
基于双单倍体群体的玉米株高和穗位高QTL分析
2016-09-26 02:09:52李浩川杨继伟曲彦志张朝林刘宗华
河南农业大学学报 2016年2期
李浩川,陈 琼,杨继伟,曲彦志,张 慧,张朝林,李 彦,贾 玺,刘宗华
(1.河南农业大学农学院,河南 郑州 450002;2.灵宝市农业局,河南 灵宝 472500)
基于双单倍体群体的玉米株高和穗位高QTL分析
李浩川1,陈 琼1,杨继伟1,曲彦志1,张 慧1,张朝林1,李 彦1,贾 玺2,刘宗华1
(1.河南农业大学农学院,河南 郑州 450002;2.灵宝市农业局,河南 灵宝 472500)
为了进一步探明玉米株高和穗位高的遗传基础,利用郑单958为基础材料构建的包含161个双单倍体(Doubled haploid, DH)的群体,分别在长葛、淇县进行田间试验,采用复合区间作图法对植株性状进行数量性状位点(Quantitative trait locus, QTL)分析。分别检测到7,11和6个株高、穗位高以及穗位系数(穗位高/株高)QTLs,贡献率变幅分别为6.77%~21.31%,5.09%~17.12%和6.89%~17.78%,其中qph-3-3, qph-2-10, qeh-5-9以及qeh/qph-3-1为主效QTLs;株高和穗位高有4对共同标记区间的QTLs,穗位系数与穗位高有3对共同标记区间的QTLs,一定程度上揭示了株高和穗位高的遗传相关性;株高和穗位高的广义遗传力高达88.52%和93.78%,表明株高和穗位高遗传稳定性强,受环境影响较小,是早代选择的重要株型指标。
玉米;双单倍体;株高;穗位高;数量性状位点
倒伏是制约玉米产量的重要因素之一,全球每年因倒伏使玉米产量损失5%~20%[1]。黄淮海夏玉米区在抽雄前后易受季风气候影响,因风灾导致产量损失严重[2]。近年来,随着机收面积的扩大和机械化程度的提高,对玉米品种抗倒伏性要求越来越高。国内外许多学者先后开展了玉米倒伏问题的相关研究。ALBRECHT等[3]研究认为,茎秆抗倒伏能力随着茎秆变粗、茎皮加厚而提高,随着节间延长、植株增高而下降;茎粗对植株的抗倒伏能力影响最大,基部节间短而粗的则抗倒伏能力强。王永学等[4]利用相关和通径分析研究表明,倒伏率与茎秆强度、茎粗呈负相关,与株高、穗位高、节间长、叶夹角呈正相关,入土的次生根数对倒伏率影响最大。付志远等[5]的研究结果表明,增加穗上节间数、适当降低穗位高,可以提高玉米植株的抗倒伏性,认为穗位系数即穗位高与株高的比值可以作为衡量植株抗倒伏性状的一个重要指标。大量研究表明,倒伏性与种植的密度、茎秆拉力、茎秆抗穿刺力、茎粗、株高、穗位高等都具有极显著相关性[6-9],尤其株高和穗位高通常被认为是影响抗倒伏性的最重要的因素。尽管RADU等[8]利用双列杂交试验对玉米抗倒伏性状进行遗传分析,认为抗倒伏遗传变异占表型变异的79%,但在表型变异中,加性效应仅占1.6%,上位效应占55%。然而,株高和穗位高是典型数量性状,有关其分子遗传机制迄今仍不十分清楚。随着分子生物学技术的发展和应用,众多学者分别利用不同分离群体,检测到许多有关玉米株高和穗位高的QTL[9-11]。如李清超等[9]利用11个RIL群体,基于单环境、联合环境的QTL分析,检测到21个株高主效QTL和15个穗位高主效QTL;郑克志等[10]对利用T 319与9406为亲本构建的242个RIL群体进行分析,检测到6个株高QTL和2个穗位高QTL。然而,这些研究由于各自所用材料和群体不同,结果不尽一致。双单倍体(Doubled haploid, DH)系的应用不仅能加速育种进程,缩短育种周期,而且DH系不存在基因的显性效应,可对数量性状基因的加性和上位性效应进行更加准确的研究,同时DH系可以在多个环境或季节进行重复试验,是研究基因型与环境互作的理想材料。为了进一步明确株高、穗位高的分子机制,本研究以DH群体为研究材料,结合简单序列重复(Simple sequence repeat, SSR)分子标记技术对玉米株高、穗位高以及穗位高/株高进行QTL分析,以期为抗倒伏品种选育提供理论依据。
1 材料与方法
1.1试验材料
本研究以优良玉米杂交种郑单958为母本,以农大高诱5号为父本,进行杂交诱导产生单倍体,通过化学加倍和繁殖形成了161个DH系,以DH系构成的群体及亲本自交系郑58和昌7-2为试验材料。2014年夏在长葛和淇县种植,田间采用完全随机区组设计,单行区,2次重复,小区行长4 m,行距0.65 m,株距0.22 m,种植密度为67 500株·hm-2。前茬为小麦,土壤肥力均匀,田间管理同大田。

1.2DNA的提取和SSR标记的分析
亲本及DH群体的DNA提取采用吴向远等[13]改良的SLS方法。SSR引物选自Maize GDB数据库,由上海生工公司合成。PCR反应体系:每11 μL的体系中含ddH2O 6 μL,10×buffer 1.5 μL,dNTP 0.3 μL,Tap酶0.2 μL,引物2 μL,DNA 1 μL。
利用SSR标记构建连锁图谱,从823个标记中筛选出亲本间带型差异明显的107个多态性标记,覆盖玉米基因组全长的2 497 cM,标记间平均距离23 cM。运用Mapmaker/QTL 1.2软件对株高、穗位以及穗位系数(穗位高/株高)进行QTLs定位分析,判断QTL存在的阈值LOD≥2.5,QTL命名参照McCOUCH等[14]1997年提出的方法。
2 结果与分析
2.1郑单958的亲本及其DH系群体的株高和穗位高变异分析
从表1看出,父本自交系昌7-2的株高和穗位高明显高于母本自交系郑58,其杂交种郑单958的株高和穗位高表现明显的超亲优势;DH群体中各家系的株高和穗位高变异广泛,但均值仍然偏高亲或超高亲。从DH群体的峰度和偏度来看,除株高的峰度值大于1外,其偏度和穗位高的峰度和偏度绝对值均小于1,表明该群体趋于正态分布。
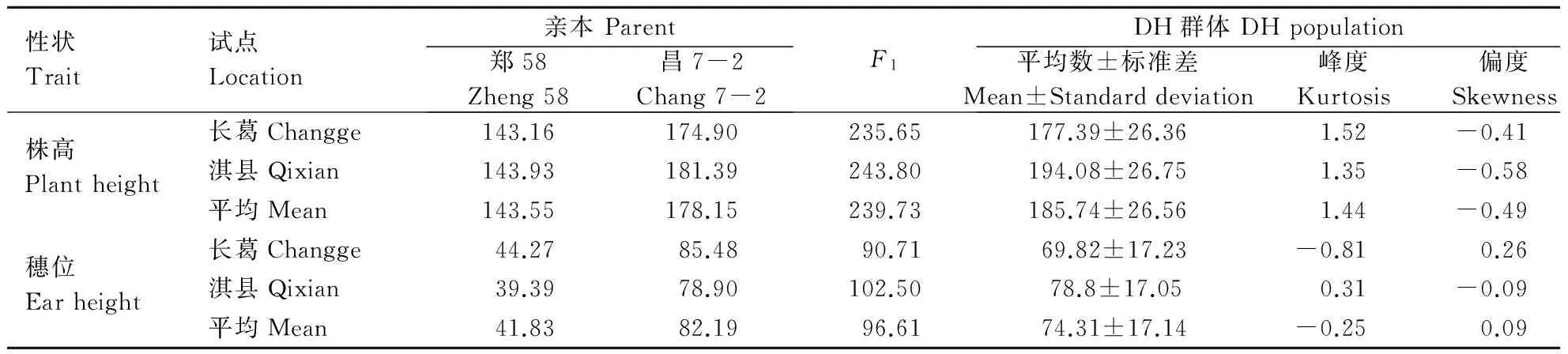
表1 郑单958 DH群体及亲本的株高和穗位高变异分析 Table 1 Varation of plant height and ear height performance for parents and their DH population from Zhengdan 958 cm
2.2郑单958DH群体株高与穗位高的相关性及其方差分析
相关分析表明,来自于郑单958 DH群体的株高与穗位高在长葛和淇县均达到了极显著正相关,相关系数分别为0.81和0.79。
联合方差分析结果(表2)表明, 试点和DH家系各自之间在株高、穗位高方面均存在极显著差异,但试点与DH家系互作差异不显著,表明各家系之间遗传变异丰富,但家系与地点效应独立。株高和穗位高的广义遗传力分别高达88.52%和93.78%,表明株高和穗位高遗传稳定性性强,受环境影响较小。

表2 郑单958 DH群体的株高和穗位高方差分析Table 2 Variance analysis of plant height and ear height for the DH population from Zhengdan 958
注:*,** 分别代表差异达0.05和0.01显著水平。
Note:*,** indicates significance at 0.05 and 0.01 level,respectively.
2.3郑单958DH群体株高和穗位高的QTL分析
2.3.1 郑单958 DH群体株高的QTL分析 从表3看出,长葛和淇县分别定位到2个和7个株高QTL,分别位于第1,2,3,5,7和8染色体上(图1),其中位于第1染色体上的qph-1-3和位于第2染色体上的qph-2-10在长葛和淇县均被检测到,2点共检测到7个株高QTL,这些QTL贡献率变幅为6.77%~21.31%,大于10%的有3个,其中qph-3-3的贡献率最高为21.31%。值得注意的是,由于qph-2-10是2个地点同时被检测到且贡献率均大于15%的QTL,据此推断该QTL可能是控制株高的主效QTL。在检测到的7个株高QTL中,除第3染色体上的qph-3-3以及第7染色体上的qph-7-10效应来自父本昌7-2之外,其他5个株高QTL效应均来自母本郑58。结合QTL的贡献率,不难看出,增加淇县点株高基因位点主要来自昌7-2的qph-3-3。
2.3.2 郑单958 DH群体穗位高的QTL分析 在长葛和淇县分别检测到5个和7个与穗位高QTL,分别位于第1,2,3,4,5和8染色体上,只有qeh-1-3是两地同时被检测到的唯一共同的QTL。尽管方差分析结果(表2)显示地点与家系之间互为不显著,但该QTL在两地的贡献率相差较大,分别为12.45%和7.54%,表明该QTL的表达受环境条件影响仍较大,存在明显的基因型与环境的互作效应;2个地点共检测到11个穗位高QTL,贡献率变幅为5.09%~17.12%,qeh-5-9位点的贡献率最高为17.12%。
2.3.3 郑单958 DH群体穗位系数的QTL分析 穗位系数是反映株型协调与否以及抗倒伏性的重要指标之一。本研究中长葛和淇县两点分别检测到3个和4个与穗位系数有关的QTL,分别位于第2,3和8染色体上, qeh/qph-3-1是两地同时被检测到的唯一共同的QTL,该QTL在两地的贡献率分别为14.67%和17.78%,是控制穗位系数的主效QTL。两地共检测到6个穗位系数QTL,贡献率变幅为6.89%~17.78%,这6个QTL均由来自母本郑58的基因起作用。值得注意的是,6个穗位系数QTL中的4个位于第2染色体上,其累加贡献率达38.25%,表明第2染色体上富集的相关QTL对决定株型具有重要作用。同时看出,位于第2染色体上与穗位系数有关qeh/qph-2-8,qeh/qph-2-15以及第3染色体上的qeh/qph-3-1分别与穗位高对应QTL相一致,而位于第2染色体上的qeh/qph-2-7和qeh/qph-2-14以及第8染色体上的qeh/qph-8-1又分别是长葛、淇县新检测到的特异表达的QTL。显然,控制穗位高和穗位系数的QTL不尽一致,其性状表现既有内在联系,又有个别性差异。

表3 郑单958 DH群体植株性状的QTL定位Table 3 Identification of QTLs for plant traits in DH population from Zhengdan 958
注:PVE/%为可解释的表型变异;a表示2个地点都被检测到的QTL。
Note: PVE/% means explainable phenotypic variation;a represents QTL detected in two locations.
3 结论与讨论
3.1有关玉米株高和穗位高QTL的一致性
目前,随着玉米机收面积的扩大和机收水平的提高,对玉米品种抗倒伏能力的要求越来越高, 大量研究表明,株高和穗位高与子粒产量呈正相关[14-16]。本研究的结果也显示,株高、穗位高表现出明显的正向杂种优势。然而,植株过高势必增加倒伏的风险,因此,深入分析株高、穗位高的遗传基础,协调好株高、穗位高与抗倒伏特性以及单株优势与群体产量之间的关系,是选育适应机收新品种中必须值得关注的重要课题。 国内外很多学者对株高和穗位高已有大量研究报道,并定位了许多相关的QTL,但不同学者因所用的群体和标记的密度不同,结果不尽一致。MILENA等[16]利用热带种质组配的F2∶3群体分别检测出6个株高和9个穗位高QTL;郑克志等[10]利用玉米自交系T 319与9406构建的RIL群体,检测到6个株高QTL,分布在第1,2,3,5,7,10染色体上,检测到2个穗位高的主效QTL,位于第1和第10染色体上。本研究利用郑单958诱导产生的DH群体,在第1,2,3,4,5,7,8染色体上共检测到7个株高和11个穗位高QTL,在株高QTL中贡献率最高的是位于bin 3.05-3.06的qph-3-3,贡献率高达21.31%,是淇县点特异表达的主效QTL,与MILENA等[16]、李清超等[9]、郑克志等[10]在bin 3.05-3.06发现的株高主效QTL一致或基本接近;位于bin 2.07的qph-2-10是长葛、淇县同时被检测到的QTL,贡献率分别达15.74%和17.55%,推测是一个控制株高的另一个主效QTL,与郑克志等[10]的结果基本一致。证明这些主效QTL是真实存在的。

图1 长葛和淇县检测到的玉米郑单958 DH群体QTL在染色体上的位置Fig.1 Position of QTLs for plant height and ear height detected in Qixian and Changge
本研究发现,控制玉米穗位高QTL的主效位点为bin 1.02处的qeh-1-3,bin 2.07的qeh-2-10和bin 4.07-4.08的qeh-4-8,与李清超等[9]研究发现的bin 1.01-1.02处富集穗位高QTL,在bin 1.02发现的穗位高QTL高度一致。同时,也检测到一些前人没有检测到位点,可作进一步发掘新的相关QTL的参考依据。
本研究检测到的株高和穗位高QTL中有4对即qph1-3与qeh1-3,qph2-10与qeh2-10,qph5-8与qeh5-8以及qph8-4与qeh8-4依次分别位于相同标记区间bnlg1429-bnlg1007,umc1637-umc1560,bnlg118-bnlg1118和umc1384-umc1069,尤其前两对QTL是两地同时被检测到的共同QTL,贡献率也比较高。如果这些成对被检测到的QTL是分别位于第1,2,5和第8染色体上的一因多效或连锁基因,则可一定程度上揭示株高和穗位高存在相关性的遗传原因,因而可进一步通过精细定位加以验证,为基因的克隆和聚合育种奠定基础。
3.2DH系群体的分离与表现
本研究中,利用的DH系是通过孤雌生殖诱导系诱导加倍后产生的,利用生物诱导孤雌生殖所产生的单倍体及其DH系的分离情况已有报道,如CHANG[17]和LASHEMES等[18]研究表明,由孤雌生殖诱导系途径产生的DH群体中,等位基因的分离均符合1∶1的遗传分离比例。文科等[19]利用孤雌生殖诱导系诱导高抗青枯病的1145和高感青枯病的Y 331的F1代而产生的单倍体进行分离的分析,通过卡方测验孤雌生殖单倍体的双亲的配子的比例数符合孟德尔1∶1的随机分离比例。对来源于杂交种1145×Y 331的92个DH和130个重组自交系群体(Recombination inbred lines, RIL)的株高、穗位等农艺性状比较分析表明,所有性状的平均值t测验结果表明二者差异不显著,2个群体主要农艺性状表现基本一致,DH群体表现出系内整齐性更高;可能的原因是由于该单倍体是利用玉米孤雌生殖诱导系诱导产生的,诱导系的作用并不影响F1代配子的重组交换及独立分离,只是促使单倍性胚的产生。此外,DH群体创建只需一年两代即可,大大缩减了构建稳定作图群体的时间,创建的DH群体可稳定繁殖,与早代的F2,F2:3等分离群体相比可消除单株间竞争的影响,减小环境的试验误差。
[1] FLINTGARCIA S A, JAMPATONG C, DARRAH L L, et al. Quantitative trait locus analysis of stalk strength in four maize populations[J]. Crop Science, 2003, 43:13-22.
[2] 李树岩, 王宇翔, 胡程达, 等. 抽雄期前后大风倒伏对夏玉米生长及产量的影响[J]. 应用生态学报,2015, 26(8):2405-2413.
[3] ALBRECHT K A, MARTIN M J , RUSSEL W A,et al. Chemical and in vitro digestible dry matter composition of maize stalks after selection for stalk strength and stalk-rot resistance[J]. Crop Science, 1986,26:1051- 1055.
[4] 王永学,张战辉,刘宗华. 玉米抗到伏性状的配合力效应及通径分析[J]. 河南农业大学学报,2011, 45(1):1-6.
[5] 付志远,邵可可,丁冬,等. 穗上节间数与玉米抗倒伏能力的相关性分析[J]. 河南农业大学学报,2011,45(2):149-154.
[6] 丰光,刘志芳,李妍妍,等. 玉米茎秆耐穿刺强度的倒伏遗传研究[J]. 作物学报,2009, 35(11): 2133-2138.
[7] 勾玲,黄建军,孙锐,等. 玉米不同耐密植品种茎秆穿刺强度的变化特征[J]. 农业工程学报,2010, 26(11):156-162.
[8] RADU A, PARASCHIVU U. Use of genetic estimates in breeding maize for resistance to stem breakage[J]. Lucrari Stiintifice(Romania),1944,7:70-80.
[9] 李清超,李永祥,杨钊钊,等. 基于多重相关RIL群体的玉米株高和穗位高QTL定位[J]. 作物学报,2013, 39(9):1521-1529.
[10] 郑克志,李元,瞿会,等. 玉米株高和穗位高的QTL 定位[J].江苏农业科学,2015,43(5):61-63.
[11] 郑德波,杨小红,李建生,等. 基于SNP标记的玉米株高及穗位高QTL定位[J]. 作物学报,2013, 39(3):549-556.
[12] KNAPP S J, STROUP W W, ROSS W M. Exact confidence intervals for heritability on a progeny mean basis[J].Crop Sci,1985, 25:1305-1320.
[13] 吴向远, 丁冬, 宋桂良,等. 玉米基因组DNA快速提取方法[J]. 河南农业大学学报,2012,46(1):7-10.
[14] MCCOUCH S R, CHO Y G, YANO M, et al. Report on QTL nomenclature[J]. Rice Genet Newslett, 1997, 14:11-13.
[15] 杨伟光,苏颖,张建华,等.玉米株高和穗位遗传模型测验[J].吉林农业大学学报,2000,22(4):28-13;44.
[16] MILENA L, SOUZA C, BENTO D, et al. Mapping QTL for grain yield and plant traits in a tropical maize population[J]. Mol Breed, 2006, 17:227-239.
[17] CHANG M. Stock 6 induce double haploidy is random[J]. Maize Genet, 1992, 67: 98-99.
[18] LASHERMES P, BECKERT M. Genetic control of maternal haploid in maize(ZeamaysL.)selection of haploid inducing lines[J]. Theor Appl Genet,1988, 76: 405-410.
[19] 文科,黎亮,刘玉强,等. 高效生物诱导玉米单倍体机器加倍方法研究初探[J]. 中国农业大学学报,2006,11(5):17-20.
(责任编辑:常思敏)
QTLanalysisofplantandearheightbyusingDHpopulationinmaize
LI Haochuan1, CHEN Qiong1, YANG Jiwei1, QU Yanzhi1, ZHANG Hui1, ZHANG Chaolin1, LI Yan1, JIA Xi2,LIU Zonghua1
(1.College of Agronomy, Henan Agricultural University, Zhengzhou 450002, China; 2.Agricultural Bureau of Lingbao City,Lingbao 472500, China)

corn; doubled haploid; plant height; ear height; quantitative trait locus
S 513
:A
2015-12-15
国家自然科学基金项目(U1204316);国家“863”计划项目(2012AA100300)
李浩川(1978-),男,河南舞钢人,副教授,博士,主要从事玉米遗传育种研究。
刘宗华(1960-), 男,河南三门峡人,教授,博士生导师。
1000-2340(2016)02-0161-06
猜你喜欢
江苏科技报·E教中国(2024年11期)2024-12-31 00:00:00
河南农业科学(2023年9期)2023-10-18 01:19:32
中国种业(2021年7期)2021-08-02 01:40:42
山西农业科学(2020年8期)2020-08-13 07:44:06
北方音乐(2020年11期)2020-07-09 03:16:08
河南水利年鉴(2020年0期)2020-06-09 05:44:24
河南农业科学(2018年5期)2018-01-19 08:25:55
河南水利年鉴(2017年0期)2017-05-19 02:30:57
中国学术期刊文摘(2015年8期)2015-10-29 09:51:18
西北农林科技大学学报(自然科学版)(2015年5期)2015-02-21 16:17:12
