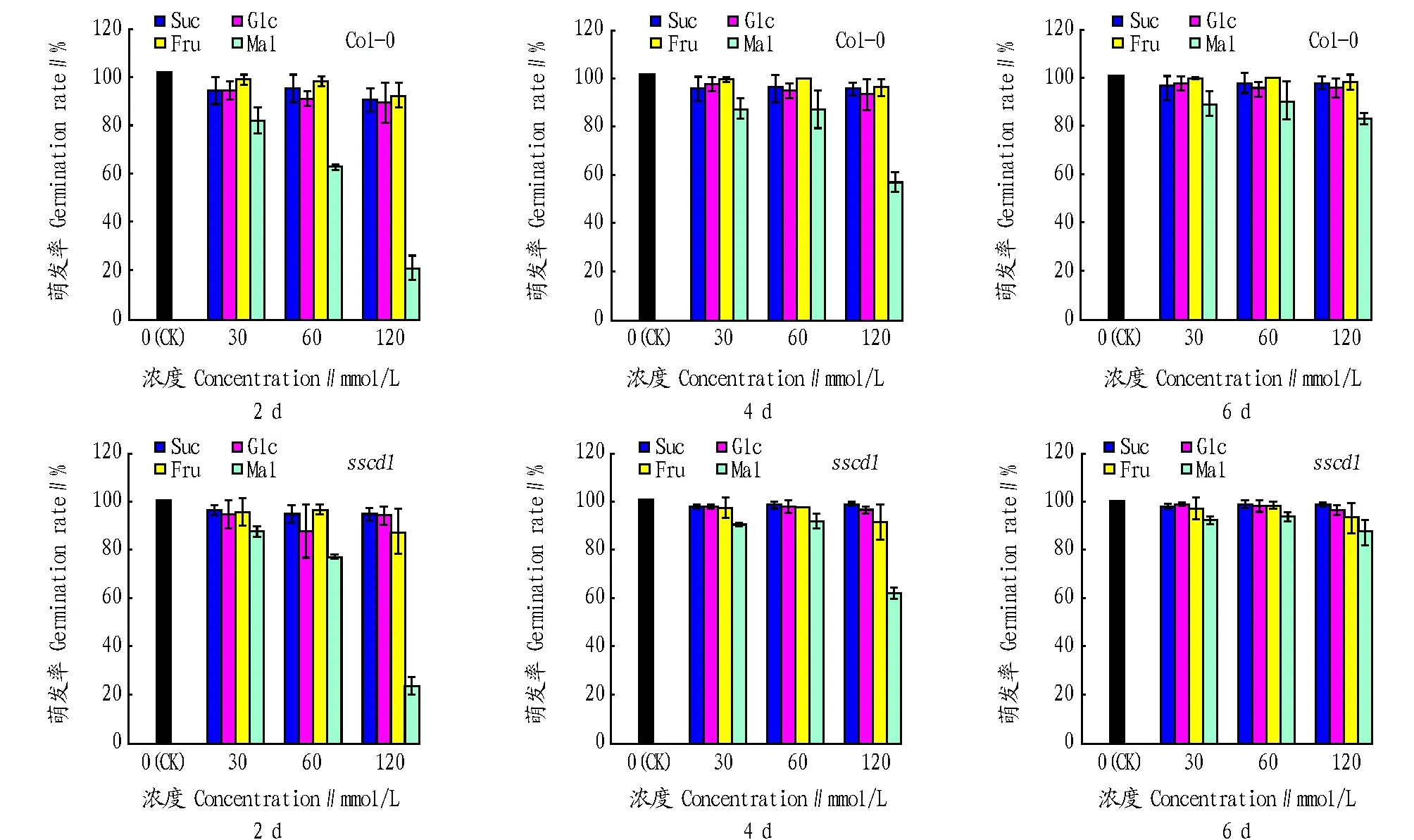
图1 外源添加不同糖的拟南芥种子萌发情况Fig.1 Germination situation of Arabidopsis seedlings with different sugars
2.2不同糖对sscd1突变体细胞死亡的影响在前期研究中发现,在培养基中外源添加糖后,拟南芥sscd1突变体细胞死亡的表型有所恢复[7]。为了分析不同糖对拟南芥sscd1突变体细胞死亡的影响,分别在含有30和120 mmol/L蔗糖、葡萄糖、果糖和麦芽糖的培养基上铺种拟南芥Col-0野生型和sscd1突变体,转入短日照条件下生长12 d后对细胞死亡表型进行观察,结果见图2。由图2可知,添加30 mmol/L不同糖的培养基上,拟南芥sscd1突变体幼苗大部分仍出现细胞死亡的表型;而在添加120 mmol/L不同糖的培养基上,拟南芥sscd1突变体的细胞死亡表型有不同程度的恢复。
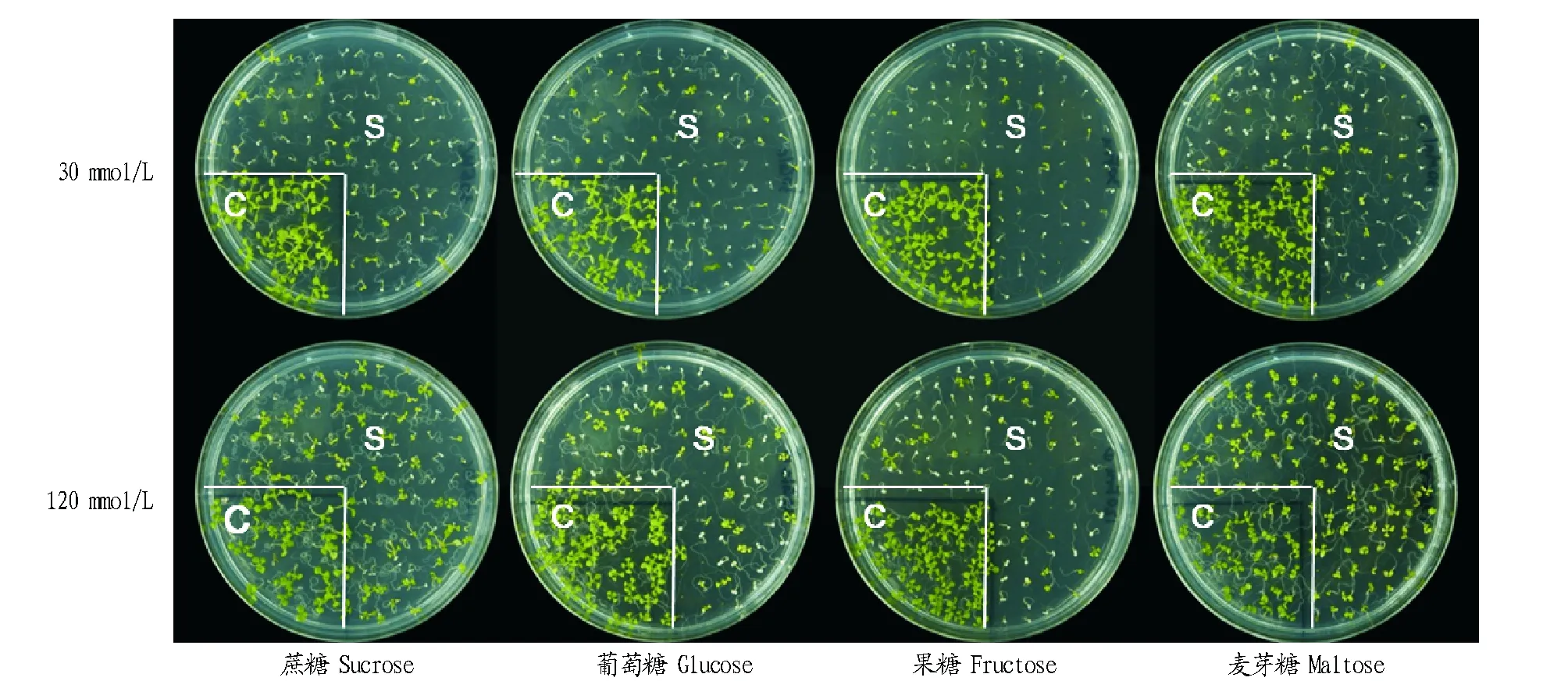
注:C.Col-0野生型;S.sscd1突变体。Note:C.Col-0 wild type; S.sscd1 mutant.图2 外源添加不同糖的拟南芥幼苗细胞死亡表型Fig.2 Cell death phenotype in Arabidopsis seedlings with different sugars
为了比较不同糖抑制拟南芥sscd1突变体细胞死亡的差异,进一步对不同浓度糖作用下拟南芥sscd1突变体的细胞死亡率进行统计,结果见图3。由图3可知,外源添加30 mmol/L葡萄糖和果糖的培养基上,sscd1突变体的细胞死亡率约为91%,与无糖对照相比无明显差别;外源添加30 mmol/L的蔗糖和麦芽糖的培养基上,sscd1突变体细胞死亡率约为83%,与葡萄糖和果糖相比有一定程度的降低,但不显著。在外源添加120 mmol/L糖的培养基上,相对于无糖以及添加30 mmol/L糖的培养基,sscd1突变体的细胞死亡率均显著降低,且不同糖对sscd1突变体细胞死亡的抑制效果有明显差异,其中,麦芽糖的抑制效果最好,其次是蔗糖、葡萄糖和果糖。这表明30 mmol/L不同糖对sscd1突变体的细胞死亡有一定程度的抑制,但抑制效果无明显差别。120 mmol/L糖对拟南芥sscd1突变体细胞死亡的抑制效果显著,且不同糖对sscd1突变体细胞死亡的抑制效果有明显差异,抑制效果由高到低依次为麦芽糖、蔗糖、葡萄糖和果糖。
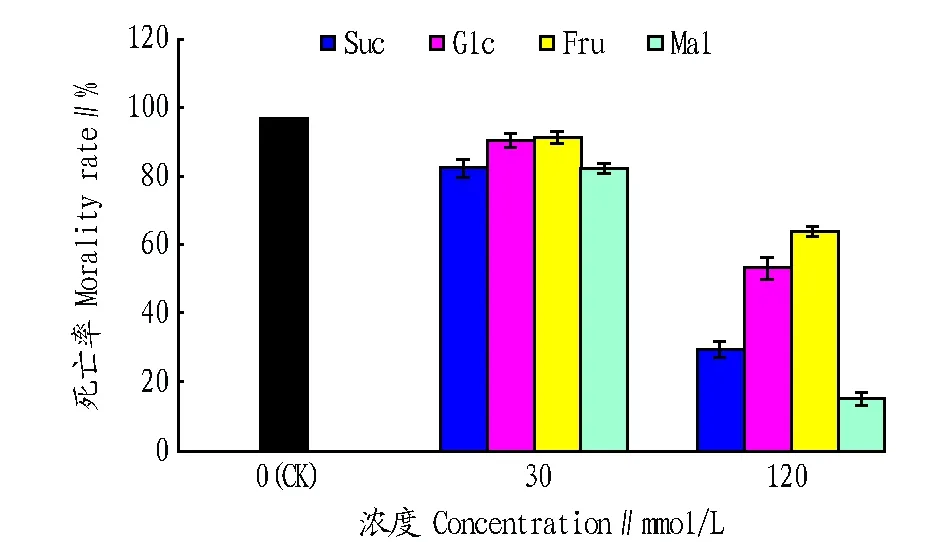
图3 外源添加不同糖的拟南芥sscd1突变体细胞死亡率Fig.3 Morality rate of Arabidopsis sscd1 mutant cells with adding different sugars
3 结论与讨论
该试验通过观察外源添加不同糖的培养基中拟南芥Col-0野生型和sscd1突变体的萌发和sscd1突变体细胞死亡情况,比较不同糖对拟南芥种子萌发以及sscd1突变体细胞死亡的影响。结果表明,在120 mmol/L浓度范围内,蔗糖、葡萄糖和果糖对拟南芥种子萌发无显著影响,而麦芽糖对Col-0野生型以及sscd1突变体种子的萌发均有延迟作用,且糖浓度越高,延迟作用越显著。其次,在培养基中外源添加120 mmol/L 糖时,不同糖对拟南芥sscd1突变体细胞死亡的抑制效果有显著差异,其中,麦芽糖的抑制效果最好,其次是蔗糖、葡萄糖和果糖。
前期研究中通过浓度梯度设计确定了1%和4%作为低浓度和高浓度的代表,该试验通过换算选取30和120 mmol/L作为试验浓度。该试验中麦芽糖对拟南芥种子萌发具有延迟作用,但生长一定时间后,大部分种子仍能萌发,但其对萌发后幼苗生长的影响还有待进一步研究。不同糖对拟南芥sscd1突变体细胞死亡的抑制效果有所不同,可能是与不同糖类作为信号分子参与调控植物体内不同的生理过程有关,而麦芽糖与蔗糖的抑制效果明显优于葡萄糖和果糖,推测可能是由于麦芽糖和蔗糖是二糖,其要水解成单糖才能参与调控植物的生长和发育,具体机制尚不明确。目前对于糖类的研究已取得了一定进展,而关于糖抑制拟南芥sscd1突变体细胞死亡的分子机制以及糖代谢途径与酪氨酸降解途径的关系还有待于进一步探讨。
[1] 韩成云.拟南芥短日照依赖型模似病斑SDL1基因的分离和功能分析[D].长沙:湖南农业大学,2012.
[2] 支添添,周舟,韩成云,等.台盼蓝染色鉴定拟南芥sdl1突变体的细胞死亡[J].作物研究,2013,27(3):217-218.
[3] APONTE J L, SEGA G A, HAUSER L J, et al.Point mutations in the murine fumarylacetoacetate hydrolase gene:Animal models for the human genetic disorder hereditary tyrosinemia type 1[J].Proc Natl Acad Sci USA,2001, 98:641-645.
[4] DIXON D P,EDWARDS R.Enzymes of tyrosine catabolism inArabidopsisthaliana[J].Plant Sci, 2006, 171:360-366.
[5] GROMPE M, AL-DHALIMY M, FINEGOLD M, et al.Loss of fumarylacetoacetate hydrolase is responsible for the neonatal hepatic dysfunction phenotype of lethal albino mice[J].Genes Dev, 1993, 7:2298-2307.
[6] HAN C Y, REN C M, ZHI T T, et al.Disruption of fumarylacetoacetate hydrolase causes spontaneous cell death under short-day conditions inArabidopsis[J].Plant physiology, 2013, 162(4):1956-1964.
[7] 支添添.拟南芥sscd1突变体细胞死亡机理的研究[D].长沙:湖南农业大学,2014.
[8] LEON P,SHEEN J.Sugar hormone connection[J].Plant Sci, 2003, 8(3):110-116.
[9] ROLLAND F, BAENA-GONZALEZ E J, SHEEN J.Sugar sensing and signaling in plants:Conserved and novel mechanisms[J].Plant Biol, 2006, 57:675-709.
Effects of Different Sugars onArabidopsisSeed Germination andsscd1 Mutant Cell Death
HUANG Yi1, ZHI Tian-tian1, REN Chun-mei1,2*
(1.College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha, Hunan 410128; 2.Key Laboratory of Crop Gene Engineering of Hunan Province, Changsha, Hunan 410128)
[Objective] To compare the effects of different sugars onArabidopsisseed germination andsscd1 mutant cell death.[Method] With wide type (Col-0) and the mutantsscd1 as research materials, effects of different sugars onArabidopsisseed germination andsscd1 cell death were compared by adding sucrose, glucose, fructose and maltose in the culture medium.[Result] At the 120 mol/L concentration range, sucrose, glucose and fructose had no significant effects onArabidopsisseed germination, but maltose delayed seed germination.And high sugar concentration had more significant delaying effects.Different sugars at 120 mol/L concentration showed significant differences in the inhibitory effects on sscd1 cell death, and the effects from high to low were in the order of maltose, sucrose, glucose and fructose.[Conclusion] This research provides important foundation for the in-depth mechanism research on regulating of tyrosine degradation pathway by glucose metabolic pathway.
Sugar;Arabidopsis;sscd1 mutant; Cell death
国家自然科学基金项目(30671121)。
黄弈(1990- ),男,湖南株洲人,硕士研究生,研究方向:植物分子遗传学。*通讯作者,教授,博士,博士生导师,从事植物分子遗传学研究。
2016-06-23
S 188;Q 94
A
0517-6611(2016)23-093-03
猜你喜欢
保健与生活(2022年13期)2022-07-06
今日农业(2020年24期)2020-12-15
科学(2020年4期)2020-11-26
阅读(低年级)(2019年11期)2019-12-27
保健与生活(2017年5期)2017-04-14
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
中西医结合心脑血管病杂志(2016年20期)2016-03-01
上海农业学报(2016年5期)2016-02-10
天津医科大学学报(2015年2期)2015-12-22
