
长链非编码RNA的研究进展
2016-09-22 08:57:14吴王亲张婷田男
世界复合医学 2016年1期
吴王亲,张婷,田男
浙江中医药大学生命科学学院,浙江杭州 310053
长链非编码RNA的研究进展
吴王亲,张婷,田男
浙江中医药大学生命科学学院,浙江杭州 310053
长链非编码RNA(long non-coding RNA,lncRNA)是位于真核细胞内,长度超过200nt的不具阅读框的非编码RNA。在哺乳动物基因组中被普遍转录,其表达具有时空特异性。lncRNA参与X染色体沉默、基因组印记、染色质修饰、转录激活、转录干扰、核内运输等多种重要的调控过程,并与疾病的发生、发展、诊断和治疗有密切的关系。lncRNA覆盖面之广、潜力之大,使得其成为当今生物学中最热门最前沿的研究领域之一。本文将从lncRNA的研究历程、来源与分类、作用机制和功能研究、在疾病中的作用这几个方面进行总结和论述。
长链非编码RNA;疾病;作用机制;诊断;治疗
1 lncRNA的研究历程
长链非编码RNA(Long non-coding RNA,lncRNA)是一类位于细胞核或细胞质内的功能性RNA分子,由RNA聚合酶 II(RNA Polymerase II)或 RNA聚合酶 III(RNA Polymerase III)转录而成,其重复序列较少,半衰期较短,结合位点单一,能在多种层面调控基因表达水平。
1965 年Holley等[1]首次在面包酵母中发现一种非编码RNA(non-coding RNA,ncRNA)。2002年,Okazaki等[2]在小鼠全长 cDNA(complementary deoxyribonucleic acid)文库大规模测序中,鉴定了大量较长的非编码RNA转录本,因而提出lncRNA的概念,但因lncRNA不具蛋白编码潜能,故被认为是基因组在进化过程中累积的不具任何生物学功能的“转录噪音”,未能引起关注。而后Ponjavic等[3]对哺乳动物基因组功能注释计划(Functional Annotation of the Mammalian Genome,FANTOM)转录组数据进行分析后发现lncRNA初级序列、启动子序列和剪接方式表现为负选择,具有不为人知的功能。直到2007年,Rinn等[4]报道了一条2.2 kb长的lncRNA HOTAIR(HOX transcript antisense RNA),可与蛋白复合体polycomb相互作用,抑制HOX(homeotic genes)基因的转录并修饰染色质,进而起到调节生物体生长发育的作用。lncRNA作为体内“暗物质”无所不在的一面被揭开一角,由此lncRNA被越来越多的关注,大量 lncRNA得到鉴定(如H19,MALAT-1,HOTAIR等),并随着人们对lncRNA的认识日益深入,研究热潮日益高涨。
2 lncRNA的来源和分类
2.1 lncRNA的来源
lncRNA的来源大致有以下几种:①蛋白编码基因结构中断,发生框架破坏,与前体编码序列一同转变为有功能的非编码lncRNA;②两个不转录且先后分离的序列通过染色质重组合并形成一个多外显子lncRNA;③非编码基因在复制过程中反移位产生lncRNA;④局部复制子串联产生相邻重复序列产生lncRNA;⑤基因中插入转座成分而产生有功能的lncRNA。
2.2 lncRNA的分类
lncRNA的命名及分类尚无统一规定,目前被大部分学者所认可的分类有以下几种:①依其相对于蛋白编码的基因位置分为:正义型(与同链蛋白编码基因转录方向相同)、反义型(与同链蛋白编码基因转录方向相反)、双向型 (可同时向同链蛋白编码基因转录方向相同或相反的方向转录)、基因内型(从基因的内含子区转录得到)、基因间型(从两基因间区域转录得到);②依其分子效应机制分为:信号分子(调控其他基因表达)、诱饵分子(间接调控目标基因转录)、引导分子(指导蛋白复合体定位至调控点)、支架分子(为多个相关分子提供复合体装配中心平台);③依其在DNA序列中所起的作用分为:顺式lncRNA(调节临近基因表达)、反式lncRNA(调节远处基因或其他基因表达);④另外 Kung等[5]将其分为独立lncRNA(stand-alone lncRNA),自然反义转录体(natural antisense transcripts,NAT)、假基因(pseudogenes)、长链固有ncRNA(long intronic,ncRNA)其他RNA(增强子RNA等)五种。
3 lncRNA的作用机制和功能研究
lncRNA不具有多聚腺苷酸尾,其保守性亦较差,但相对于其他RNA,lncRNA的序列更长,空间结构更复杂,所含信息更丰富,参与表达的分子机制更加多样,其表达具有较高的组织性和细胞特异性。它主要从表观遗传、转录水平和转录后水平这三个层面调控基因表达,根据其发挥作用的主要生物学机制又可再细分为:DNA甲基化与去甲基化、组蛋白修饰、染色质重塑、细胞周期调控、剪接调控、mRNA降解和翻译调控等[6]。
3.1 表观遗传调控
表观遗传主要指遗传表型和遗传表达发生了可遗传的改变,但不涉及DNA序列的变化,主要包括DNA甲基化与去甲基化、组蛋白修饰、染色体重塑等形式。
3.1.1 DNA甲基化与去甲基化
DNA甲基化是由DNA甲基化转移酶(DNA methyltransferase,DNMT)介导催化和维持的一种表观遗传修饰形式,发生在细胞的印记控制区。lncRNA在由DNA甲基化介导的基因转录失活过程和DNA去甲基化介导的基因活化过程中都起着重要的作用。在通过对小鼠7号染色体远端Kcnql区和17号染色体近端Igf2受体区的研究发现,lncRNA能够通过诸如顺式作用介导印迹中断的父源性基因甲基化[7-8]。
3.1.2 组蛋白修饰
组蛋白修饰是组蛋白经某些修饰酶的作用而发生去乙酰化、二甲基化、三甲基化等共价修饰。lncRNA通过参与组蛋白修饰,抑制或激活基因的表达。如Heo JB等[9]在对拟南芥的研究中发现,春化作用诱导产生的 lncRNA COLDAIR与起始复合体 (Polycomb repressive complex 2, PRC2)结合能引起H3K27三甲基化,从而抑制拟南芥开花相关的基因。
3.1.3 染色质重塑
染色质重塑包括核小体转位、重组及稳定性降低等改变。通过改变DNA调控序列与转录因子的亲和性,从而影响相关基因的表达。lncRNA于此亦有重要作用,其通过招募抑制性复合物(如Repa protein招募PRC2)在染色体外形成外包裹、甲基化组蛋白等方式,使染色体上某一片段失活(如X染色体中Xist顺式作用介导X染色体失活,HOTAIR诱导HOX D基因座上转录文本发生沉默等),但其活性可被相应的反转录文本所限制(如Tist限制Xist对于X染色体的失活)。
3.2 转录调控与转录后调控
lncRNA可通过干扰临近基因的表达引发染色体重塑调控基因表达,也可结合基础转录因子(Basal transcription factor)灭活启动子,激活辅助蛋白灭活靶基因,如二氢叶酸还原酶(Dihydrofolatereductase,DNFR)次级基因启动能产生与一般转录因子IIB和二氢叶酸还原酶主要启动子DNA结合的lncRNA,三者形成特殊稳定三重复合体,导致起始前复合物的分解和转录抑制,从而达到调节代谢性基因编码二氢叶酸还原酶的目的。另外,lncRNA可作为共因子调节转录因子的活性等机制参与转录调控。
lncRNA还能够作为小RNA前体,靶向的与mRNA形成双链RNA复合物从而掩盖mRNA主要顺式作用元件等方式,影响mRNA的剪接、转运和降解等过程,调控基因表达,达到转录后调控的目的。如人肺腺癌转移相关转录本 1(Metastasis associated lung adenocarcinoma transcript 1,MALAT1)通过交接丝氨酸/精氨酸剪切因子的磷酸化,达到使pre-mRNA发生选择性剪接的目的[10]。
3.3 其他功能
受核心转录因子调控的lncRNA也可调控多能性干细胞的发展过程,据报道有26种lncRNA是维持细胞全能性所必须,并且lncRNA在促进细胞分化上起着非常重要的作用,近年来对拟南芥的研究和对“海绵作用”的研究表明lncRNA能通过协同作用调控基因表达。lncRNA同样影响许多涉及突触发生和组织重排的形态发生过程,并且作为假基因成员影响同源基因表达。可以说lncRNA在生物学功能上的作用是无处不在的。
4 lncRNA与肿瘤
lncRNA作为一种重要的基因表达调控元件,其与疾病的发生、发展、诊断和治疗有着密切的关系。目前为止已发现多种与肿瘤密切相关的lncRNA,根据其在参与肿瘤发生发展、凋亡调控、浸润转移中的作用,大致可分为促肿瘤作用lncRNA和抑肿瘤作用lncRNA。
4.1 促肿瘤作用的lncRNA
促肿瘤作用的lncRNA往往在肿瘤特别是转移性肿瘤中高表达,如HOTAIR,自HOX基因转录出后,既可招募PRC2至HOXD基因座,触发HOXD的沉默,又可直接招募PRC2至细胞并使其失活,影响HOXD的甲基化,从而增强PRC2依赖的肿瘤细胞的入侵和代谢途径。人肺腺癌转移相关转录本MALAT 1在细胞中多存在于核小斑结构(储存、加工MRNA前体剪切子的重要场所),其过度表达可下调肿瘤抑制因子PSF,参与肿瘤发生的共同环节。此外,在对结肠癌细胞的研究中发现,MALAT 1 3’端存在一段特殊序列核苷酸(6918-8441 nt),与结直肠癌SW620、SW480细胞的转移行为直接相关,且下调MALAT 1能有效抑制肿瘤细胞转移[11]。
4.2 抑肿瘤作用的lncRNA
p53是一种重要的肿瘤抑制基因,能通过肿瘤抑制途径诱发多个lncRNAs的表达,其中p53能直接诱导lncRNA-p21,lncRNA-p21可通过与 hnRNP-K(heterogeneous nuclear ribonucleoprotein K)的相互作用而控制其下游基因的表达,从而抑制肿瘤细胞的增殖或促进异常细胞的凋亡[12]。同时抑肿瘤作用的lncRNA的缺失或因序列突变导致lncRNA失活往往导致肿瘤的发生,如现为人熟知的母系表达基因3(Maternally expressed gene 3,MEG3),其缺失能引起肿瘤的发生,而其正常表达则能促进细胞凋亡,抑制肿瘤细胞增殖。
4.3 有些基因则具促癌和抑癌的双重功能
Shi Y等[13]发现H19的低表达能抑制miR-675的表达,而miR-675能直接靶向结合钙粘蛋白(Cadherin,Cad)3’非编码区,从而达到调节钙黏蛋白的表达并促使细胞侵袭的目的。但另一方面,Matouk等[14]又发现H19在肺癌及前列腺癌中低表达,尤其在前列腺癌中,H19和胰岛素样生长因子2(insulin-like growth factor 2,IGF2)的表达受甲基化分化区域(differentially methylated region,DMR)调控(DMR基因位于H19上游2kb处,富含CpG,,而CpG的差异甲基化调控H19/IGF2的表达),有研究发现,前列腺增生中DMR基因的CpG甲基化与前列腺癌相比,是后者的2倍,这证明了H19在前列腺癌中的抑癌作用。以上结果均表明H19具有促癌和抑癌的双重作用。
我们查阅了大量了资料,总结出lncRNA与癌症相关性图表如下[15-25]。
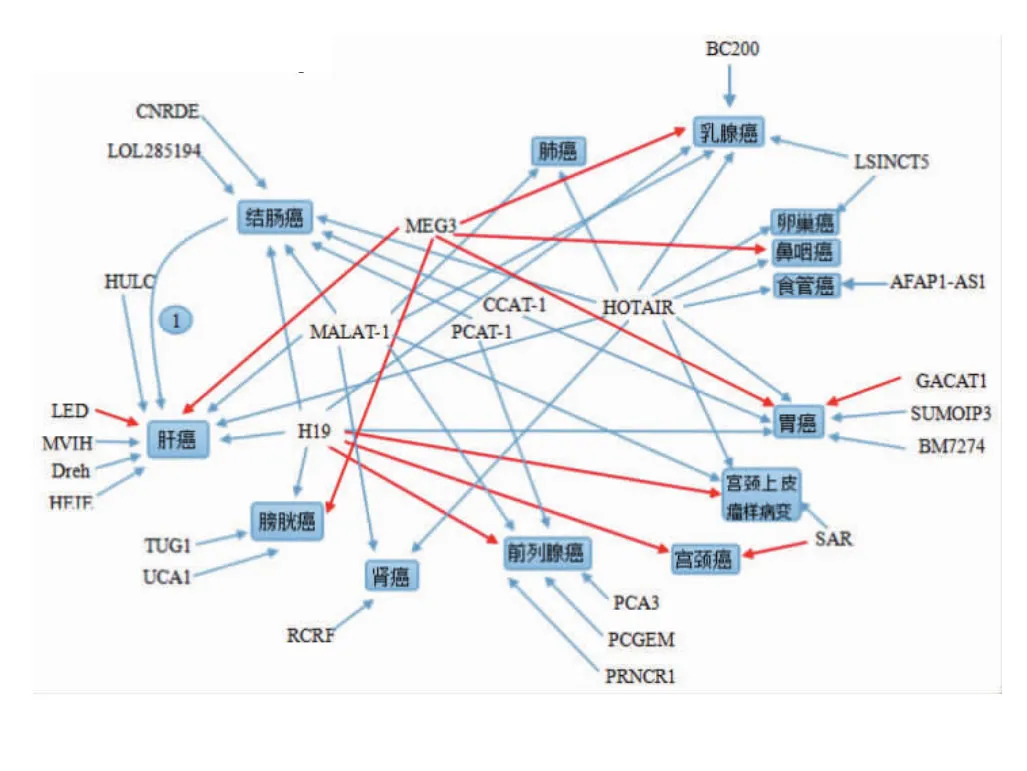
图1 lncRNA与的肿瘤相关性
5 lncRNA与其他疾病
人们在对lncRNA的研究中,除发现与肿瘤相关的的一些lncRNA以外,还发现了许多与其他疾病有密切关系的lncRNA。这些lncRNA在这些疾病的发生发展过程中也起着极为重要的作用。
5.1 lncRNA在心血管疾病中的作用
随着对全基因组的关联研究,人们发现人染色体9p21区域是一重要的冠心病易感区域,其上有一位于INK4基因座的lncRNA ANRIL(Antisense noncoding RNA in the INK4 locus),ANRIL可以连接和起始复合体1(Polycomb repressive complex 1,PRC1)和起始复合体2(Polycomb repressive complex 2,PRC2),沉默ANRIL/PRC介导的INK4位点的基因,使p14、p15、p16的表达下调,促进血管平滑肌细胞增殖和动脉粥样板块的形成[26]。这一位点还与糖尿病、颅内动脉瘤、牙周炎、子宫内膜异位、青光眼等疾病相关[27]。而对平滑肌的研究发现与血管紧张素 II相关的lncRNA可调节平滑肌细胞对血管紧张素II的敏感性,并可降解产生两种与平滑肌细胞增殖密切相关的miRNA[28]。另一个lncRNA,NPPA-antisense可通过调控钠尿肽前体蛋白A(natriuretic peptide precursor A,NPPA)基因的选择性剪切,影响心房钠尿肽的浓度,进而参与血压调节,虽然NPPA只在婴儿期表达,但Horsthuis等[29]研究发现,其在心肌肥厚和心衰病人中可被重新激活。罗雨虹[30]对比在心梗后左心房重塑和心梗后不发生左心房重塑的病患的lncRNA后,发现七个变化显著且较稳定的lncRNA,且这些lncRNA都是来源于线粒体DNA。Mahadevan等[31]还发现了一种能导致强直性肌营养不良的毒性lncRNA DMPK 3'UTR,该疾病中,CTG三连密码子在萎缩性及强直蛋白激酶(Human myotonin-protein kinase,DMPK)基因的3’UIR重复数可达2000多次,并且Yadava等[32]还通过在小鼠身上复制该疾病成功证明了lncRNA在心脏功能障碍中的独立功能。
5.2 lncRNA在糖尿病中的作用
目前,相关研究发现与糖尿病密切相关的lncRNA有以下几种:lncRNA细胞周期激酶抑制因子4(INK4)位点的反义编码 RNA(antisense non-coding RNA in the INK4 lo-cus,ANRIL)、葡萄胎相关印迹基因(hydatidiformmole associated and imprinted,HYMAI)、胰岛素样生长因子2反义RNA(insulin-like growth factor 2 anti-sense RNA,IGF-2AS)、母系表达基因 3(maternallyexpressed gene,MEG3)、浆细胞瘤变体易位基因1(plasmacytoma variant translocation 1 gene,PVT1)和HI-LNC25,这些 lncRNAs通过表达的异常、甲基化和基因多态性等多种途径广泛参与糖尿病的发生与发展[33]。
5.3 lncRNA在神经系统疾病中的作用
lncRNA通过与其他表观遗传调控形式的相互作用,参与调控和维持神经系统功能的稳态,其在神经系统疾病中起着极为重要的作用。
Faghihi等[34]发现淀粉蛋白前体β位分解酶1(BACE1)的一种自然反义lncRNA BACE1-AS,能在外界条件刺激下与BACE1 mRNA形成复合物,增加其稳定性从而防止被核酸酶降解。这一过程会导致更多β-淀粉样蛋白(Aβ)的积累,从而再次促进BACE1-AS的表达并形成正反馈循环,加速阿尔茨海默症(AD)的发生。此外Mus等[35]发现额叶皮质中lncRNA BC200的变异表达和非正常定位也会影响AD的发生和推动病程进展。
再如与脆弱X综合症(Fragile X Syndrome,FXS)密切相关的lncRNA FMR1和ASFMR1,在其患者中呈现表达增加[36]。NaPINK1通过稳定期靶基因的正义转录产物svPINK1,干扰线粒体呼吸链功能,提高细胞对凋亡信号的敏感性,从而参与帕金森病的发病进程[37]。除此之外,lncRNA还与神经分裂症、唐氏综合症、多发性硬化等多种神经系统疾病有关。
5.4 lncRNA在感染性疾病中的作用
Peng等[38]用SARS冠状病毒(SARS-CoA)感染小鼠,以不完全随机6寡核苷酸方法[39]反转录rRNA以外其他RNA,并测定了细胞lncRNA在感染前后的变化,发现509个已经注释的lncRNA和1046个未注释的lncRNA在感染前后发生了显著的变化,对其中的37个lncRNA进一步研究发现感染流感病毒后大多数lncRNA发生了与SARS-CoA感染相似的变化。不过对于lncRNA在病毒感染中的具体作用,到目前为止只有一些关于其可能会影响蛋白质编码基因的表达的研究。
6 展望
lncRNA通过DNA甲基化与去甲基化、组蛋白修饰、染色质重塑、细胞周期调控、剪接调控、mRNA降解和翻译调控等多个途径调控生物生长发育及生理病理过程,与各类癌症、心血管疾病、神经系统疾病、感染、糖尿病等息息相关。可以说,lncRNA在生物体整个生命阶段都是无处不在的。
迄今所鉴定的功能性lncRNA与根据生物信息学所推测的数目相比仅是冰山一角。但lncRNA在疾病的分级检测、分子分型、疗效评估、芯片应用等方面均已展现出了巨大的应用前景。随着技术的更新和数据库的建立完毕,对lncRNA研究的限制将越来越少,lncRNA的开发将进入一个新时期,包括lncRNA在内的极其复杂的生物体内的动态信息交换网络将会一步步被揭开,这些会带给我们关于生老病死等问题新的答案[40]。
(
)
[1]Holley RW,Apgar J,Everett GA,et al.Structure of aribonucleic acid[J].Science,1965,147(3664):1462-1465.
[2] Okazaki Y,Furuno M,Kasukawa T,et al.Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs[J].Nature,2002,420(6915):563-573.
[3]Ponjavicn J,Ponting CP,Lunter G..Functionality or transcriptional noise Evidence for selection within long noncoding RNAs[J].Genome Research,2007,17(5):556-565.
[4]Rinn JL,Kertesz M,Wang JK et al.Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs[J].Cell,2007,129(7):1311-1323.
[5]Kung JT,Colognori D,Lee JT.Long noncoding RNAs:past, present,and future[J].Genetics,2013,193(3):651-669.
[6]Zhu S,Zhang XO,Yang L.Panning for long noncoding RNAs[J].Biomolecules,2013,3(1):226-241.
[7]Pauler FM,Koerner MV.Barlow DP.Silencing by imprinted noncoding RNAs:is transcription the answer [J].Trends Genet,2007,23:284-292.
[8]Nagano T,Mitchell JA,Sanz LA,et al.The Air noncoding RNA epigenetically silences transcription by targeting G9a to chromatin[J].Science,2008,322(5908):1717-1720.
[9]Heo JB,Sung S.Vernalization-mediated epigenetic silencing by along intronic noncoding RNA [J].Science,2011, 331(6013):76-79.
[10]Tripathi V,Ellis JD,Shen Z,et al.The enuclear-retained noncodingRNA MALA T1 regulates alternative splicing by modulating SR splicing factor phosphorylation [J].Mol Cell,2010,39(6):925-938.
[11]Xu C,Yang M,Tian J,et al.MALAT-1:a long non-coding RNA and its important 3’end functional motif in colorectal cancer metastasis[J].Int J Oncol,2011,39(1):169-175.
[12]Huarte M,Guttman M,Feldser D,et al.A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response[J].Cell,2010,142(3):409-419.
[13]Shi Y,Wang Y,Luan W,et al.Long Non-Coding RNA H19PromotesGliomaCellInvasionbyDerivingmiR-675[J]. PLoS ONE 2014,9(1):e86295.
[14]Matouk IJ,Mezan S,Mizrahi A,et al.The oncofetal H19 RNA connection:hypoxia,p53 and cancer [J].Biochim Biophys Acta,2010,1803(4):443-451.
[15]崔玲,谢晓砚,王和,等.长链非编码RNA HOTAIR mRNA在卵巢癌中的表达[J].四川大学学报,2013,44(1):57-59.
[16]杨韬,李俊堂,王丽娟,等.干涉lncRNAs HOTAIR对人乳腺癌细胞MDA-MB-231的影响[J].细胞与分子免疫学杂志,2012,28(1):97-98.
[17]裴昌松,范方田,武艺,等.肾癌细胞系中HOTAIR表达的测定及稳定高表达HOTAIR肾癌细胞株的建立 [J].南京医科大学学报,2014,34(5):548-552.
[18]宋尚卿,吴震杰,王承,等.RCRF在高级别肾癌组织中表达上调并影响肾癌患者预后 [J].第二军医大学学报, 2014,35(5):566-568.
[19]宫再兴,王共先,刘伟鹏,等.泌尿系恶性肿瘤中长链非编码RNA表达变化的研究进展 [J].山东医药,2013,53 (38):96-99.
[20]林梦洁,陈志强,尹凌帝,等.长链非编码RNA HOTAIR对非小细胞肺癌迁移和侵袭能力的影响 [J].临床肿瘤学杂志,2014,19(8):684-689.
[21]王剑,崔明.长链非编码RNA与乳腺癌相关性的研究进展 [J].西南国防医药,2013,23(3):342-344.
[22]陈标,朱萱.长链非编码RNA在肝癌中作用的研究进展[J].世界华人消化杂志,2014,22(31):4762-4768.
[23]杨慧,余敏敏,陆晓媛.长链非编码RNA在宫颈癌及宫颈上皮内瘤样病变中的表达 [J].江苏医药,2014,40(22): 2685-2688.
[24]金桂花,王丽娟,朱青,等.长链非编码RNA在膀胱癌中的研究进展[J].西部医学,2013,9(25):1418-1421.
[25]胡谦,胡小云.长链非编码RNA在胃癌中的研究进展 [J].世界华人消化杂志,2014,22(4):501-507.
[26]成捷,刘新光,熊兴东,等.miRNA,lncRNA与心血管疾病 [J].中国生物化学与分子生物学报,2014,30(4): 328-335.
[27]Congrains A,Kamide K,Katsuya T,et al.CVD-associated non-coding RNA,ANRIL,modulatesexpression of atherogenic pathways in VSMC [J].Biochem Biophys Res Commun,2012,419:612-616.
[28]Gizard F,Heywood EB,Findeisen HM,et al.Telomerase activation in Atherpsclerosis and induction of telomerase reverse transcriptase expression by inflammatory stimuli in macrophages[J].Arterioscler Thromb Vasc Biol,2011,31 (2):245-252.
[29]Yin Z,Guan D,Fan Q,et al.LncRNA expression signatures in response to enterovirus 71 infection [J].Biochem Biophys Res Commun,2013,430(2):629-633.
[30]罗雨虹.循环中的长非编码RNA-LIPCAR预示心衰患者的生存率 [J].生理科学进展,2014,114(4):1569-1575.
[31]Horsthuis T,Houweling AC,Habets PE,et al.Distinct regulation of developmental and heart disease-induced atrialnatriuretic factor expression by two separate distal sequences[J].Circ Res,2008,102(7):49-859.
[32]Mahadevan M,Tsilfidis C,Sabourin L,et al.Myotonic dystrophymutation:an unstable CTG repeat in the 3’untranslated region of the gene [J].Science,1992,255 (5049):1253-1255.
[33]韩泽平,何金花,黎毓光,等.长链非编码RNA与糖尿病关系的研究进展[J].中国病理生理杂志,2014,30(4): 751-756.
[34]Faghihi MA,Modarresi F,Khalil AM,et al.Expression of a noncoding RNA is elevated in Alzheimer’s disease and drives rapid feed-forward regulation of beta-secretase[J]. Nat Med,2008,14(7):723-730.
[35]Mus EI,Hof PR,Tiedge H.Dendritic BC200 RNA in aging and in Alzheimer’s disease[J].Proc Natl Acad Sci USA,2007,104(25):10679-10684.
[36]Qureshi IA,Mattick JS,Mehler MF.Long non-coding RNAs in nervous system function and disease[J].Brain Res,2010,1338:20-35.
[37]Sai Y,Zou Z,Peng K,et al.The Parkinson’s disease-related genes act in mitochondrial homeostasis[J].Neurosci Biobehav Rev,2012,36(9):2034-2043.
[38]Peng X,Gralinski L,Armour CD,et al.Unique signatures of long noncoding RNA expression in response to virus infection and altered innate immune signaling [J].MBio, 2010,1(5):e00206-10-e00206-18.
[39]Armour CD,Castle JC,Chen R,et al.Digital transcriptome profiling using selective hexamer priming for cDNA synthesis[J].Nat Methods,2009,6(9):647-649.
[40]徐鹏辉.美国启动精准医疗计划[J].世界复合医学,2015, 1(1):44-46.
The Research Progress of Long non-coding RNA
WU Wang-qin,ZHANG Ting,TIAN Nan
College of Life Science,Zhejiang Chinese Medical University,Hangzhou,Zhejiang Province 310053 China
Long non-coding RNA;Disease;Mechanism of action;Diagnosis;Treatment
R394.3
A doi 10.11966/j.issn.2095-994X.2016.02.01.18
2015-12-18;
2016-02-04
浙江中医药大学校级基金重点项目(2014ZZ05);浙江省自然科学基金青年基金项目(no.LQ12C07001);浙江省教育厅一般科研项目(Y201121464);国家级大学生创新创业训练计划项目(201410344004)。
田男(通讯作者),博士,讲师,研究方向:RNA,E-mail:tiannanlux@126.com。
吴王亲,张婷,田男.长链非编码RNA的研究进展[J].世界复合医学,2016,2(1):63-68.
Abstrat Located in eukaryotic cells,long non-coding RNA(lncRNA)does not have the reading frame,with a length of more than 200nt.It is widely transcribed in mammalian genome and its expression spatiotemporal specificity.lncRNA takes part in many important regulational processes,such as X chromosome silence,genomic imprinting,chromatin modification,transcription activation,transcription interference,nuclear transport etc.And it's closely related with occurrence,development,diagnosis and treatment of the disease.It has such wide coverage and potential,which makes the study of lncRNA become one of today's most cutting-edge biology of the most popular areas.This article will summarize the history of study,origin and classification,mechanism action and function of lncRNA,and the role of lncRNA in the disease.
猜你喜欢
昆明医科大学学报(2021年4期)2021-07-23 01:21:44
昆明医科大学学报(2021年3期)2021-07-22 07:40:08
制造技术与机床(2019年10期)2019-10-26 02:47:12
中成药(2017年9期)2017-12-19 13:34:44
中国塑料(2015年10期)2015-10-14 01:13:16
医学研究杂志(2015年4期)2015-06-10 06:42:43
现代检验医学杂志(2015年2期)2015-02-06 02:00:48
沈阳医学院学报(2014年4期)2014-12-27 13:44:30
肿瘤预防与治疗(2014年5期)2014-10-18 01:05:23
遗传(2014年3期)2014-02-28 20:59:04
