
内蒙古农牧交错带小叶杨人工林碳汇效应研究
2016-09-19 06:38:38郭月峰祁伟姚云峰王娟张美丽刘思禹韩兆敏内蒙古农业大学沙漠治理学院内蒙古呼和浩特000内蒙古水利水电勘测设计院内蒙古呼和浩特00020
生态环境学报 2016年6期
郭月峰,祁伟, 2,姚云峰*,王娟,张美丽,刘思禹,韩兆敏. 内蒙古农业大学沙漠治理学院,内蒙古 呼和浩特 000;2. 内蒙古水利水电勘测设计院,内蒙古 呼和浩特 00020
内蒙古农牧交错带小叶杨人工林碳汇效应研究
郭月峰1,祁伟1, 2,姚云峰1*,王娟1,张美丽1,刘思禹1,韩兆敏1
1. 内蒙古农业大学沙漠治理学院,内蒙古 呼和浩特 010011;2. 内蒙古水利水电勘测设计院,内蒙古 呼和浩特 010020
造林工程已经对生态恢复起到重要作用,并将对全球碳素循环起到积极作用。以内蒙古农牧交错带小叶杨(Populus simonii)人工林为研究对象,并以天然草地为对照,分析了退牧还林对生态系统碳储量和碳循环的影响。结果表明,造林活动使植被碳库迅速增加,凋落物碳库及生态系统碳库基本上随林龄的增加而缓慢增加,土壤碳库则随林龄增加出现先降低再增加的趋势;小叶杨林木平均碳汇速率表现为:幼龄林>中龄林>近熟林>成熟林;以成熟林(30 a)为参照,小叶杨林木固碳潜力为4 500 g·m-2,生态系统固碳潜力为5 141.79 g·m-2。研究结果表明,内蒙古农牧交错区退牧还林后种植的小叶杨人工林在长时间尺度上是一个可观的碳汇。
碳储量;小叶杨;内蒙古农牧交错带
引用格式:郭月峰, 祁伟, 姚云峰, 王娟, 张美丽, 刘思禹, 韩兆敏. 内蒙古农牧交错带小叶杨人工林碳汇效应研究[J]. 生态环境学报, 2016, 25(6): 920-926.
GUO Yuefeng, QI Wei, YAO Yunfeng, WANG Juan, ZHANG Meili, LIU Siyu, HAN Zhaomin. Carbon Sink Effect of Afforested Populus simonii Plantations in the Agro-pasture Zigzag Zone in Inner Mongolia [J]. Ecology and Environmental Sciences, 2016,25(6): 920-926.
大规模的造林工程已经对生态恢复起到重要作用,并将对全球碳素循环起到积极作用。森林植被生物量占全球植被生物量的86%,而森林土壤中的碳占全球土壤碳的73%(Guo et al.,2014)。因此,森林生态系统在调节全球碳平衡,减缓大气中CO2等温室气体浓度上升以及维护全球气候等方面具有不可替代的作用(Banaitis et al.,2007;Paul et al.,2002;Guo et al.,2002;聂道平等,1997)。由于全球生态环境恶化日趋严重,这使得国内外研究人员对全球碳循环愈加关注(杨义波,1995;陶波等,2001;周玉荣等,2000;Wu et al.,2014)。因此,研究生态系统固碳功能的意义重大。目前相关学者对退耕、退牧造林工程对全球(Wu et al.,2014)和区域(吴建平等,2013;Enting et al.,2012;Shvidenko et al.,1997;Turner et al.,1995)碳循环的研究较多,而基于生态系统尺度上的碳汇能力的研究相对较少(陈先刚等,2008;魏书精等,2013;张东来等,2008),尤其是对于人工小叶杨林分碳储量的研究报道甚少。在北方农牧交错地带进行退牧还林还草工程,除了恢复植被和水土保持效果外,其植被固碳功能也逐渐引起人们的关注。但基于造林工程起到的固碳能力的研究报道仍然较少,例如,退牧还林还草工程的碳汇评价方法以及固碳潜力等问题报道较少或不足(Zinn et al.,2002)
小叶杨(Populus simonii)为杨柳科杨属乔木,为良好的防风、固沙、保土和绿化树种,是我国主要乡土树种和栽培树种。于20世纪80年代以后开始成为我国北方农牧交错地带的主要造林树种,同时也是内蒙古农牧交错带人工林面积最大的树种。
本文选择内蒙古农牧交错带黄花甸子流域为研究地点,利用时空替代法研究退牧还林后生物量碳储量、凋落物碳储量和土壤碳储量的变化规律,以期为进一步开展造林固碳政策的制定提供科学的指导,为政府参与国际有关全球气候变化的谈判提供良好的案例研究和科学根据。
1 材料和方法
1.1研究区概况
研究区位于内蒙古农牧交错带黄花甸子流域(42°23′~42°27′N,119°40′~119°49′E),面积32 km2,海拔440~906 m。年均降雨量为400~470 mm,年均蒸发量为2290~2400 mm。主要土壤类型为栗钙土,土壤呈碱性或弱碱性。主要植被以人工植被小叶杨(Populus simonii)为主。研究区有比较完整的退牧还林(小叶杨)年限系列。同时,选择未退牧的天然牧草地作为对照点。流域属于半干旱森林草原植被类型区,由于人为破坏自然植被丧失殆尽,逐渐变为半风沙荒草地。为了地区生态恢复,于 20世纪80年代前后开始大规模种植人工林。
1.2小叶杨林林组的划分
将小叶杨林龄划分为幼龄林、中龄林、近熟林及成熟林4个龄组(那妞儿,2012),具体划分见表1。

表1 小叶杨林龄组划分Table1 Age groups division of Populus simonii
1.3样品采集和测试
1.3.1样点选择和样方布设
选择5个不同林龄的小叶杨样区,每个样区均设置3个20 m×20 m的标准样方;同时在每个样方的四角及样方对角线的中央设置草本样方(共 5个),各草本样方的面积均为1 m×1 m;再于每个草本样方内设置3个0.25 m×0.25 m的枯落物样方;同时,在各草本样方内分别对0~10、10~20、20~40、40~60、60~100 cm土层进行土壤样品采集,本文土壤分析数据取0~100 cm的平均值。
天然草地样方(对照样地)设置:选择立地条件与小叶杨林分相一致的未退牧还林的天然草地样区作为对照样地。随机选择5个草地样区,并在每个草地样区随机布设5个1 m×1 m的草本样方,每个草本样方内的植被、枯落物以及土壤的取样方法同小叶杨样方。
1.3.2样品采集和测定
乔木层植被生物量测定:在每个小叶杨样方内,伐倒标准木后称其各器官的生物量,同时取一部分样品带回实验室分析测量其含碳率。
草本生物量测定:将草本样方地上、地下部分全部收获并进行称重,并将采集的样品带回实验室进行烘干计算样品的含水率,进而备测样品的含碳率。
凋落物生物量测定:在每个凋落物样方内收集全部枯枝落叶,取部分样品备测碳含量。
不同乔木、草本植物、枯枝落叶的含碳率测定:植被不同器官及凋落物的含碳率用重铬酸钾硫酸法测定(田大伦,2004)。
土壤样品的测定:土壤容重的测定方法采用环刀取土法,土壤有机碳含量的测定采用重铬酸钾滴定法(中华人民共和国林业行业标准,1999)。
1.4统计分析
1.4.1生物量计算
根据标准地树木的株数,结合上述 1.3.2中小叶杨林分各级生物量的测定方法,经折算进而求出单位面积乔木层生物量以及草本和枯落物层单位面积生物量。
1.4.2植被及凋落物碳储量计算
植被(或凋落物)碳储量由植被生物量(或凋落物)干重乘以测得的含碳率得到:其中,SOC为单位面积植被不同器官或凋落物的碳储量,g·m-2;

M 为单位面积植被不同器官或凋落物的生物量干重,g·m-2;
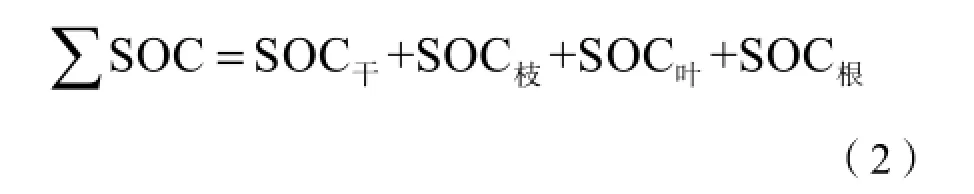
C为单位面积植被不同器官或凋落物的含碳率,g·kg-1。∑SOC为单位面积小叶杨林木碳储量,g·m-2;SOC干为单位面积小叶杨林木树干碳储量,g·m-2;
SOC草为单位面积小叶杨林木枝条碳储量,g·m-2;
SOC叶为单位面积小叶杨林木叶子碳储量,g·m-2;
SOC根为单位面积小叶杨林木根系碳储量,g·m-2;
1.4.3土壤碳储量计算
土壤碳储量计算参考程先富等(2004)的方法,略作修改:
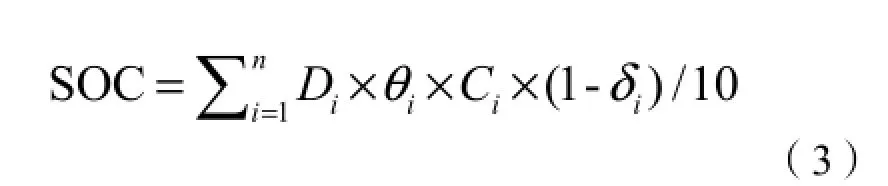
其中,SOC为土壤有机碳密度,g·m-2;n是取土层数,本研究中取5;Di表示取样中不同土层的厚度,cm;θi表示不同土层的土壤容重,g·cm-3;Ci表示不同土层的土壤有机含碳率,g·kg-1;δi为>2 mm砾石含量(体积%)。由于研究区属于黄土丘陵及黄土漫岗,土壤以黄土母质发育而成,砾石含量少;在处理土壤样品时,将全部土样磨碎,因此公式(3)中的δi为0。
1.4.4森林生态系统碳储量计算
森林生态系统总碳储量=植被碳储量+凋落物碳储量+土壤有机碳储量
1.4.5生态系统固碳潜力计算
本研究中造林植被的生态系统固碳潜力是指受自然干扰和基于目前气候状态下,地域由天然草地转化为人工森林后,在后续很长时间内的最大可能固碳量,其计算公式为:

其中,CSP为生态系统固碳潜力,g·m-2;CSr为是参考植被类型的最大碳储量(Keith et al.,2009),g·m-2,本文参考植被类型的最大碳储量为小叶杨林林分碳储量达最大时的林分生态系统碳储量。CS为初始植被类型的碳储量,g·m-2,CS的数值采用的是未种植林木的天然草地生态系统碳储量数值。
2 结果与分析
2.1不同碳库碳储量基线值
以天然草地碳库为研究不同林龄小叶杨固碳潜力的基线值,以小叶杨成熟林碳库为参考值,计算小叶杨的固碳潜力。其中,天然草地各碳库碳储量分别为:植被层为69.69 g·m-2,土壤层为6276.25 g·m-2,凋落物层为44.43 g·m-2,生态系统为6390.37 g·m-2。
2.2小叶杨林分植被层固碳效应
对于小叶杨纯林而言,植被碳储量包括乔木层和草本层的碳储量。小叶杨林分乔木层碳储量变幅为780.00~4500.00 g·m-2,平均为2410.86 g·m-2。从幼龄林—成熟林阶段,林木碳汇过程符合对数曲线函数,拟合方程为y=2288.6lnx-3302.7,R2=0.9984(图1)。林木在幼龄林阶段固碳速率较快,随着年龄的增长,速率在逐渐减缓。
从图1和图2还可看出,幼龄林的固碳速率为203.4 g·m-2·a-1,中龄林的固碳速率为196 g·m-2·a-1,近熟林的固碳速率为 174 g·m-2·a-1,成熟林的固碳速率为 150 g·m-2·a-1。可见对于小叶杨林木平均碳汇速率最大出现在幼龄林阶段,而在林木逐渐进入中龄林直到成熟林的过程,碳汇速率是逐渐降低的,这一结果与高岗(2009)认为小叶杨在5~10 a生长最快的研究结果基本一致。此外,小叶杨植被固碳速率随着林木生长年限的增加而下降,因为随着林木不断生长,树木单株营养面积过小,在没有及时疏株间伐的情况下,满足不了林木正常生长的需求,致使小叶杨树势衰弱;加之林木不断生长,不予以合理修枝,将削弱林木顶端优势或使光合作用积累干物质减少,造成小叶杨林分植被碳储量随林龄增加逐渐降低(郭月峰等,2013)。
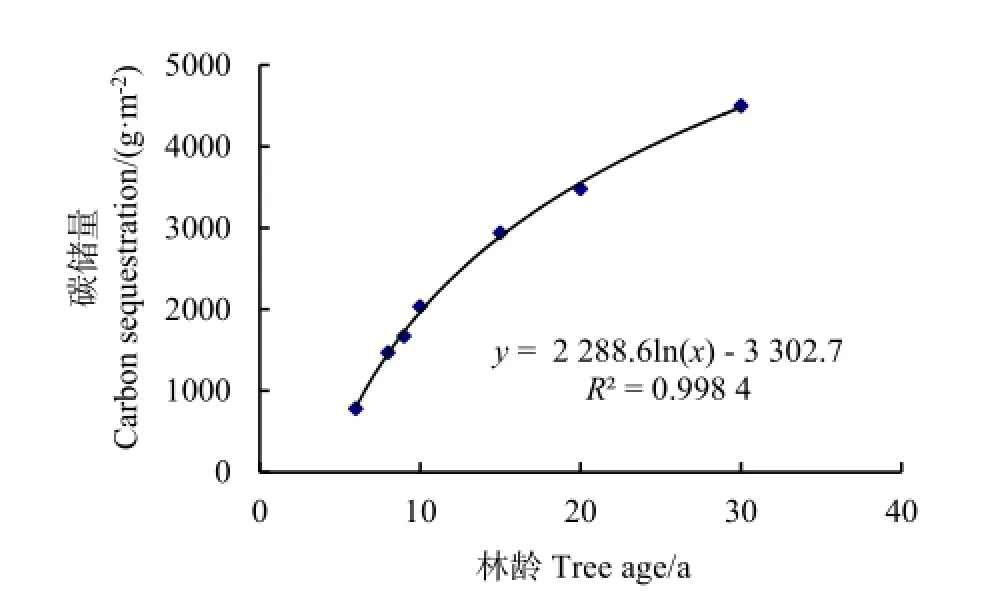
图1 小叶杨林分乔木层固碳过程Fig. 1 Effects of forest carbon sequestration with Populus simonii stand
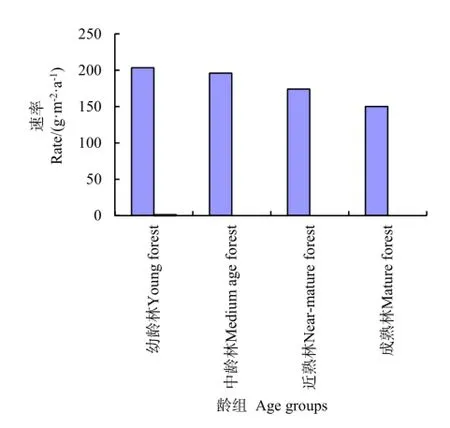
图2 小叶杨林分乔木层平均碳汇速率Fig. 2 The average forest carbon sequestration process with Populus simonii stand
小叶杨林分林下草本层碳储量很少,变幅为124.25~203.25 g·m-2,平均为154.42 g·m-2(图3)。从植被碳储量的组成来看,草本层碳储量较少。随着林木年限增长,草本层碳汇过程基本表现为先增加后减小的趋势,拟合方程为:y=-0.2881x2+ 12.196x+57.994,R2=0.7225。原因是在幼龄林和中龄林阶段,林分郁闭度较低,林下草本不仅可以得到充分的光照,而且还可以较好地利用林冠的遮荫作用,从而为其生长提供了相对有利的条件;随着林木不断生长,林木逐渐达到郁闭,林分的透光性较差,林下形成阴暗环境,导致林下草本的光照资源受到限制,至少没有幼树阶段充足,这使得林下草本出现减少的趋势,故近熟林和成熟林阶段草本层碳储量呈下降的趋势(郭月峰等,2013)。崔国发(1996)在研究中指出,人工林生态系统结构较简单,尤其是纯阔叶单层林,当林分达到一定郁闭度以后,培育树种成为优势树种,使得林下植物难以生存,林下草本也很稀少,形成了所谓的“清膛林”现象。

图3 小叶杨林分草本层固碳过程Fig. 3 Herbaceous layer of carbon sequestration effect with Populus simonii stand
2.3小叶杨林分凋落物层固碳效应
虽然有研究认为植物通过根系脱落输入到土壤中的碳远远超过了地上凋落物的碳输入量,例如,随着植物根系碳分配比例的提高,根系的生理活动进一步增强,根系呼吸、周转和根际分泌等过程的碳流量提高,最终土壤碳含量提高(吴伊波等,2009)。但也有学者认为土壤碳储量的增加主要来源于凋落物的分解,调落物是土壤碳变化的先驱,凋落物的性质和分解速率影响土壤碳库的变化(Liski et al.,2002)。从图4可以看出,随着林龄的增长,小叶杨林地凋落物层碳储量不断增加。其变化趋势可用二次多项式描述,拟合曲线为y=-0.092x2+10.764x-46.763,R2=0.8974。6、8、9、10、15、20及30 a小叶杨林地凋落物层碳储量分别为 32.07、30.28、32.63、49.61、63、168.34及185.54 g·m-2。凋落物主要来源于林木的枯枝落叶和杂草,林木生长为每年凋落物的积累作一定贡献,在造林初期,由于幼树比较小,凋落物的归还量较少,之后,随着林木不断长大,地上生物量的积累使得凋落物回还量增加。此外,因凋落物沉积于林下,受人为扰动相对较轻,每年的分解量有限,所以其碳储量随着林龄增大呈不断增加趋势。凋落物碳储量在整个生态系统中所占比例虽少,但也是一个不容忽略的碳库,减缓凋落物的分解速度对于生态系统的碳截留也起到一定的作用。

图4 小叶杨林分凋落物层碳汇过程Fig. 4 Litter carbon sequestration process with Populus simonii stand
2.4小叶杨林分土壤固碳效应
土壤碳库大小主要由两部分决定:一是植物枯枝落叶的输入量,二是枯枝落叶的分解率。如前面所述,凋落物是土壤碳的先驱,凋落物的性质及分解速率影响着土壤碳库的变化,而且土壤碳储量70%~80%增量来源于凋落物的分解(Liski et al.,2002)。人工造林后,土壤碳库的变化速度较地上生物量碳库的变化速度慢,主要是因为土壤中碳储量变化存在缓冲作用。不同林龄的小叶杨林地土壤碳汇过程见图5。从图5可知,土壤碳储量随林龄增加呈增加趋势,其变化可用二次多项式来描述,拟合方程为y=0.9091x2-10.424x+6162,R2=0.9634。说明不同林龄的林木对土壤碳增汇的影响不同,不同林龄的林木在林中所处的地位各异。
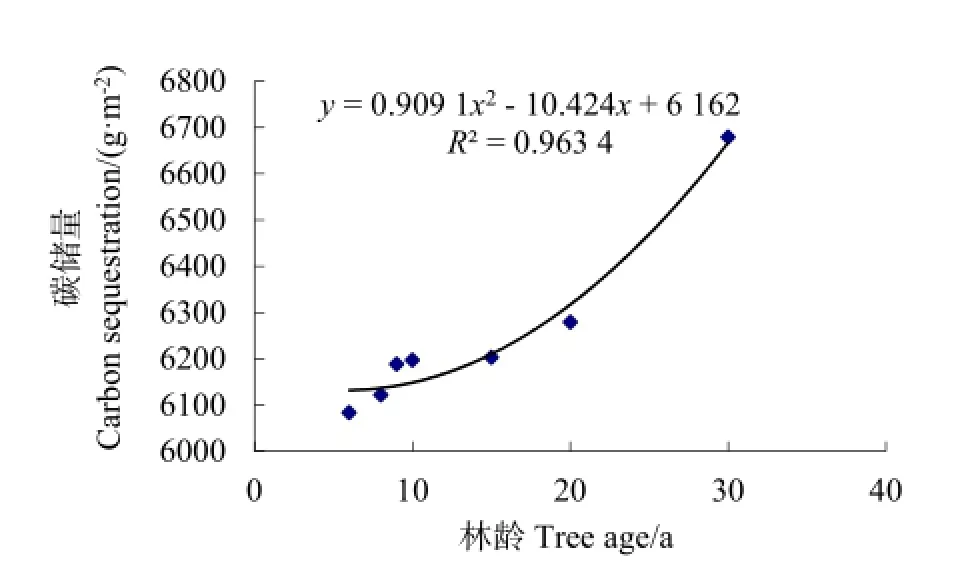
图5 小叶杨林分土壤碳汇过程Fig. 5 Soil carbon sequestration process with Populus simonii stand
土壤有机碳随林龄增加的变幅为 6083.97~6679.21 g·m-2,具体表现为6、8、9、10、15、20 及 30 a小叶杨林地土壤碳储量分别为 6083.97、6122.27、6188.34、6197.41、6203.35、6279.36 及6679.21 g·m-2。土壤碳储量基本呈现出在造林初期,测定值比基线值有所下降,之后随林龄增加而缓慢增加,当小叶杨林达20 a时,土壤碳储量恢复到基线值。当林龄为30 a时,小叶杨林土壤碳储量出现净累积。整个过程可以解释为:当林木种植以后,土壤中碳存变化有个缓冲作用。造林初期,土壤碳储量低于基线值水平,因扰动改善了土壤结构,增加了土壤通气性,提高了土壤微生物活性,加速了土壤有机碳的释放;同时,土壤有机碳积累依赖于凋落物的供给与分解,在造林初期,凋落物量虽有所增加,但凋落物的分解程度不深,从而使土壤有机碳的固定小于释放(崔国发,1996);也有人认为,在造林初期,林木吸收了土壤中的速效养分,在养分归还土壤的过程中,有相当长时间处于难利用状态,营养元素生物循环受到了阻滞(崔国发,1996);小叶杨林木在幼龄林和中龄林阶段生长较快,大量消耗土壤养分。之后随林龄增大,植被生长与土壤养分的耦合效应使土壤条件得到改善,土壤质量提高,使得土壤碳储量开始增加,并最终出现净累积现象。
2.5小叶杨林分生态系统固碳效应
不同林龄的小叶杨系统碳汇过程及平均碳汇速率,见图6和图7。从图6可知,系统碳储量随林龄增加呈增加趋势,其变化趋势可用对数式来描述,拟合方程为y=2662.7lnx+2240.9,R2=0.9901。系统碳储量在幼龄林阶段增长较快,之后变化缓慢。系统平均碳汇速率与林龄的关系呈负相关关系,拟合方程为y=4250.4x-0.709 2,R2=0.9997(图7)。小叶杨林地系统碳汇过程具体表现为6、8、9、10、15、20及 30 a小叶杨林的系统碳储量分别为7139.67、7833.04、8034.49、8346.76、9290.22、10042.96及 11532.16 g·m-2;固碳速率分别为1189.95、979.13 g·m-2、892.72、834.78、619.35、502.15及384.41 g·m-2·a-1。
2.6小叶杨林分生态系统固碳潜力
通过公式(4)计算可知,在研究区种植小叶杨林之后,其幼龄林、中龄林、近熟林及成熟林的系统固碳潜力分别为 1956.39、2899.85、3652.59 及5141.79 g·m-2(图8);30 a内的生态系统固碳潜力为5141.79 g·m-2,其中植被碳库、土壤碳库及凋落物碳库的固碳潜力分别为 4597.72、402.96及141.11 g·m-2(图9)。研究表明,在研究区种植小叶杨林后,生态系统在长时间尺度上是一个可观的碳汇,尤其以植被碳库增汇最明显。
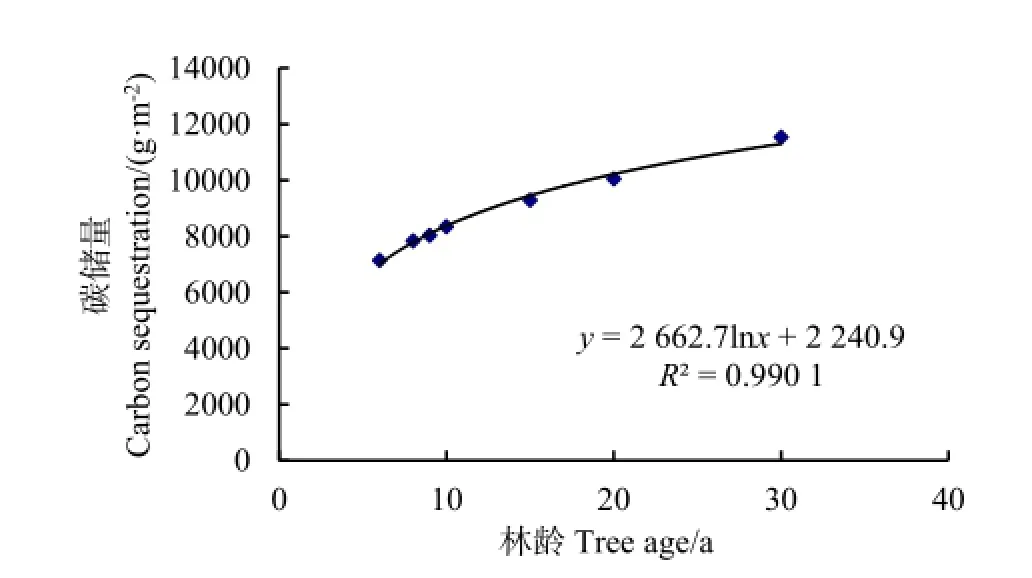
图6 小叶杨林分生态系统碳汇过程Fig. 6 Ecosystem carbon sequestration process with Populus simonii stand
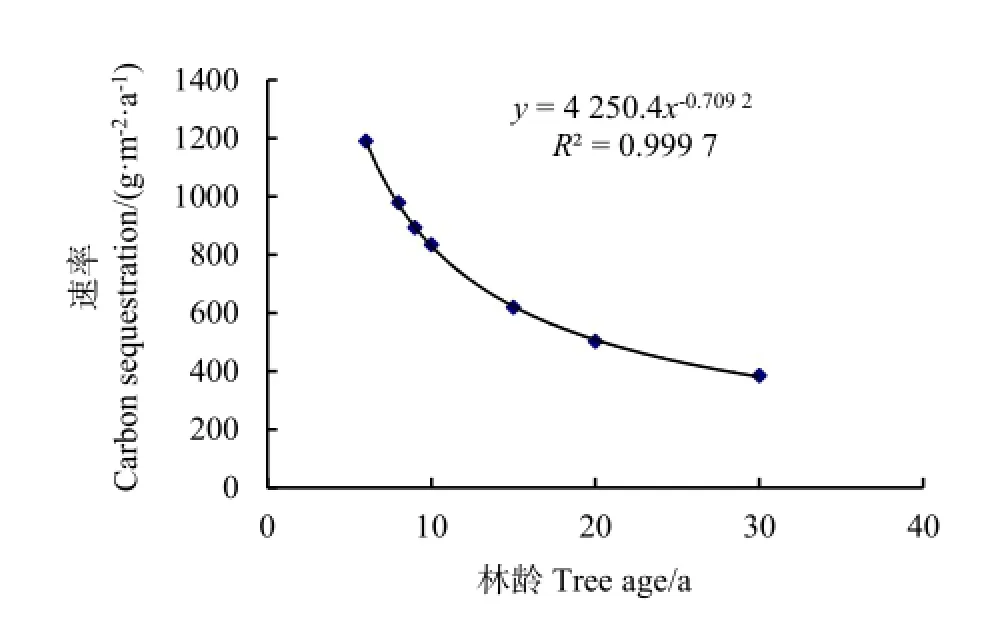
图7 小叶杨林分生态系统平均碳汇速率Fig. 7 Ecosystem carbon sink rate with Populus simonii stand

图8 小叶杨林分不同龄组生态系统固碳潜力Fig. 8 Different age groups forest carbon sequestration potential with Populus simonii stand
3 讨论
造林后生态系统碳储量及各组成碳库碳储量随林龄的变化与已有概念理论和研究结果相似(Goulden et al.,2011),植被碳储量基本呈现逐年增加,而土壤碳库基本呈现先降低后增加的趋势。在没有人为干扰的情况下,生态系统碳储量增加量主要以植被增加的碳储量为主。
从不同林龄的植被固碳速率来看,小叶杨林在22 a后由近熟林转为成熟林(张连翔等,2001),应该予以必要的抚育管理,如间伐或全部皆伐,清除原有植株后重新栽植适宜于当地生长的碳汇林树种或重新栽植原种。或者改纯林为混交林,使林木更好利用林地资源,向健康的方向发展。因此,在我国人工林面积已经非常大的情况下,除造林之外,还应该对已有人工林加强管理,提高生态系统固碳增汇效果。
从天然草地到森林的土地利用方式的改变意味着碳循环周期的改变。改变后的生态系统,会产生更多的生物量。生物量碳作为生态系统中积累的植物有机物总量,是整个生态系统运行的能量基础和营养物质来源,因此分析不同生态系统类型下的植被碳储量具有重要意义。退牧还林后,生物量碳储量明显增加。
土壤碳库变化的速度比在生物量中慢得多,因为土壤中碳储存的变化有个缓冲作用。土壤碳含量主要由两部分决定:一是植物枯枝落叶的输入量,二是枯枝落叶的分解率。当天然草地被转化为森林的时候,天然草地生态系统的碳输入和输出平衡被打破,造林形成新的生态系统的循环机制还不健全,处于一种无序状态。人工林的林木吸收了土壤中的速效养分,在养分归还土壤的过程中,有相当长时间处于难利用状态,营养元素生物循环受到了阻滞(崔国发,1996)。在退牧还林初期,由于幼树比较小,凋落物的归还就较少。因此,退牧还林后土壤碳密度降低,随着地上生物量的积累和凋落物回归的增加,人工林生态系统逐渐形成新的循环机制。在一个稳定的森林生态系统中,根据森林里凋落物的输入和分解率,土壤碳积累将会达到一个新的动态平衡,这时年度变化就不明显。本文研究发现从草地到人工林的土地利用变化过程中,小叶杨林土壤有机碳储量分别比未退牧之前的草地减少8.2%和8.5%。与Guo et al.(2002)的研究有一定的相似性。

图9 小叶杨林分不同碳库固碳潜力Fig. 9 Carbon sequestration potential of different libraries with Populus simonii stand
4 结论
退牧还林后生态系统的植被碳储量、凋落物碳储量和土壤碳储量都随退牧还林年限延长而发生变化。造林活动使植被碳库迅速增加,凋落物碳库及系统碳库基本上随林龄的增加而缓慢增加,土壤碳库则随林龄增加出现先降低再增加的趋势;小叶杨林木平均碳汇速率为:幼龄林>中龄林>近熟林>成熟林;以成熟林(30 a)为参照,小叶杨林木固碳潜力为 4500 g·m-2,系统固碳潜力为 5141.79 g·m-2,其中植被碳库、土壤碳库及凋落物碳库的固碳潜力分别为4430.31,402.96及141.11 g·m-2。以上结果表明,燕山北部低山丘陵区退牧还林后,人工林生态系统在长时间尺度上是一个可观的碳汇。
BANAITIS M R, LANGLEY-TURNBAUGH S J, ABOUEISSA A A. 2007. Variations of soil organic carbon in three urban parks: A Maine Case Study [J]. International Journal of Applied Environmental Sciences, 2: 119-128.
ENTING G, RAYNERL P J, CIAIS P. 2012. Carbon Cycle Uncertainty in regional Carbon Cycle Assessment and Processes (RECCAP)[J]. Biogeosciences, (9): 2889-2904.
GOULDEN M L, MCMILLAN A M S, WINSTON G C, et al. 2011. Patterns of NPP, GPP, respiration, and NEP during boreal forest succession [J]. Global Change Biology, 17(2): 855-871.
GUO L B, GIFFORD R M. 2002. Soil carbon stocks and land use change: a meta analysis [J]. Global Change Biology, (8): 345-360.
GUO Y F, YAO Y F, QIN F C. 2014. Effects of land use changes on soil organic carbon and soil microbial biomass carbon in low hills of north Yanshan Mountains [J]. Range Management and Agroforestry, 35(1): 15-21.
KEITH H,MACKEY B G, LINDENMAYER D B. 2009. Re-evaluation of forest biomass carbon stocks and lessons from the world's most carbon-dense forests [J]. Proceedings of the National Academy of Sciences of the United States of America, 106(28): 11635-11640.
LISKI J, PERRUCHOUD D, KARJALAINEN T. 2002. Increasing carbon stocks in the forest soils of western Europe [J]. Forest Ecology and Management, 169(1-2): 159-175.
PAUL K I, POLGLASE P J, NYAKUENGAMA J G, et al. 2002. Change in soil carbon following afforestation [J]. Forest Ecology and Management, 168(1-3): 241-257.
SHVIDENKO A, NILSSON S, ROSHKOV V. 1997. Possibilities for increased carbon sequestration through the implementation of rational forest management in Russia [J]. Water, Air, Soil Pollution, 94(1): 137-162.
TURNER D P, KOERPER G J, HARMON M E, et al. 1995. Carbon sequestration by forests of the United States-current status and projections to the year 2040 [J]. Tellus Series B-chemical & Physical Meteorology, 47(1-2): 232-239.
WU J, LIU Z, HUANG G, et al. 2014. Response of soil respiration and ecosystem carbon budget to vegetation removal in Eucalyptus plantations with contrasting ages [J]. Scientific Reports, 4: 6262.
ZINN Y L, RESCK D V S, SILVA J E D. 2002. Soil organic carbon as affected by afforestation with Eucalyptus and Pinus in the Cerrado region of Brazil [J]. Forest Ecology and Management, 166(1-3): 285-294.
陈先刚, 张一平, 詹卉. 2008. 云南退耕还林工程林木生物质碳汇潜力[J]. 林业科学, 44(5): 24-30.
程先富, 史学正, 于东升, 等. 2004. 兴国县森林土壤有机碳库及其与环境因子的关系[J]. 地理研究, 23(2): 211-217.
崔国发. 1996. 人工林地力衰退机理及其防止对策[J]. 世界林业研究,(5): 61-69.
高岗. 2009. 以水源涵养为目标的底功能人工林更新技术研究[D]. 呼和浩特: 内蒙古农业大学.
郭月峰, 姚云峰, 秦富仓, 等. 2013. 燕山典型流域两种造林树种生态系统碳储量及固碳潜力研究[J]. 生态环境学报, 22(10): 1665-1670.
那妞儿. 2012. 林龄的划分[EB/OL]. 2014-03-17. http://blog.sina.com.cn/ s/blog_8733be4b0101lmxf.html.
聂道平, 徐德应, 王兵. 1997. 全球碳循环与森林关系的研究——问题与进展[J]. 世界林业研究, 10(5): 34-40.
陶波, 葛全胜, 李克让. 2001. 陆地生态系统碳循环研究进展[J]. 地理研究, 20(5): 564-575.
田大伦. 2004. 杉木林生态系统定位研究方法[M]. 北京: 科学出版社: 81-87.
魏书精, 罗碧珍, 孙龙, 等. 2013. 森林生态系统土壤呼吸时空异质性及影响因子研究进展[J]. 生态环境学报, 22(4): 689-704.
吴建平, 刘占锋. 2013. 森林净生态系统生产力及其生物影响因子研究进展[J]. 生态环境学报, 22(3): 535-540.
吴伊波, 崔骁勇. 2009. 草地植物根系碳储量和碳流转对 CO2浓度升高的响应[J]. 生态学报, 29(1): 378-388.
杨义波. 1995. 长白落叶松(Larix olgensis)幼树生物量的初步研究[J]. 吉林林学院学报, 11(2): 22-25.
张东来, 毛子军, 朱胜英, 等. 2008. 黑龙江省帽儿山林区6种主要林分类型凋落物研究[J]. 植物研究, 28(1): 104-108.
张连翔, 梅秀艳, 姜镇荣. 2001. 小叶杨生长规律的研究[J]. 防护林科技,47(2): 10-12.
中华人民共和国林业行业标准. 1999. 森林土壤分析方法[M]. 中国标准出版社: LY/T 1210-1275.
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522.
Carbon Sink Effect of Afforested Populus simonii Plantations in the Agro-pasture Zigzag Zone in Inner Mongolia
GUO Yuefeng1, QI Wei1, 2, YAO Yunfeng1*, WANG Juan1, ZHANG Meili1, LIU Siyu1, HAN Zhaomin1
1. Desert Science and Engineering College, Inner Mongolia Agricultural University, Hohhot, China;
2. Department of Forestry, Inner Mongolia Agricultural University, Hohhot, China
Afforestation projects play an important role in ecosystem restoration and will contribute to global carbon cycles. The study target was the Populus simonii man-made forests in the agro-pasture zigzag zone of Inner Mongolia, with natural grasslands as a control. We summarized and analyzed how the "conversion from grazing land into forest" affected the ecosystem carbon stocks and carbon cycles.The afforestation activities led to rapid increase of vegetation carbon stocks, since with the prolonging of forest age,the litter carbon stocks and ecosystem carbon stocks slowly increased, but soil carbon stocks first dropped and then rose. The average carbon sink rates change with forest age as follows: young forest > medium age forest > near-mature forest > mature forest.With mature forest (30 a) as reference, the P. simonii forest carbon sequestration potential (CSP) is 4 500 g·m-2, while the P. simonii ecosystem CSP is 5 141.79 g·m-2. These results indicate that after conversion from grazing land into forest in the study area, the P. simonii ecosystem is a considerable carbon sink in the long term.
carbon stock; Populus simonii; agro-pasture zigzag zone in Inner Mongolia
10.16258/j.cnki.1674-5906.2016.06.002
X173; Q148
A
1674-5906(2016)06-0920-07
国家自然科学基金项目(31500584);内蒙古应用研究与开发计划项目(20110732);高等学校科学研究项目(NJZZ16055);内蒙古自然科学基金项目(2016MS0407)
郭月峰(1982年生),女,讲师,博士,主要从事水土保持与荒漠化防治研究。E-mail: guoyuefeng0525@126.com
姚云峰,教授,博士,主要从事水土保持研究。E-mail: 18904718855@163.com
2016-01-08
猜你喜欢
农民致富之友(2023年30期)2023-11-21 07:09:48
防护林科技(2020年10期)2020-12-22 11:46:42
绿色科技(2020年20期)2020-11-20 01:56:34
农民致富之友(2020年15期)2020-05-25 09:05:41
福建农林大学学报(自然科学版)(2020年1期)2020-01-10 06:27:00
河南农业大学学报(2017年6期)2018-01-10 06:03:56
江苏农业科学(2017年6期)2017-05-11 14:35:37
林业勘查设计(2017年1期)2017-03-11 16:49:57
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
