
大气CO2浓度和温度互作对水稻生长发育的影响
2016-09-21 02:33景立权赖上坤王云霞杨连新王余龙
生态学报 2016年14期
景立权,赖上坤,王云霞,杨连新,*,王余龙
1 扬州大学, 江苏省作物遗传生理国家重点实验室培育点, 粮食作物现代产业技术协同创新中心,扬州 225009 2 扬州大学, 环境科学与工程学院,扬州 225009
大气CO2浓度和温度互作对水稻生长发育的影响
景立权1,赖上坤1,王云霞2,杨连新1,*,王余龙1
1 扬州大学, 江苏省作物遗传生理国家重点实验室培育点, 粮食作物现代产业技术协同创新中心,扬州2250092 扬州大学, 环境科学与工程学院,扬州225009
大气二氧化碳(CO2)浓度和温度的增加是全球气候变化的两个最主要特征。目前空气中的CO2浓度已从1800年的不到280 μmol/mol上升到391 μmol/mol,预测本世纪末最高将增至936 μmol/mol。伴随CO2及其它温室气体增强的温室效应,相比1980—1999年,2100年之前全球地表平均气温将增高1.5—4.0℃。水稻是人类最重要的食物来源,为全球半数以上人口提供营养。在介绍CO2浓度和温度增高试验平台的基础上,系统总结了CO2浓度和气温这两个重要的环境因子特别是两者的交互互作对水稻影响的实验进展,内容包括光合作用、生育进程、分蘖发生、物质生产、籽粒产量、受精过程、碳氮代谢、稻米品质以及水稻/杂草竞争等方面。结果表明,作为光合作用的底物,大气CO2浓度增高对水稻生产力的直接影响通常是有益的;相反,气温升高及其与CO2的互作对水稻各生长过程的影响变异很大(从负到正),反映了处理因子(包括CO2-温度处理水平和时间)、供试品种及其生长条件之间复杂的交互作用。目前这一方向有限的认识多来自于封闭或半封闭气室的研究,未来研究的重点是利用稻田T-FACE(Temperature-Free Air CO2Enrichment)技术结合气室试验展开更多更深入的学科交叉研究,研明CO2浓度与温度的交互作用对水稻关键生长过程的影响,并找出这些互作效应的生物学机制,增强人们对气候假定情景下水稻响应的预测能力,进而更加有效地制订出应对气候变化的适应策略。
水稻;气候变化;二氧化碳;温度;生长发育
大气二氧化碳(CO2)浓度持续增高是全球气候变化最为突出和确定的现象之一。截止2011 年,大气中CO2浓度已经达到391 μmol/mol,比工业革命前的1750 年升高了40%,达到80万年以来的最高值,预计到2100年最高可达936 μmol/mol[1]。空气中的CO2是植物光合作用碳固定的主要来源。大多数植物利用C3或C4光合途径固定空气中的碳。C3植物(如水稻)约占全球所有植物的95%[2],其CO2饱和点高于当前大气CO2水平,因此空气中CO2浓度增高能直接增强C3植物光合能力,进而会促进绝大多数栽培和野生物种的生长[3]。大气CO2和其它温室气体浓度增高对环境的一个主要影响是全球暖化潜势:通过吸收空气中的太阳能量,进而使地表气温升高[4]。过去3个10年的地表已连续偏暖于1850年以来的任何一个10年,在北半球,1983—2012年可能是过去1400年中最暖的30年。对于未来趋势,IPCC预测[5],相比1980—1999年,本世纪末地球表层气温将平均增加1.8—4.0℃,可能的变幅为1.1—6.4℃。气温升高将会直接或间接地影响植物的一系列代谢过程,进而改变植物的最终生产力[6-7]。
在全球气候变化以及资源竞争不断加剧的背景下,2050年全球粮食需求将会加倍[8]。水稻是人类最重要的食物来源,为全球半数以上人口提供营养。大气CO2浓度[9-10]和温度[11]都是影响水稻生长发育的关键因子,两者同时升高将对稻作生产产生重大影响。即使不考虑全球变暖,由于水稻生长在非常广阔的气候带,这些气候带的气温往往存在很大差异。因此,无论对世界粮食安全还是地区可持续的稻作生产,研明CO2浓度、温度特别是两者互作对水稻的影响在现在和未来均有重要意义。由于温室气体导致的全球变暖受到普遍关注,目前国际学术界越来越重视CO2,温度互作的实验研究,但同单因子操作试验[9-11]相比,研究的广度和深度均显不足。
本文在简要回顾水稻作物CO2浓度与温度互作研究中主要实验平台发展历史的基础上,系统总结了大气 CO2浓度和气温升高对水稻生长发育影响的主要研究进展,重点评估了两因子间可能存在的互作效应,并对该领域未来研究提出建议。尽管也有不少CO2,温度互作对水稻影响的模型研究,但不属本篇综述的范围。
1 实验平台的发展历史
尽管科学家早在50年前就利用熏气法试验系统开展CO2对水稻影响的研究[9],但直到20世纪90年代这些系统经过改进才被陆续用于CO2与温度的互作研究,这些改进的试验系统主要包括全封闭的环境模拟室、CO2-温度梯度气室(CTGC)、开顶式气室(OTC)、开放式空气中CO2浓度和气温增高(T-FACE)系统(表1)。前两种方式是在1个封闭的腔体中采用人工控制水稻生长所需的光、温、水、肥、气等条件,最突出的优点是试验成本较低而处理控制精度高,最主要的缺点是封闭导致的壁箱效应,对作物生长环境改变很大。由于气室广泛采用盆钵培育作物,其根系生长受限加上边际效应的存在,与大田群体条件下水稻对CO2的响应存在较大差异,一般认为气室数据采信度相对较低[12-13]。

表1 水稻CO2-温度互作研究主要试验系统的比较
为了能在自然条件下开展气候变化因子对水稻影响的研究,能观察大田水稻响应的设备应运而生,首先出现的是半封闭式的顶部开口的室外气候模拟室技术,即开顶式气室(OTC),这种多为室外搭建的自然采光系统,可在近似大田条件下设置不同CO2浓度和温度水平(表1)。但总体上这种系统与纯自然条件还是存在差异。20 世纪80 年代末,美国科学家Hendrey及其同事率先发展了自由空气中CO2浓度增高的控制技术,即FACE(Free-Air CO2Enrichment)系统,为解决这一问题提供了可能[45]。与气室系统相比,FACE研究采用试验地标准的作物管理技术,在空气完全自由流动的农田环境下运行,是目前最接近于自然生态环境的模拟系统[13]。FACE技术首先被用于棉花的研究[45],1998年起先后被日本[46]和中国科学家[47]用于水稻研究。2013年,中国科学家通过技术攻关,将原有的稻田CO2-FACE系统(已连续运行12年)升级改建为T-FACE试验平台(图1),在空间尺度较大的开放空气中模拟CO2浓度和温度伴随升高的未来情景,研究两者交互作用下水稻主要生长过程的响应和适应[44]。

图1 位于中国扬州(119°42′E,32°35′N)的稻田T-FACE平台(2014年8月拍摄)Fig.1 T-FACE platform located in paddy field at Yangzhou, China (119°42′E,32°35′N)(Photograph in August 2014)(a) CO2供气装置, CO2 supply apparatus; (b) Ambient圈全景, The full view of Amibient ring; (c) FACE圈全景, The full view of FACE ring; (d) FACE圈增温区, temperature increment plot in the FACE ring
2 光合作用和呼吸作用
全球气候变化生物学的一个重要课题就是确定CO2浓度和温度对作物光合作用的互作效应。两者对水稻光合作用是否有互作效应存在不同观点。高温使作物光呼吸及随后的碳损失增加,而CO2浓度升高通过减少光呼吸使碳损失总量减少,因此理论上单叶光合作用的最适温度随CO2浓度升高而增加[48]。分析不同CO2浓度下短期熏蒸试验数据发现,在15—35℃范围内,对水稻单叶光合速率而言,温度对CO2响应的调节效应为4%—5%/℃[14,16](即温度每增加1℃,CO2响应值增加4%—5%),这一结果与生化光合模型预测的结果基本一致[48]。但在最适温度以下,长期CO2熏蒸试验[38]获得的光合响应数据(2%—3%/℃)要明显小于这一结果。例如Lin等[38]稻田OTC试验发现,在25.6—29.5℃范围内温度每增加1 ℃,IR72单叶光合作用对CO2浓度升高300 μmol/mol的响应平均只增加2.3%。与此不同,在最适温度之上进一步增温对CO2影响的调节效应明显减弱[23,31]。例如Vu等[23]报道,在32—38 ℃范围内,IR72光合速率对长期CO2倍增的响应几乎不受温度变化的影响。与单叶水平不同,CO2浓度和温度对水稻冠层水平光合作用的影响很小,这与大田生长水稻生物量的响应一致[29,49]。Baker等[22]报道,气温在25—37℃区间变化,温度对CO2倍增环境下水稻的冠层光合作用影响很小。大豆试验亦发现CO2浓度与温度的互作对冠层光合作用的影响通常小于单叶水平[50]。一个可能的解释是,CO2浓度升高使稻叶气孔关闭导致冠层温度升高[34],而高温条件下蒸腾降温增强而导致作物冠层温度下降[22]。但与这一假设不同,Bamford等[20]温室试验发现,籼稻IR30移栽后79 d,大气CO2浓度升高使28、34、40℃条件下冠层净光合速率分别增加21%、32%和143%。所以在这一点上还需更多的试验加以确认。
一般来说,短期CO2浓度增加使水稻光合作用增强,但长期生长于CO2熏蒸环境下会出现光合适应或下调现象,但亦有例外[10]。水稻生长温度的改变与光合适应现象有什么关系?Lin等[38]报道,在25.6℃生长温度下,CO2浓度升高对IR72单叶光合速率的促进作用不因生育进程推移而改变,但生长环境增温4℃,光合速率对CO2的响应随时间逐渐变得钝感,表现出明显的光合下调,伴随这一变化的是叶片Rubisco活性或/和浓度下降。Lin等[38]认为这与高温导致的花粉败育使库强减弱有关,后者会限制水稻对碳的利用进而使光合响应的减弱,但其确切的分子机理还需更多的研究。
CO2浓度和温度升高对水稻呼吸作用的影响目前只有一例报道。Cheng等[18]气室试验发现,CO2浓度增加300 μmol/mol使IR72抽穗前、抽穗后夜间呼吸作用分别增加12%和13%,夜温增加10℃使抽穗前和抽穗后分别增加35%和17%,但CO2与温度间没有显著的互作效应。
3 生育进程
生育期是作物的重要性状,通常与产量潜力呈正相关。已有文献表明,CO2浓度或温度升高使水稻抽穗期提前或没有变化,但亦有例外[10,51]。CO2浓度和温度同增试验对水稻生育期响应的报道并不一致。一种观点认为温度越高,高CO2浓度环境下水稻生育期缩短的程度越为明显。Ziska等[51]利用自然光玻璃温室对17个品种的观察发现,所有品种平均,与对照相比,664 μmol/molCO2使29℃和37℃生长温度下播种至抽穗期的天数分别缩短8 d(由97 d缩短至89 d)和10 d(由91 d缩短为81 d)。从不同材料看,9个品种CO2与温度间呈现协同作用,即CO2浓度升高使高温处理水稻抽穗期提前的天数多于适温条件下生长的水稻。日本CTGC试验证实,CO2浓度升高对水稻发育速率的影响明显依赖于生长温度的高低:28、30 ℃条件下,CO2倍增使粳稻Akihikari抽穗之前的天数平均分别缩短6%、11%[29]。但亦有不少研究没有观察到这种协同作用[21-22,25,34,36,38,44]。例如Baker等[22,49]气室试验表明, 34℃ 处理水稻的开花期较28℃ 处理水稻提前10 d,但两种CO2浓度水平趋势一致。Lin等[38]OTC试验发现,增温4℃ 使IR72开花期提前4—5 d,但CO2处理及其温度的互作对开花期均无显著影响。中国T-FACE最新研究表明,CO2浓度升高使杂交稻Ⅱ优084成熟期推迟3 d,但增温及其与CO2处理的互作对生育进程均无影响[44]。综合前人研究,CO2熏蒸水稻生育进程的变化除了与生理响应(如植株氮碳比增加)有关外,可能亦与品种特性和生长环境相关联。
4 分蘖发生
关于大气CO2浓度和温度升高对水稻分蘖发生的影响,文献报道差异很大。日本Kim等[30]利用CTGC设置2个CO2水平和4个温度梯度(区间为26—30℃),结果发现CO2倍增使最高生长温度下粳稻Akihikari的穗数平均增加29%,明显高于其余3个温度水平下穗数对CO2倍增的响应(平均增幅为6%—9%)。韩国CTGC试验亦有类似结果,Yun等[32]报道673 μmol/molCO2使粳稻Ilmybyeo生长前期的分蘖数增加,但增温1.7℃对分蘖数没有影响,CO2浓度和温度同增条件下分蘖数的增幅变得更大,两处理存在微弱的互作效应(P=0.053)。Moya等[36]多年OTC原位试验发现,CO2浓度与温度对分蘖数的影响因品种而异:CO2浓度升高使敏感品种N22常温和高温条件下的分蘖数均增加(低温下增幅更大),IR72分蘖数只在高温条件下才有增加,而钝感品种NPT2分蘖数两个生长温度下均无响应。有趣的是,Baker等[49]气室试验发现,尽管CO2与温度对IR30最终穗数没有互作效应,但对最高分蘖数有明显的交互作用:常温(28℃)下,长期CO2倍增对最高分蘖数没有影响,但高温(34℃和40℃)下CO2倍增导致最高分蘖数明显增加,这一现象可能与该试验温度处理的跨度很大有关。除此之外,亦有一些文献报道CO2与温度对分蘖或最终穗数的影响没有交互作用[18,37]。
5 物质生产与分配
5.1物质生产
CO2浓度和温度的互作对水稻物质生产的影响存在正向、负向和没有三种报道。如前所述,从生化水平看CO2与温度间存在协同作用[48],因此日温升高可能有利于增强CO2浓度升高对作物生长的促进作用。Nakagawa等[12]对文献的分析发现,与大田水稻不同,盆栽水稻生物量对CO2浓度升高的响应明显受气温的影响:温度越高,CO2响应越大[15]。这一现象的可能原因是:盆栽水稻光照条件明显优于大田水稻,因此其分蘖发生的时期延长,特别是在高温条件下。Yun等[32]CTGC试验表明,CO2浓度或温度升高均使Ilmybyeo生物量显著增加,两处理间存在显著的正向互作:CO2浓度升高使穗分化始期、齐穗期和成熟期生物量平均分别增加15%、9%和2%,同时增温条件下则分别增加39%、31%和8%。Baker等[49]报道,尽管CO2与温度对IR30最终生物量没有互作效应,但对最大生物量表现出明显的正向互作:28、34、40℃温度下,CO2倍增使最大生物量分别增加8%、34%和52%。
与此相反,另一种观点认为高温生长环境使水稻生物量对CO2浓度升高的响应明显下调。Ziska等[51]气室试验表明,所有17个品种平均,664 μmol/mol CO2使29、37℃生长温度下全株生物量平均分别增加了70%、22%,不同器官表现出类似趋势:以根系响应最大(142%对59%,低温对高温,下同),从大到到小依次为分蘖重(97%对27%)、穗重(54%对49%)、叶重(46%对5%)。Moya等[36]报道供试品种IR72和N22表现出类似趋势,例如CO2浓度增加200、300 μmol/mol使常温下N22总生物量分别增加29%和59%,高温(+4℃)下则没有响应。近期的夜温升高试验亦发现,水稻生殖生长期增温使CO2浓度升高对水稻生长的增益效应减少:低温(22℃)和高温(32℃)条件下,CO2浓度升高使IR72植株干重分别增加38%和13%[18]。
更多的文献报道,水稻生物量对高CO2浓度的响应不受生长温度的影响。Ziska等[37]OTC试验表明,CO2浓度增加200、300 μmol/mol使IR72成熟期总生物量平均分别增加31%和40%,但CO2与温度间互作没有互作效应。Cheng等[18]报道,生殖生长期CO2浓度增加300 μmol/mol和夜温增加10℃均使全株和茎鞘干重显著增加,但两处理间不存在互作效应。日本科学家对长期CO2熏蒸试验的分析发现[12,29],CO2倍增使近似大田生长水稻Akihikari的物质生产量平均增加24%,但这一增幅比较稳定,几乎不受温度变化的影响:温度每增1℃,CO2响应值平均增加1.8%。
可见,物质生产对CO2浓度和温度的响应不能简单地根据光合速率的响应来预测,因为水稻碳氮平衡、生育进程、叶片扩展对CO2和温度互作的响应不同,而物质积累量正是这些过程共同作用的结果[52]。CO2与温度的交互作用并不一定表现为协同效应,说明生化过程并不总会在全株水平上表现出来。
5.2物质分配
大气CO2浓度和温度升高对水稻物质分配的影响报道很少而且结果不一致。收获指数(HI)反映了作物光合产物向结实器官运转的效率。Baker等[49]报道,高温处理使不同CO2水平下IR30的HI均明显下降,但CO2与温度间没有互作效应。日本温度梯度(36—40℃)试验发现,CO2倍增使最低生长温度下水稻HI略增(+5%),但使其余3个生长温度下(从低到高)分别减少了17%、42%和56%(Kim等[30])。Ziska等[37]OTC试验表明,CO2与温度的互作对供试品种HI的影响因年度而异。Moya等[36]报道CO2浓度升高对常温下供试品种HI没有影响,但使高温环境下生长的水稻HI多呈下降趋势。Ainsworth[53]对上述气室研究的整合分析发现,CO2浓度升高使无胁迫环境下生长的水稻HI平均增加11%(n=18),但在高温胁迫下(n=5)这种促进作用完全消失。根冠比是反映同化产物在稻体内分配的重要指标。Kim等[29]两年CTGC试验发现,CO2倍增对Akihikari成熟期根干重与总干重之比呈增加趋势,但生长温度过高过低增幅均减少,而Ziska等[37]两季OTC试验发现,CO2浓度升高使不同温度处理水稻成熟期的根冠比均显著增加,但两处理间没有互作效应。
6 籽粒产量
与生物量一样,CO2浓度和温度对水稻最终产量的影响存在协同、抑制和独立三种情形。一种观点认为CO2和温度会协同作用于水稻,从而发挥增产潜力,但目前只有1篇相关文献。日本FACE研究发现[54],2003年低温寡照条件下,CO2浓度升高使水稻产量平均只增加了6%,而2004年高温多照条件下(全生育期气温比2003年高1.5℃),同一品种增产幅度高达17%,CO2和年度间存在显著的互作效应,推测在低温地区,适当增温可增加CO2浓度升高对水稻产量的肥料效应。
另一种观点认为CO2浓度和温度升高对水稻产量的影响相对独立。Baker等[49]温室试验发现,CO2倍增使28℃和34℃生长温度下IR30的籽粒产量略增,但均未达显著水平,这主要与试验期寡照有关。该试验高温处理对产量影响很大,但其与CO2浓度没有互作效应。Yun等[32]报道,673 μmol/mol CO2使Ilmybyeo穗重增加17%,增温1.7℃使穗重减少12%,两者同时升高条件下产量无显著变化,CO2与温度间没有互作效应。Madan等[25]发现,开花期短期高温使供试品种N22籽粒产量显著下降,但CO2处理本身及其与温度的互作均无显著影响。近期OTC试验表明,与对照相比,CO2浓度和温度同时增高下水稻的增产幅度与两因子单独升高下增产幅度相近[41-42],处理间无相互协同或抑制的迹象。
与此不同,大量水稻文献报道了高温对CO2肥料效应的抑制作用。20世纪90年代的温室[51]、CTGC[30]和OTC试验[36,37]均表明,高温可以部分甚至全部抵消大气CO2浓度升高对水稻产量的增益效应。Kim等[29]温度梯度(26—30℃)试验发现,在最低温度下,CO2倍增使1991和1992年Akihikari糙米产量平均分别增加45%和20%,但使最高生长温度下分别减产15%和91%。国际水稻研究所(IRRI)Lin等[38]单季OTC试验发现,CO2浓度增加200、300 μmol/mol使25.6℃生长温度下籼稻IR72穗重分别增加48%和51%,而在增温4℃条件下只分别增加27%和39%。利用同样的平台、品种和处理,Ziska等[37]两季试验发现,CO2浓度增加200、300 μmol/mol使常温下籽粒产量分别增加15%和27%,但增温4℃条件下这种肥料效应几乎消失。这一时期IRRI其它试验亦验证了这一结果[26,36,39,51]。进入21世纪,Cheng等[18]夜温增高试验亦发现了CO2与温度间的负向互作:CO2浓度升高使低温和高温处理水稻的糙米产量分别增加27%和9%。Madan等[25]近期气室试验发现,生长温度为29℃时,CO2浓度升高使籼稻IR64和杂交稻IR75217H籽粒产量显著增加,但当开花期温度升至35℃和38℃时,这种增益效应完全消失。以上研究表明,尽管CO2浓度升高增加籽粒产量,但如果同时暴露在高温(包括夜温)环境下则会减少甚至抵消CO2的肥料效应。一般认为这可能与高温使水稻育性下降,进而使高浓度CO2环境下增加的光合产物无法顺利转运至籽粒有关[18,29,37]。CO2,温度间广泛存在的负向互作对未来包括当前气候条件下的稻作生产均是一个严重威胁。
水稻最终产量由整个生命周期的不同生长过程所决定,这些过程对CO2浓度和温度增高的潜在响应可能并不相同并且存在基因型差异,这可能是CO2与温度间交互作用的程度甚至方向变异较大的重要原因[52]。另外,增温幅度特别是关键生长期(尤其是开花期最高气温)增温幅度的不同可能也是导致结果不一致的重要原因。Figueiredo等[43]最新大田OTC试验发现,在温度较低的2011年,CO2浓度和温度同增条件下直播稻Ariete产量明显增加,CO2浓度升高表现出对高温伤害的补偿效应,而2011年生殖生长期最高温度多超过34℃,两因子同时升高导致该品种大幅减产。
7 受精过程
大量研究表明,水稻对温度最敏感的时期是孕穗至抽穗扬花期[55]。众所周知,因授粉失败导致的小穗败育通常因花粉小孢子发育阶段或开花期日平均温度低于20℃[24],或开花期温度高于33—35℃而造成[55-57]。对水稻而言,白天高温可能会导致花药不开裂、花药授粉量减少[34,57]、花粉管萌发受阻,进而导致受精率和结实率减少。当然在更高温度下,雌性不育亦会发生[55]。可见,花粉是水稻在高温条件下最易受伤害的器官之一,但目前只有一例关于CO2浓度与温度互作对花粉育性的报道。
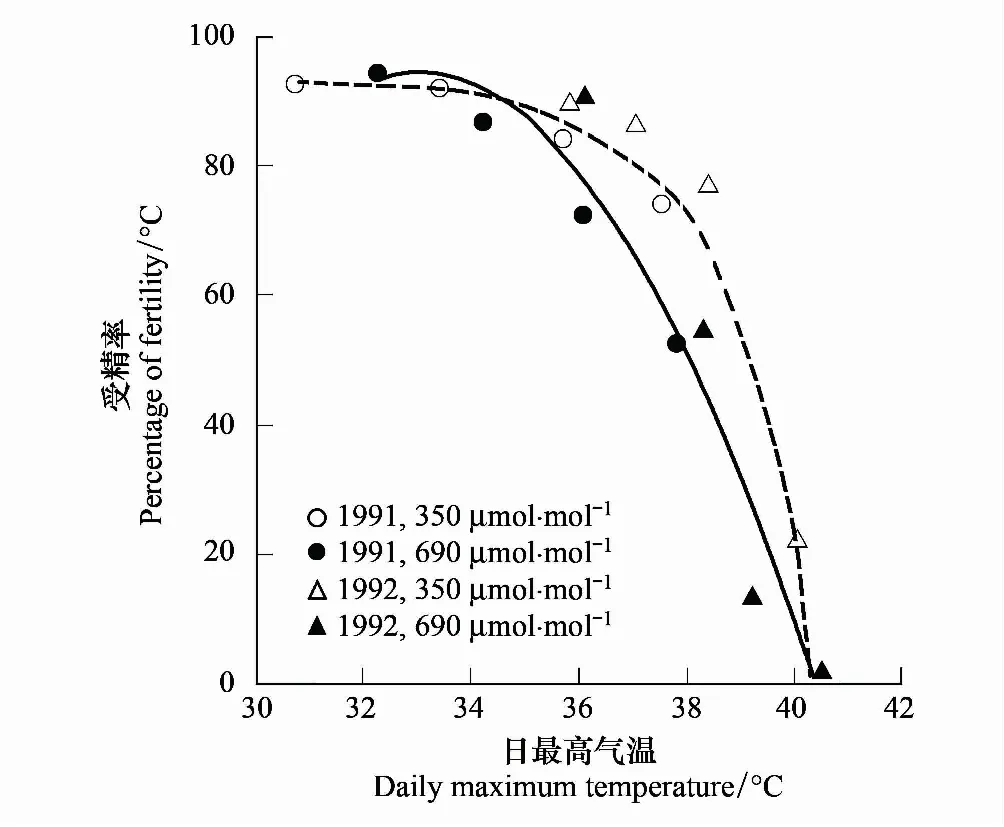
图2 环境CO2浓度(○,△)和高CO2浓度下(●,▲)水稻受精率与开花期平均最高气温的关系(1991和1992年)Fig.2 Percentage of fertility as a function of daily maximum air temperature averaged over flowering period for rice grown under ambient (○,△) and elevated (●,▲) CO2 concentrations in 1991 and 1992数据来自日本粳稻Akihikari的CTGC数据. Data are from japonica rice cv. Akihikari grown in CTGC in Japan
IRRI大田OTC发现[34],开花授粉期温度升高4℃使水稻IR72败育花粉增多,如果CO2浓度同时增加(+300 μmol/mol),花粉败育程度明显加剧。研究还发现,CO2浓度升高使小穗不育(由柱头上的花粉萌发数量而定)的临界气温降低了1℃等。Matsui等[34]推测,临界温度下调可能与观察到的CO2熏蒸水稻冠层空气和穗温升高的间接作用有关,而后者可能又与高CO2浓度下气孔关闭和蒸腾降温减少有关。与这一假设一致,迄今确有不少试验观察到这种负向互作对水稻受精率或结实率的影响[30,34,38,37]。Kim等[30]两年温度梯度试验发现,CO2倍增对最低生长温度下水稻的受精率均无显著影响,但使其余3个温度处理下(从低到高)分别下降6%、14%和29%(1991年)和36%、82%和88%(1992年),受精率降幅与开花期平均最高气温关系密切(图2)。Lin等[38]亦发现,200 和300 μmol/molCO2对常温下IR72的受精率均无影响,但使高温下(+4℃)结实率分别下降4.7%、12.4%。当然亦有不同报道[18,25,36],如Madan等[25]报道,CO2倍增特别是开花短期高温(5 d)均使供试品种结实率下降,但两者没有交互作用。Cheng等[18]气室试验表明,夜温升至32℃导致的受精率和结实率降低与CO2间亦不存在互作效应。
必须指出的是,结实率或受精率对CO2浓度和温度互作的响应与花粉育性的变化并非完全一致。因为,CO2-温度升高条件下结实率的响应同时可能还与花药开裂、花粉萌发、花粉管伸长等重要过程是否受阻等有关;其次,虽然高温胁迫下部分花粉育性下降,但其他可育花粉仍然可能完成授粉作用。另外,其它一些环境因子(如风速、湿度)也被发现影响高温下小穗败育程度[57]。因此,由于这一机制的复杂性[55,57],上述交互作用的确切机理需更多试验来阐明。
8 碳氮代谢
有关高温和高CO2浓度条件下水稻碳氮积累及其在不同器官中分配变化的试验数据非常有限。Cheng等[19]首次报道了生殖生长期CO2和夜温处理对水稻碳同化、氮吸收及其分配的影响。盆栽试验表明,CO2浓度或夜温升高对IR72全株的碳、氮浓度均无显著影响,但使全株碳同化量(包括碳的净增量和新碳量)、氮吸收量均显著增加,且两处理间存在显著的互作效应,表现在夜间增温使CO2浓度升高对处理期全株碳同化和氮吸收的促进作用减小。尽管夜温升高使夜间呼吸增强导致碳损失增加,高温使两个CO2水平下水稻的碳同化均增加,这与夜温升高使绿叶氮浓度(+7.8%)、单位干重叶面积显著增加(+14.5%)进而导致生长后期光合能力增强有关。该研究还发现,CO2浓度对生殖生长期稻穗和茎鞘间碳、氮分配没有影响,但夜温升高明显减少茎鞘中碳、氮向稻穗的分配。结合该试验的产量响应可知[19],由于夜温升高限制碳、氮向稻穗的转运,因此CO2浓度和夜温升高并存情形下,高温会减少CO2对产量的肥料效应,这可能会直接威胁未来的稻作生产。
水稻物质分配与碳水化合物在植株体内的转化和转移有关。淀粉和蔗糖是作物光合和生长的主要中间贮藏产物。Bamford等[20]日光型温室试验发现,尽管高温对播种后79 d叶片蔗糖浓度没有影响,但CO2倍增使播种后79 d叶片蔗糖浓度显著增加:28、34、40℃生长温度下分别增加11%、23%和37%。与此对应,CO2倍增使34℃和40℃生长温度下白天叶片蔗糖累积速率显著增加,但对28℃条件下没有影响。CO2浓度与温度的互作对叶片和稻穗总的非结构性碳水化合物(TNC)浓度没有影响,但对茎鞘TNC浓度有显著影响:高温胁迫下CO2效应更大,特别是生长中后期。研究还发现,CO2浓度升高使28、34℃处理水稻单位叶重生产的碳水化合物总量无显著影响,但使40℃生长温度下大幅增加(+116%)。这些数据说明,大气CO2浓度升高能使遭受高温胁迫的水稻维持碳水化合物的生产速率,进而减轻高温对营养生长的一些负面影响。对同一品种的观察发现,40℃高温胁迫下,生长在330 μmol/molCO2环境下的水稻在茎伸长期就已死亡,而660 μmol/mol熏蒸水稻得以存活,尽管稻穗没有结实[20,49]。
9 稻米品质
水稻灌浆阶段即是籽粒不断增加淀粉、蛋白质、脂肪和其它营养物质的过程[58],因此既是产量也是品质形成过程。该阶段最适的日均温为20—25℃,明显低于其它生长发过程的最适温度[56]。高CO2浓度环境下生长的水稻同化产物较为充裕,推测可能缓解高温引起的粒重下降。但实验数据并非如此。多数文献报道CO2与温度对粒重不存在交互作用[18,25,36-37,49]。与此不同,Kim等[30]CTGC试验发现,CO2倍增对较低生长温度下水稻千粒重的影响均无显著影响,但使最高温度处理水稻的千粒重平均下降了27%(约4.5 g)。
已有综述表明高浓度CO2环境下稻米加工和外观品质多呈变劣趋势[59],这种现象是否因灌浆期温度改变而异?谢立勇等[60]CTGC试验表明,同步增加CO2浓度与大气温度对优质粳稻中作93稻米加工和外观品质的负面影响最大:表现在糙米率、精米率和整精米率均呈明显下降的趋势,而垩白率和垩白度增加,但遗憾的是该试验没有设置单独的CO2和温度处理。日本最新FACE数据表明,高温可能会恶化CO2浓度升高环境下稻米加工和外观品质变劣的程度[61],研究发现CO2浓度升高使5个热敏品种完整米百分率平均减少10个百分点、腹白和背白粒率平均增加了8.7个百分点,而7个耐热品种这些参数均无显著变化。年度比较发现,高温生长季使热敏品种高浓度CO2导致的碎米明显增多, 而耐热品种CO2熏蒸导致的碎米不同年度均较低(Usui等[61]图2)。与此不同,Madan等[25]气室短期高温试验发现,CO2倍增使3个供试品种籽粒宽度和完整米百分率均显著增加,但高温及其与CO2的互作均无显著影响。对粒长和垩白而言,CO2、温度处理以及两者的互作均无显著影响。稻米淀粉颗粒形成的高峰期一般发生在花后5d之后,这可能是该试验短期高温胁迫没有导致垩白变化的主要原因[7]。
直链淀粉决定稻米的粘性。Ziska等[37]OTC试验表明,CO2浓度升高对稻米直链淀粉无影响,但高温下直链淀粉浓度增加或没有影响。谢立勇等[60]发现随着CO2浓度和温度的升高呈先降后增的趋势,而胶稠度刚好相反。Madan等[25]报道,开花期短期高温使供试品种稻米直链淀粉含量显著下降,且在高CO2浓度环境下更为明显。该试验还发现,CO2浓度、温度及其互作对耐高温品种N22稻米胶稠度没有影响,但对杂交稻IR75217H和籼稻IR64均有显著影响:高温胁迫只使高CO2浓度环境下生长的稻米胶稠度显著下降。品质较差是目前影响杂交稻推广的一个主要瓶颈,未来气候条件下品质更差将影响杂交稻的商业化应用,进而可能对粮食安全产生严重影响。因此,强化这方面的研发已势在必行。
10 水稻/杂草竞争关系
随着大气CO2浓度升高,一个可以预见的结果是重要作物(多数为C3作物如水稻)对杂草(多数是C4作物)相对的竞争力增强,但气温升高却可能对C4植物更加有利。目前CO2和温度互作对水稻与杂草竞争关系的影响只有一例报道。Alberto等[26]采用气室水培试验设置不同种植比例的水稻和稗草,研究发现,昼夜温度为27/21℃时,所有种植比例平均,594 μmol/molCO2浓度使IR72地上部生物量和籽粒产量平均分别增加47%和55%,但稗草的生物量和籽粒产量均无显著变化。在两物种混植小区,CO2浓度升高使水稻相对于稗草生物量的比例显著增加,不同混植比例处理趋势一致。两物种比较,CO2浓度升高使水稻的竞争优势增加,而稗草的竞争力减弱。然而在37/29℃条件下,CO2浓度升高对水稻营养和生殖生长的促进作用减少,这导致所有间植方式下水稻和稗草生物量之比较27/21℃下降,水稻和稗草产量之比的降幅更大。该试验表明,单独CO2浓度升高情形下水稻相对于稗草的竞争力增强,然而CO2浓度和温度同时升高的条件下,稗草的竞争力更强。这对未来气候条件水稻的种植提出了更高的要求。由于其他因子如光、水、养分以及其它类型杂草等的变化常影响作物与稗草的相对竞争力,因此有必要进一步研究开放稻田条件下水稻与杂草竞争关系的变化及其机理。
11 展望
综观前人研究,尽管该领域已经取得了一些重要进展特别在生育期、生物量和籽粒产量方面,但很多方面还没有或很少涉及,而且已有结果并不一致甚至矛盾。未来研究无论从研究内容的深入和拓宽上,还是研究方法(手段)的改进和创新上,均有很大空间。
(1)从研究手段看,目前该方向的认知多来自于封闭或半封闭气室的研究,这些结果能否在开放大田重演还需在土壤-植物-大气连续体扰动较小的实验平台的验证。2013年中国科学院朱建国研究员及其同事将原有稻田CO2-FACE系统改建为T-FACE试验平台(图1),模拟本世纪中叶的气候变化情景(即目前被广泛引用的CO2倍增后气温将上升2—2.5℃的情形[5]),开展CO2浓度和气温同时升高对水稻影响的开放式实验研究。这一新技术为开展自然条件下气候变化对水稻影响的多因子操作试验开辟了新途径,有利于推动我们对于许多未解问题的探索。
(2)未来需要强化的研究内容,主要包括种子萌发、幼苗生长、水分关系、根系发育、根际过程、源库平衡、颖花形成、花粉育性、籽粒灌浆、品质形成、养分吸收利用、世代响应、冠层微气候以及逆境抗性(如病虫草害)。为了准确揭示这些过程的响应特征及其生物学机制,需大力推进多学科交叉融合,包括作物栽培、生理、生化、生态、遗传、土壤和农田微气象等,方能看清假定气候情景对水稻影响的全貌。
(3)水稻生长在多变的栽培和环境条件下,在研究CO2浓度-温度增加对水稻影响的同时,必须考虑其他重要外界因子(包括生物和非生物因子)的协同效应,这样才具有更实际的生态学意义。前期研究表明CO2与温度间的交互作用受供试品种的影响[25,36],但不同类型水稻(如不同亚种间水稻、早中晚稻等)的响应存在多大差异目前尚无直接证据。另外,CO2浓度-温度互作是否以及多大程度上受其它重要生物和非生物因子的影响更不清楚。目前迫切需要了解包括种植期、肥料运筹、水分管理、桔杆还田甚至种植制度等对这一交互作用的调控效应,这些信息对开发出适应未来气候变化的稻作生产技术是必需的。最新出现的T-FACE系统,为我们提供了一个非常好的机会来推进这项工作。
(4)大气CO2浓度的变化是全球均一的,但气温上升存在明显的时空变化[5]。尽管长时间CO2和温度处理对水稻影响的研究较多[21-22,24,29-30,32,34,38,51],但更加切合实际的开花或灌浆期高温与不同CO2水平间的互作仍需加强研究。另一方面,全球变暖过程中夜温增加的速度快于昼温增加的速度[5,62],前者已对全球水稻生产力构成威胁[63]。因此,加强夜温与CO2互作对水稻影响的研究很有必要[18]。
[1]IPCC (Intergovernmental Panel on Climate Change). The Physical Science Basis // Lisa V A, Simon K A, Nathaniel L B, eds. Contribution of Working GroupⅠto the Fifth Annual Assessment Report of the IPCC. Cambridge UK: Cambridge University Press, 2013: 29-29.
[2]Idso K E, Idso S B. Plant responses to atmospheric CO2enrichment in the face of environmental constraints: a review of the past 10 years′ research. Agricultural and Forest Meteorology, 1994, 69(3/4): 153-203.
[3]Kimball B A. Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agronomy Journal, 1983, 75(5): 779-788.
[4]Schönwiese C. Houghton J T, Meira Filho L G, Callander B A, Harris N, Kattenberg A, Maskell K, (eds.). Climate Change 1995, The Science of Climate Change. UK, Cambridge: Cambridge University Press, 1996.
[5]IPCC (Intergovernmental Panel on Climate Change). Climate Change 2007: The Physical Science Basis // Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L, eds. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK/New York: Cambridge University Press, 2007.
[6]Wassmann R, Jagadish S V K, Sumfleth K, Pathak H, Howell G, Ismail A, Serraj R, Redona E, Singh R K, Heuer S. Regional vulnerability of climate change impacts on Asian rice production and scope for adaptation. Advances in Agronomy, 2009, 102(5): 91-133.
[7]Fitzgerald M A, Resurreccion A P. Maintaining the yield of edible rice in a warming world. Functional Plant Biology, 2009, 36(12): 1037-1045.
[8]Tilman D, Balzer C, Hill J, Befort B L. Global food demand and the sustainable intensification of agriculture. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(50): 20260-20264.
[9]杨连新, 王云霞, 朱建国, 王余龙. 十年水稻FACE研究的产量响应. 生态学报, 2009, 29(3): 1486-1497.
[10]杨连新, 王云霞, 朱建国, Hasegawa T, 王余龙. 开放式空气中CO2浓度增高(FACE)对水稻生长和发育的影响. 生态学报, 2010, 30(6): 1573-1585.
[11]段骅, 杨建昌. 高温对水稻的影响及其机制的研究进展. 中国水稻科学, 2012, 26(4): 393-400.
[12]Nakagawa H, Horie T. Rice responses to elevated CO2and temperature. Global Environmental Research, 2000, (3): 101-113.
[13]Long S P, Ainsworth E A, Leakey A D B, Nösberger J, Ort D R. Food for thought: lower-than-expected crop yield stimulation with rising CO2concentrations. Science, 2006, 312(5782): 1918-1921.
[14]Ishii R, Ohsugi R, Murata Y. The effect of temperature on the rates of photosynthesis, respiration and the activity of RuDP carboxylase in barley, rice and maize leaves. Japanese Journal of Crop Science, 1977, 46(4): 516-523.
[15]Imai K, Coleman D F, Yanagisawa T. Increase in atmospheric partial pressure of carbon dioxide and growth and yield of rice (OryzasativaL.). Japanese Journal of Crop Science, 1985, 54(4): 413-418.
[16]Imai K, Okamoto-Sato M. Effects of temperature on CO2dependence of gas exchanges in C3and C4crop plants. Japanese Journal of Crop Science, 1991, 60(1): 139-145.
[17]Cheng W G, Sakai H, Hartley A, Yagi K, Hasegawa T. Increased night temperature reduces the stimulatory effect of elevated carbon dioxide concentration on methane emission from rice paddy soil. Global Change Biology, 2008, 14(3): 644-656.
[18]Cheng W G, Sakai H, Yagi K, Hasegawa T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agricultural and Forest Meteorology, 2009, 149(1): 51-58.
[19]Cheng W G, Sakai H, Yagi K, Hasegawa T. Combined effects of elevated [CO2] and high night temperature on carbon assimilation, nitrogen absorption, and the allocations of C and N by rice (OryzasativaL.). Agricultural and Forest Meteorology, 2010, 150(9): 1174-1181.
[20]Rowland-Bamford A J, Baker, J T, Allen L H Jr, Bowes G. Interactions of CO2enrichment and temperature on carbohydrate accumulation and partitioning in rice. Environmental and Experimental Botany, 1996, 36(1): 111-124.
[21]Baker J T, Allen L H Jr, Boote K J. Temperature effects on rice at elevated CO2concentration. Journal of Experimental Botany, 1992, 43(7): 959-964.
[22]Baker J T, Allen L H Jr. Effects of CO2and temperature on rice: a summary of five growing seasons. Journal of Agricultural Meteorology, 1993, 48(5): 575-582.
[23]Vu J C V, Allen L H Jr, Boote K J, Bowes G. Effects of elevated CO2and temperature on photosynthesis and Rubisco in rice and soybean. Plant, Cell and Environment, 1997, 20(1): 68-76.
[24]Baker J T. Yield responses of southern US rice cultivars to CO2and temperature. Agricultural and Forest Meteorology, 2004, 122(3/4): 129-137.
[25]Madan P, Jagadish S V K, Craufurd P Q, Fitzgerald M, Lafarge T, Wheeler T R. Effect of elevated CO2and high temperature on seed-set and grain quality of rice. Journal of Experimental Botany, 2012, 63(10): 3843-3852.
[26]Alberto A M P, Ziska L H, Cervancia C R, Manalo P A. The influence of increasing carbon dioxide and temperature on competitive interactions between a C3crop, rice (Oryzasativa) and a C4weed (Echinochloaglabrescens). Australian Journal of Plant Physiology, 1996, 23(6): 795-802.
[27]Horie T, Nakano J, Nakagawa H, Wada K, Kim H Y, Seo T. Effect of elevated CO2, and high temperature on growth and yield of rice. 1. Development of temperature gradient tunnels. Japanese Journal of Crop Science, 1991, 60(extra issue 2): 127-128.
[28]Horie T, Nakagawa H, Nakano J, Homotani K, Kim H Y. Temperature gradient chambers for research on global environment change. Ⅲ. A system designed for rice in Kyoto, Japan. Plant, Cell & Environment, 1995, 18(9): 1064-1069.
[29]Kim H Y, Horie T, Nakagawa H, Wada K. Effects of elevated CO2concentration and high temperature on growth and yield of rice. I. The effect on development, dry matter production and some growth characteristics. Japanese Journal of Crop Science, 1996, 65(4): 634-643.
[30]Kim H Y, Horie T, Nakagawa H, Wada K. Effects of elevated CO2concentration and high temperature on growth and yield of rice. Ⅱ. The effect on yield and its components of Akihikari rice. Japanese Journal of Crop Science, 1996, 65(4): 644-651.
[31]Nakagawa H, Horie T, Kim H Y, Ohnishi H, Homma K. Rice responses to elevated CO2concentrations and high temperatures. Journal of Agricultural Meteorology, 1997, 52(5): 797-800.
[32]Yun S I, Kang B M, Lim S S, Choi W J, Ko J, Yoon S, Ro M H, Kim H Y. Further understanding CH4emissions from a flooded rice field exposed to experimental warming with elevated [CO2]. Agricultural and Forest Meteorology, 2012, 154-155: 75-83.
[33]谢立勇, 林而达, 赵海燕, 孙芳, 仝乘风. 基于改进的CTGC系统下不同CO2浓度对水稻生长发育的影响. 中国农业通报, 2006, 22(8): 139-143.
[34]Matsui T, Namuco O S, Ziska L H, Hori T. Effects of high temperature and CO2concentration on spikelet sterility in indica rice. Field Crops Research, 1997, 51(3): 213-219.
[35]Moya T B, Ziska L H, Weldon C, Quilang J E P, Jones P. Microclimate in open-top chambers: Implications for predicting climate change effects on rice production. Transactions of the ASABE, 1997, 40(3): 739-747.
[36]Moya T B, Ziska L H, Namuco O S, Olszyk D. Growth dynamics and genotypic variation in tropical, field-grown paddy rice (OryzasativaL.) in response to increasing carbon dioxide and temperature. Global Change Biology, 1998, 4(6): 645-656.
[37]Ziska L H, Namuco O, Moya T, Quilang J. Growth and yield response of field-grown tropical rice to increasing carbon dioxide and air temperature. Agronomy Journal, 1997, 89(1): 45-53.
[38]Lin W H, Ziska L H, Namuco O S, Bai K Z. The interaction of high temperature and elevated CO2on photosynthetic acclimation of single leaves of rice in situ. Physiologia Plantarum, 1997, 99(1): 178-184.
[39]Olszyk D M, Centeno H G S, Ziska L H, Kern J S, Matthews R B. Global climate change, rice productivity and methane emissions: comparison of simulated and experimental results. Agricultural and Forest Meteorology, 1999, 97(2): 87-101.
[40]万运帆, 游松财, 李玉娥, 王斌, 高清竹, 秦晓波, 刘硕. 开顶式气室原位模拟温度和CO2浓度升高在早稻上的应用效果. 农业工程学报, 2014, 30(5): 123-130.
[41]万运帆, 游松财, 李玉娥, 王斌, 高清竹, 秦晓波, 刘硕. CO2浓度和温度升高对早稻生长及产量的影响. 农业环境科学学报, 2014, 33(9): 1693-1698.
[42]Roy K S, Bhattacharyya P, Neogi S, Rao K S, Adhya T K. Combined effect of elevated CO2and temperature on dry matter production, net assimilation rate, C and N allocations in tropical rice (OryzasativaL.). Field Crops Research, 2012, 139: 71-79.
[43]Figueiredo N, Carranca C, Trindade H, Pereira J, Goufo P, Coutinho J, Marques P, Maricato R, de Varennes A. Elevated carbon dioxide and temperature effects on rice yield, leaf greenness, and phenological stages duration. Paddy and Water Environment, 2014, doi 10.1007/s10333-014-0447-x.
[44]赖上坤, 庄时腾, 吴艳珍, 王云霞, 朱建国, 杨连新, 王余龙. 大气CO2浓度和温度升高对超级稻生长发育的影响. 生态学杂志, 2015, 34(5): 1253-1262.
[45]Hendrey G R. Free-air Carbon Dioxide Enrichment for Plant Research in the Field. Boca Raton, FL: Smoley, 1993.
[46]Okada M, Lieffering M, Nakamura H, Yoshimoto M, Kim H Y, Kobayashi K. Free-air CO2enrichment (FACE) using pure CO2injection: system description. New Phytologist, 2001, 150(2): 251-260.
[47]刘钢, 韩勇, 朱建国, 冈田益己, 中村浩史, 吉本真由美. 稻麦轮作FACE 系统平台I. 系统结构与控制. 应用生态学报, 2002, 13(10): 1253-1258.
[48]Long S P. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2concentrations: Has its importance been underestimated? Plant, Cell & Environment, 1991, 14(8): 729-739.
[49]Baker J T, Allen L H Jr, Boote K J. Response of rice to carbon dioxide and temperature. Agricultural and Forest Meteorology, 1992, 60(3/4): 153-166.
[50]Jones P, Allen L H, Jones J W. Responses of soybean canopy photosynthesis and transpiration to whole-day temperature changes in different CO2environments. Agronomy Journal, 1985, 77(2): 242-249.
[51]Ziska L H, Manalo P A, Ordonez R A. Intraspecific variation in the response of rice (OryzasativaL.) to increased CO2and temperature: growth and yield response of 17 cultivars. Journal of Experimental Botany, 1996, 47(9): 1353-1359.
[52]Morison J I L, Lawlor D W. Interactions between increasing CO2concentration and temperature on plant growth. Plant, Cell & Environment, 1999, 22(6): 659-682.
[53]Ainsworth E A. Rice production in a changing climate: a meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration. Global Change Biology, 2008, 14(7): 1642-1650.
[54]Shimono H, Okada M, Yamakawa Y, Nakamura H, Kobayashi k, Hasegawa T. Rice yield enhancement by elevated CO2is reduced in cool weather. Global Change Biology, 2008, 14(2): 276-284.
[55]Statake T, Yoshida S. High temperature-induced sterility in indica rices at flowering. Japanese Journal of Crop Science, 1978, 47(1): 6-17.
[56]Yoshida S, Satake T, Mackill D J. High temperature stress in rice // IRRI Research Paper Series 67. Manila, Philippines: International Rice Research Institute, 1981.
[57]Matsui T, Omasa T, Hori T. High temperature-induced spikelet sterility of japonica rice at flowering in relation to air temperature, humidity and wind velocity conditions. Japanese Journal of Crop Science, 1997, 66(3): 449-455.
[58]Jablonski L M, Wang X Z, Curtis P S. Plant reproduction under elevated CO2conditions: A meta-analysis of reports on 79 crop and wild species. New Phytologist, 2002, 156(1): 9-26.
[59]Wang Y X, Frei M, Song Q L, Yang L X. The impact of atmospheric CO2concentration enrichment on rice quality—A research review. Acta Ecologica Sinica, 2011, 31(6): 277-282.
[60]谢立勇, 马占云, 韩雪, 林而达. CO2浓度与温度增高对水稻品质的影响. 东北农业大学学报, 2009, 40(3): 1-6.
[61]Usui Y, Sakai H, Tokida T, Nakamura H, Nakagawa H, Hasegawa T. Heat-tolerant rice cultivars retain grain appearance quality under free-air CO2enrichment. Rice, 2014, 7(1): 6-9.
[62]Easterling D R, Horton B, Jones P D, Peterson T C, Karl T R, Parker D E, Salinger M J, Razuvayev V, Plummer N, Jamason P, Folland C K. Maximum and minimum temperature trends for the globe. Science, 1997, 277(5324): 364-367.
[63]Peng S B, Huang J L, Sheehy J E, Laza R C, Visperas R M, Zhong X H, Centeno G S, Khush G S, Cassman K G. Rice yields decline with higher night temperature from global warming. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(27): 9971-9975.
Combined effect of increasing atmospheric CO2concentration and temperature on growth and development of rice: a research review
JING Liquan1, LAI Shangkun1, WANG Yunxia2, YANG Lianxin1,*, WANG Yulong1
1JiangsuKeyLaboratoryofCropGeneticsandPhysiology,Co-InnovationCenterforModernProductionTechnologyofGrainCrops,YangzhouUniversity,Yangzhou225009,China2CollegeofEnvironmentalScienceandEngineering,YangzhouUniversity,Yangzhou225009,China
The increases in air temperature and atmospheric carbon dioxide (CO2) concentrations are the two most important attributes of global climate change. CO2concentrations have increased from 280 μmol/mol in 1800 to 396 μmol/mol at present, and in the worst case scenario, it is estimated to reach 936 μmol/mol by the end of this century. Concomitant with the enhanced greenhouse effect caused by the increasing concentrations of CO2and other greenhouse gases, the projected rise in the global average surface air temperature before 2100 relative to 1980—1999 is about 1.8—4.0℃. Rice is one of the most important food crops for more than half of the world′s population. In this review, we introduced the experimental platforms that have been used to study the effects of rising temperatures and CO2comcentrations on crops. Subsequently, the experimental progress achieved so far was summarized systematically, highlighting the effects of two important environmental factors—temperature and CO2concentration—on rice growth and development. The factors studied include photosynthesis, development process, tillers formation, biomass production, grain yield, process of fertilization, carbon and nitrogen metabolism, rice quality and rice/weed competition etc. As the main substrate for photosynthesis, the elevated concentrations of atmospheric CO2exhibited direct beneficial effects on rice productivity in most cases. In contrast, the effects of temperature rise, or its combined effect with higher CO2concentrations, on the rice growth process varied substantially (from negative to positive). This reflects the complex relationship between the treatment factors (including the CO2, temperature treatment level and duration) and the varieties and growth conditions of rice. Until now, our knowledge in this field has mainly been obtained from the studies conducted in closed or semi-closed gas chambers. The focus of future research will be the use of riceT-FACE (Temperature-Free Air CO2Enrichment), in combination with chamber facilities, to carry out further multidisciplinary research. The ultimate goal will be to understand the effects of temperature, CO2interactions on the key processes of rice growth and the biological mechanisms involved in such interactions to improve our ability in predicting the variation in rice growth with changing climatic conditions and to develop more effectively the adaptation strategies to cope with climate change.
rice; climate change; carbon dioxide (CO2); temperature; growth and development
国家自然科学基金面上项目(31371563, 31171460, 31571597, 31471437);江苏省高校自然科学重大基础研究项目(11KJA210003);江苏省博士后科研资助计划(1501077C);江苏高校优势学科建设工程资助项目
2014-12-27; 网络出版日期:2015-10-30
Corresponding author.E-mail: lxyang@yzu.edu.cn
10.5846/stxb201412272588
景立权,赖上坤,王云霞,杨连新,王余龙.大气CO2浓度和温度互作对水稻生长发育的影响.生态学报,2016,36(14):4254-4265.
Jing L Q, Lai S K, Wang Y X, Yang L X, Wang Y L.Combined effect of increasing atmospheric CO2concentration and temperature on growth and development of rice: a research review.Acta Ecologica Sinica,2016,36(14):4254-4265.
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27
今日农业(2022年16期)2022-11-09
今日农业(2022年15期)2022-09-20
环球时报(2022-06-20)2022-06-20
军事文摘(2021年22期)2021-11-26
小读者(2021年2期)2021-03-29
文苑(2020年6期)2020-06-22
文苑(2019年22期)2019-12-07
华人时刊(2019年13期)2019-11-17
文苑(2018年22期)2018-11-19
