
马尾松改造林分多样性及种子雨与土壤种子库
2016-09-15 15:46:17王丽艳刘光正岳军伟金洪平
福建林业科技 2016年1期
王丽艳,刘光正,岳军伟,龙 蔚,金洪平,杨 桦
(江西省林业科学院,江西 南昌 330013)
马尾松改造林分多样性及种子雨与土壤种子库
王丽艳,刘光正,岳军伟,龙 蔚,金洪平,杨 桦
(江西省林业科学院,江西 南昌 330013)
以江西省万年县中北部低效马尾松改造后25年生左右的林分为研究对象,采用样地调查法调查林区植被类型,利用种子收集器收集种子,取样分析改造后马尾松林下木荷、杉木种子雨和土壤种子库特征及垂直分布格局。结果显示:改造后的马尾松林中,林下乔木以木荷、杉木为主。从垂直分布来看,木荷、杉木土壤种子库总量分布明显呈现从上到下逐渐减少的趋势。木荷枯落物层种子库总量均超过取样收集的种子数量的70%,0~5 cm土层中单位面积种子库总量在4~11粒·m-2,木荷无生活力种子的数量和比率都较高。杉木种子库总量平均值分别为49.6、24.8、23粒·m-2。杉木土壤种子库中枯落物层、0~5 cm、5~10 cm土层有生活力种子密度均值分别为8.77、1.23、3.50粒·m-2,占种子库总量均值的53.02%、14.82%、45.05%。
低效马尾松林;改造;植物多样性;种子雨;土壤种子库
种子是植物有性繁殖的重要器官,“雨”表示数量集中,种子雨是对植物繁殖体在特定的时间和空间从母株上散落的种子量[1]。土壤种子库种子储备作为潜在繁殖转化为植物个体的重要阶段,对种群的更新有着重要影响[2-3]。土壤种子库是存在于枯落物层和土壤中的全部种子之和[4]。国内外对亚热带常绿阔叶林种群的种子雨、种子库动态的研究主要集中在建群种,如:石栎(Lithocarpusglaber(Thunb.) Nakai)[5]、青冈(Cyclobalanopsisglauca(Thunb.) Oerst.)[2]、扁刺栲(Castanopsisalatyacantha)[6]、苦枥(Fraxinusinsularis)[7]等,而对于阔叶化改造的马尾松林种子雨及种子库的研究较少。本研究以江西省石镇阔叶化改造后马尾松林作为研究对象[8],采用种子收集器,通过野外大量取样观测,探讨林区内乔木种子散落特征和土壤种子库的分布格局,研究成果对阔叶林类型中种子雨与土壤种子库动态的认识将为制订适合江西省的森林修复措施和促进鄱阳湖生态经济区建设提供理论参考。
1 研究区概况
研究区位于万年县中北部的石镇镇大黄村,于1987年对低效马尾松林(林龄在35 a左右,平均胸径10.82 cm,树高9.51 m)进行阔叶化改造,主要栽植乡土树种木荷、枫香等。木荷是目前表现最好的树种,枫香也是林分中数量相对较多的阔叶树种。经过25 a的演替变化,林分中原有的幼龄杉木也逐渐成长为局部的优势树种,与马尾松形成混交林。该研究区是典型的低丘岗地,为亚热带季风气候,年均日照时间1803.5 h,年平均温度17.4 ℃,年均相对湿度为82%,年均无霜期达259 d,年均降水量为1808.0 mm,年降水量最大、最小值分别为2879.7 mm和1230.7 mm。8—10月雨量最少。土壤主要是第四纪红色粘土母质形成的红粘性红壤[9]。该区主要木本植物种类有马尾松(PinusmassonianaLamb.)、杉木(Cunninghamialanceolata)、苦槠(Castanopsissclerophylla)、石栎、木荷(Schimasuperba)、菝葜(Smilaxchina)、黄瑞木(Adinandramillettii)、乌饭(Vacciniumbracteatum)、黄栀子(Gardeniajasminoides)、胡枝子(Lespedezabicolor)、淡竹叶(Lophatherumgracile)、铁芒萁(Dicranopterislinearis)等。
2 研究方法
2.1 样地布设
2011年7月,在试验林中设置面积为400 m2的乔木样地8个。样地设置方法为:先在林中拉出2条平行的条带,每个条带长约100 m,宽约20 m,在每个条带内以间距5 m左右的距离布设4个20 m×20 m的正方形样地,8个样地平行分布于2个条带内。在样地内对直径≥4 cm的乔木种进行每木检尺,同时调查乔木层郁闭度、种类及数量。在乔木样方内按对角线法设5 m×5 m的灌木样方3个,分别记录每个方格内灌木、草本的种类、数量、高度和盖度[10]。根据植物群落的地上植被数据计算物种丰富度、多样性和均匀度。在设定样地内按S型随机挖取5个土壤剖面,分别选取土壤剖面0~20、20~40、40~60 cm土样待测。
2.2 种子雨观测
种子雨收集采用种子收集器进行。用4块长×宽×厚约100 cm×10 cm×1 cm的木板做成木框,然后将网眼为2 mm×2 mm的尼龙网钉于框底,制成网口面积为1 m×1 m种子收集器。在每个样地内设置3组种子收集器,每组种子收集框包括1个离地种子收集器(用4根木棍做支架,选择比较平坦的地方,将种子收集器用铁丝固定于网底高出地面1.0 m的位置)和1个地面种子收集器(除去离地种子收集器旁边的杂草和小灌木,将种子收集器平放于地面上),8个样地设置24个离地1 m高的种子收集器,同时设置24个地面种子收集器收集落到地面的种子。
种子收集与统计:2012年8月下旬,根据2011年的物候观察和结实情况,开始收集掉落的种子,每15 d左右收集1次,直到没有种子降落为止。每次收集的种子分别装袋,带回实验室后按完好种子(子叶活力良好)、受虫害种子、未成熟种子、败育种子4个标准类型统计。当种子成熟高峰时,离地种子收集器中种子和材料观测的频率较高,而在其它时间较少。每次记录尽可能保持样地的自然状态。最后,将收集的所有成熟完好的种子用氯化三苯四氮哩(TTC)染色法进行种子活力的鉴定。
2.3 土壤种子库测定
于2011年11月27日(第1次取样)、2012年 11月28日(第3次取样)种子散落即将结束时及种子萌发结束的2012年8月16日(第2次取样),分3次从每个地面种子收集器附近选取1个10 cm×10 cm小样方,按0~5、5~10 cm采取含有枯落物层的土壤样品。因为枯落物层也可能含有大量种子,取样时应含有枯落物层,采样总数为24×3=72个。采样带回后,枯落物层中的种子直接筛选分拣,0~5、5~10 cm土壤样品则转入孔径为2mm的尼龙网袋中,用水冲洗掉泥土,将余下的枯落物、根系、种子等杂物,摊开晾晒后,分拣出其中的种子,按不同土层记录完好种子、破损霉变种子、空粒种子的数量,并将完好的种子计数后做生活力测定。
2.4 数据计算
以每个样方为单位,分别计算群落各层物种重要值,乔木层物种重要值=[相对优势度(%)+相对密度(%)+相对高度(%)]/3,灌木、草本层物种重要值=[相对频度(%)+相对盖度(%)+相对高度(%)]/3[11]。
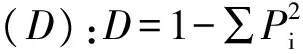
通过Excel 2007和统计软件DPS 7.5进行数据计算分析。
3 结果与分析
3.1 研究区植被类型
通过对8块样地的每木检尺,马尾松低效改造林林分结构特征见表1。调查样地数量占比大的树种为马尾松、杉木和木荷。

表1 改造后马尾松林分结构特征
对群落结构有明显控制作用的植物中通常是个体数量多、体积较大的优势度较大种,而重要值是用来表示某个体在群落中的地位和作用的综合数量指标[12-13]。根据各样地乔木层物种的重要值进行聚类分析,将石镇镇大黄村马尾松改造分为2种群落类型,结果见图1。Ⅰ类:马尾松+木荷林,包括1号、2号、5号、6号样地,林下优势灌木种为乌饭、木荷、白栎(QuercusfabriHance)、黄瑞木、乌药(Linderaaggregata(Sims) Kosterm.)、杜鹃(RhododendronsimsiiPlanch.),草本主要为铁芒萁;Ⅱ类:马尾松+杉木林,包括3号、4号、7号、8号样地,林下优势灌木种乌饭、黄瑞木、乌药、杜鹃等,草本主要为铁芒萁等。
3.2 林地土壤性质
改造后的马尾松林分土壤理化性质见表2。土壤pH值均在4.21~4.56之间,且均呈现随土壤深度增加pH值增大的趋势,2种群落类型pH值大小为马尾松+杉木群落>马尾松+木荷群落。马尾松+杉木群落的土壤有机质、速效N平均含量分别为10.40 g·kg-1、54.20 mg·kg-1,均大于马尾松+木荷群落的土壤有机质、速效N平均含量(6.98 g·kg-1、44.45 mg·kg-1),这可能与马尾松+杉木群落林分透光性较好,林下小乔木和灌、草种类较多有关。同一群落类型的土壤有机质、速效N含量同样都表现出随土层深度增加而减小的趋势。马尾松+木荷群落的土壤速效P、速效K平均含量分别为0.63 mg·kg-1、32.09 mg·kg-1,均小于马尾松+杉木群落的土壤速效P、速效K平均含量(0.97 mg·kg-1、39.85 mg·kg-1)。同一群落内,土壤速效P、速效K的含量都随土层深度的增加表现出先减后增的变化趋势。

表2 改造后的马尾松林分土壤养分
3.3 改造后马尾松林分植物多样性指数
3.3.1 乔木层植物多样性 马尾松改造林植物多样性结果见表3。改造后低效马尾松形成的2种林分类型乔木层的物种丰富度大小为马尾松+杉木林(7.75)>马尾松+木荷(5.00),各样地乔木树种较少,丰富度最高的7号样地也只有11个树种,丰富度最低的1号样地和2号样地只有3个树种。马尾松+木荷林分类型样地(1号和2号)的优势树种为原有马尾松及阔叶化改造后保存下来的木荷,木荷冠幅宽大,枝叶浓密,对其它树种的更新生长产生了抑制作用,这可能是其物种丰富度低的原因。马尾松+杉木林分主要树种是马尾松和杉木,冠幅稀疏,林下光照相对充足,故其物种丰富度高于马尾松+木荷林分。2种林分类型乔木层的D、DMC、H′和H指数大小顺序表现为马尾松+杉木>马尾松+木荷,说明马尾松+杉木林分乔木层物种多样性高(表3)。均匀度指数(E)则相反,马尾松+木荷(0.63)>马尾松+杉木(0.62)。
3.3.2 灌木层物种多样性指数 8块样地的物种丰富度指数(S)相差不大,即2种林分灌木层物种丰富度相差不大,马尾松+木荷林分物种丰富度为9,马尾松+杉木林分物种丰富度为9.25。各样地灌木层植物多样性指数D、DMC和H′大小顺序大体相同,均为8号>4号>5号>6号>3号>7号>2号>1号样地。各样地的Brillouin指数(H)和均匀度指数(E)大小顺序不同,指数H由大到小顺序为4号>5号>6号>3号>2号>7号>1号>8号样地,指数E大小顺序为8号>7号>6号>3号>4号>5号>2号>1号样地。灌木层物种多样性受乔木层树种影响较大,乔木层中物种多样性最小的1号、2号样地,由于树种较少,且受木荷的郁闭抑制作用,其林下灌木层生物多样性也最低。而乔木层生物多样性最高的8号样地为马尾松+杉木林分,其林下灌木层物种多样性也最高。马尾松+杉木林分灌木多样性高于马尾松+木荷林分。改造后马尾松林分灌木层的各多样性指标均大于乔木层相应的多样性指标,表明在改造后的马尾松林中,灌木层的物种多样性高于乔木层物种多样性。
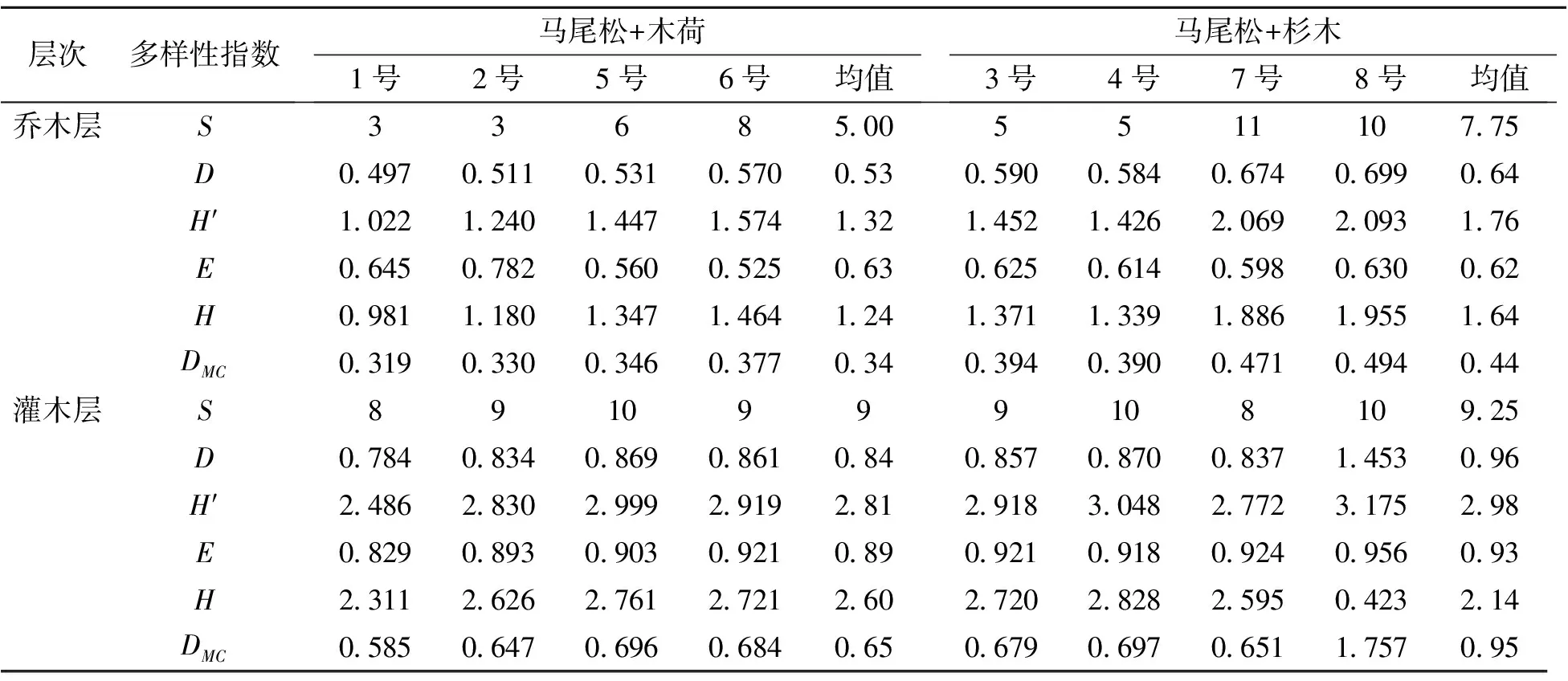
表3 改造后马尾松林分植物多样性指数
3.4 种子雨特征
在整个种子雨样品收集研究过程中,马尾松的种子很少,几乎没有发现,故本文没有对马尾松的种子雨和种子库进行分析,仅针对低效马尾松阔叶化改造后木荷、枫香和杉木的种子雨过程开展分析。
3.4.1 改造后马尾松林分主要树种种子雨基本特征
1)木荷种子雨基本特征。木荷种子大约在8月底—9月初开始逐渐成熟散落,到12月上旬结束,种子下落期持续3个多月(图2)。木荷种子从8月底开始散落,之后种子下落数量持续增加,到10月25日前后达到峰值43.67粒·m-2,此后半个月种子雨散落量略有下降,为41.67粒·m-2,11月上旬之后,种子散落量急剧下降,至12月上旬末种子雨过程基本结束。木荷种子雨集中下落时间也为10月9日—11月8日,集中散落期落下的种子数量为下落种子总量的63.58%。
木荷种子同样分为有生活力种子、霉变破损种子和败育种子。整个种子雨过程有生活力种子总数占种子雨总量的69.42%,霉变破损种子和败育种子数量分别占种子总数的41.82%和21.92%(图2)。
2)杉木种子雨基本特征。杉木种子雨动态过程见图3。杉木种子雨从9月上旬开始,之后种子下落数量快速增加,至10月9日达87.35粒·m-2,此后下落数量稳定在一个较高的水平,到11月8日达到峰值97.77粒·m-2,然后下落数量急剧下降,到11月23日前后种子雨过程基本结束。10月上旬—11月上旬为种子集中降落期,杉木种子散落的数量占落种总数的84.12%。
在杉木种子中,败育种子、破损霉变种子和虫害种子分别占25.85%、5.83%和34.43%,而有生活力种子数量少,仅为33.90%,这与何福基[14]的研究结果一致,何福基认为由于受遗传因素、授粉方式及气象因素的影响,杉木有生活力种子的占比较低。
3)枫香种子雨基本特征。枫香种子从9月底开始下落(图4),之后散落量快速攀升至23.21粒·m-2,到10月下旬,种子散落量达到峰值30.64粒·m-2,之后种子降落量缓慢下降,至11月下旬,种子降落量低至20.96粒·m-2。此后种子散落量急剧下降,至12月上旬基本结束,整个种子雨过程持续60 d左右。10月上旬—11月下旬为枫香种子集中散落期,期间散落的种子数占散落的种子总量的97.82%。
在枫香种子中,败育种子、破损霉变种子和虫害种子分别占35.07%、10.85%和30.73%,而有生活力种子数量仅占24.24%。
3.4.2 改造后马尾松林土壤种子库特征 根据样地调查资料,改造后马尾松林中,林下乔木幼苗和小乔木以木荷、杉木为主,只有有极少量的枫香,故土壤种子库部分只对木荷和杉木进行研究。
1)木荷土壤种子库特征。3次取样木荷种子库总量平均值分别为40.7、19、19.2粒·m-2(表4),后2次取样种子库总量无显著差异,但均与第1次取样时的种子数量有显著差异。2011年11月27日与2012年11月28日2次种子雨即将结束之际收集的种子库总量可能由于大小年问题差异显著,另外,与常绿阔叶林中的木荷相比,马尾松林中种子库总量偏少,可能是其林下土壤瘠薄造成的。从第1、2次取样可以看出,单位面积有生活力种子的数量随时间的变化逐渐减少,破损霉变种子数量随时间变化逐渐增加,败育种子数量随时间变化而减少,这符合一般常理。3次取样收集到的有生活力种子数量差异显著,败育种子数量也存在显著差异,破损霉变种子数量第1次与第3次之间无显著差异,但均与第2次取样数量差异显著。

表4 木荷土壤种子库变化动态
*:同列不同小写字母为差异显著。下同。
将各次取样的各土层木荷种子按有生活力种子、无生活力种子汇总,并进行方差分析,结果见表5。从垂直分布来看,3次取样中,木荷土壤种子库总量分布明显呈现从上到下逐渐减少的趋势,和其它树种一致。第2次样品收集的枯落物层中种子单位面积数量约为该次取样时种子库总量的70%,另外2次取样枯落物层种子库总量均超过本次取样收集的种子数量的70%。第2次取样与第3次取样之间种子密度差异不显著,但均与第1次取样之间差异显著。0~5 cm土层中单位面积种子库总量在4~11粒·m-2之间,大约占该次取样的比例在20%~30%之间,本层中,后2次取样种子总量差异不显著,但均与第1次取样时数量差异显著。5~10 cm土层,由于所处土层较深,单位面积种子数量极少,不作更多讨论。
枯落物层,有生活力种子密度3次取样间均有显著差异,3次取样有生活力种子数量单位面积平均值分别为14.5、1.9、6.7粒·m-2,0~5 cm土层后2次取样之间有生活力种子密度无显著差异,但均与第1次取样时有显著差异。第1次到第3次,0~5 cm 土层有生活力种子单位面积的平均数量分别为5.4、1.5、1.6粒·m-2。
每次取样时,木荷无生活力种子的数量、比率都较高,在木荷种子萌发结束的8月,取样时枯落物层中无生活力种子比例高达85.7%。即使2次在种子雨过程尚未完全结束的11月份收集的样品中,无生活力种子所占的比例也在50%左右或者更高。

表5 木荷土壤种子库垂直分布格局
2)杉木土壤种子库特征。3次取样杉木种子库总量平均值分别为49.6、24.8、23粒·m-2(表6),后2次取样种子库总量无显著差异,但均与第1次取样时的种子数量有显著差异。2次种子雨即将结束之际(11月)收集的种子库总量之间差异显著。从第1、2次取样可以看出,有生活力种子数量随时间的变化逐渐减少,破损霉变种子数量随时间变化逐渐增加,败育种子数量随时间变化而减少,这符合一般常理。有生活力种子数量3次取样之间存在显著差异,败育种子数量3次取样之间也存在差异显著,第1次与第3次收集的样品,破损霉变种子数量之间无显著差异,但均与第2次收集的样品中破损霉变种子数量差异显著。

表6 杉木土壤种子库变化动态
将各次取样的各土层杉木种子按有生活力种子、无生活力种子汇总,进行方差分析,结果见表7。从垂直分布来看,3次取样中,杉木土壤种子库总量分布同样呈现随枯落物层、土层0~5 cm和5~10 cm 3个层次从上到下逐渐减少的趋势。3次样品收集的枯落物层中种子数量所占比例均接近或超过该次取样时种子库总量的70%。第2次取样与第3次取样之间种子密度差异不显著,但均与第1次取样之间差异显著。0~5 cm土层中单位面积种子库总量在4~13粒·m-2之间,大约占该次取样的20%~30%,本层中后2次取样种子总量差异不显著,但均与第1次取样时数量差异显著。5~10 cm土层,第1次与第3次种子数量差异不显著,第2次取样由于种子下落时间久和所处土层较深的原因,单位面积种子数量极少。
在枯落物层3次取样中,有生活力种子密度有显著差异,3次取样杉木有生活力种子数量分别为19.4粒·m-2(2011年11月27日)、2.1粒·m-2(2012年8月16日)和8.5粒·m-2(2012年11月28日),占枯落物层种子库总量的比例分别是54.1%、12%和48.8%;0~5 cm 土层后2次取样有生活力种子数量无显著差异,它们与第1次取样的生活力种子数量有显著差异。每次取样时,杉木由于自身遗传及环境条件影响,无生活力种子的数量、比率都较高,在杉木种子萌发结束的8月份,由于霉变和部分有生活力种子萌发成幼苗,枯落物层、土层0~5 cm、5~10 cm 3个层合计的无生活力种子比例高达85.08%。即使2次在种子雨过程尚未完全结束的11月份收集的样品中,3个取样层合计无生活力种子所占的比例分别为46.77%、54.35%。3次取样中,杉木土壤种子库中枯落物层、土层0~5 cm、5~10 cm 3个层次有生活力种子密度均值分别是8.77、1.23、3.50粒·m-2,占各取样时间种子库总量均值的比例为53.02%、14.82%、45.05%。

表7 杉木土壤种子库垂直分布格局
4 结论
1)江西万年县石镇镇大黄村马尾松改造林群落类型可分为:Ⅰ马尾松+木荷群落,Ⅱ马尾松+杉木群落2种类型。马尾松+木荷群落林下土壤速效P、K含量高于马尾松+杉木混交林地,而土壤速效N和有机质为马尾松+杉木林地含量高于马尾松+木荷群落。
2)改造后的马尾松林,其灌木层生物多样性受乔木层多样性影响较大,乔木层生物多样性高的样地,其林下灌木层物种多样性也较高。马尾松和杉木混交林林冠稀疏,林下光照相对充足,故其物种丰富度是各样地中最高。
3)木荷种子雨大约在8月底—9月初开始逐渐成熟散落,到12月上旬结束,10月25日前后到达峰值43.67粒·m-2。整个种子雨过程有生活力种子总数占种子雨总量的69.42%。枫香种子从9月底开始下落,整个种子雨过程持续时间为60 d左右,从10月上旬—11月下旬,为枫香种子集中散落期,期间散落的种子数占散落种子总量的97.82%。在枫香种子中,败育种子、破损霉变种子、虫害种子分别占35.07%、10.85%、30.73%,而有生活力种子数量仅占24.24%。杉木种子雨从9月上旬开始,到11月8日达到峰值97.77粒·m-2,到11月23日前后种子雨过程基本结束。10月上旬—11月上旬的种子集中降落期,生活力种子数量少,仅为33.90%。
4)改造后的马尾松林中,林下乔木幼苗和小乔木以木荷、杉木为主。从垂直分布来看,3次取样中,木荷和杉木土壤种子库总量分布明显呈现从上到下逐渐减少的趋势。木荷枯落物层种子库总量均超过取样收集的种子数量的70%,0~5 cm土层中单位面积种子库总量在4~11粒·m-2之间,大约占该次取样的比例在20%~30%之间,每次取样时,木荷无生活力种子的数量、比率都较高。在枯落物层3次取样中,杉木土壤种子库有生活力种子密度均有显著差异。3次取样杉木有生活力种子数量分别为19.4粒·m-2(2011年11月27日)、2.1粒·m-2(2012年8月16日)和8.5粒·m-2(2012年11月28日),占枯落物层种子库总量的比例分别是54.1%、12%和48.8%。3次取样杉木种子库总量分别为49.6、24.8、23粒·m-2。
[1]于顺利,郎南军,彭明俊,等.种子雨研究进展[J].生态学杂志,2007,26(10):1646-1652.
[2]黄雍荣,马祥庆,庄凯,等.福建闽清福建青冈天然林种子雨和种子库[J].热带亚热带植物学报,2010,18(1):68-74.
[3]Caballero J.M.,Olano J.L.,Escudero A..Seed bank structure along a semi-arid gypsum gradient in Central Spain.Journal of Arid Environments[J].2003(55):287-299.
[4]吴敏,张文辉,周建云,等.秦岭北坡不同生境栓皮栎种子雨和土壤种子库动态[J].应用生态学报,2011,22(11):2807-2814.
[5]彭军,李旭光,付永川,等.重庆四面山常绿阔叶林建群种种子雨、种子库研究[J].应用生态学报,2000,11(1):22-24.
[6]浙江天童受损常绿阔叶林烤树与木荷的种子雨、种子库和幼苗库[D].上海:华东师范大学,2005.
[7]杜彦君,马克平.浙江古田山自然保护区常绿阔叶林种子雨的时空变异[J].植物生态学报,2012,36(8):717-728.
[8]杨桦,刘光正,岳军伟.红壤低丘低效马尾松林林相改造树种选择研究[J].江西林业科技,2009,37(12):1-6.
[9]岳军伟,朱恒,龙蔚.红壤低丘马尾松低效林主要植物群落物种多样性[J].林业科技开发,2010,24(5):53-56.
[10]杨玉海,陈亚宁,李卫红.新疆塔里木河下游土壤特性及其对物种多样性的影响[J].生态学报,2008,28(2):602-611.
[11]刘光正,林小凡,岳军伟.江西德兴天然次生林林下幼苗更新特征[J].江西林业科技,2014,42(6):10-14.
[12]薛建辉.森林生态学[M].北京:中国林业出版社,2006.
[13]刘光正,岳军伟,林小凡,等.德兴不同类型常绿阔叶林土壤养分状况[J].江西林业科技,2014,42(6):26-30.
[14]何福基.杉木种子涩粒成因的初步研究[J].种子,1985,14(6):6-8.
Characteristic of Plant Diversity,Seed Rain and Soil Seed Banks on Low-efficiencyPinusmassonianaLamb.Transformation Forest
WANG Li-yan,LIU Guang-zheng,YUE Jun-wei,LONG Wei,JIN Hong-ping,YANG Hua
(JiangxiAcademyofForestry,Nanchang330013,Jiangxi,China)
In north-central Wannian County Jiangxi Province after transforming the about 25 years low-efficiency ofPinusmassonianaforest as the research object,was used the plot set investigation of forest vegetation type,and used the collector and the ground seed collector from the ground,the sample analysis after transforming masson pine forests schima superba and characteristics ofCunninghamialanceolataseed rain and soil seed Banks and vertical distribution patterns.The results showed that modified the ioefficiency ofPinusmassonianaforest,forests trees is given priority toSchimasuperbaandCunninghamialanceolata.From the point of vertical distribution,three sampling,SchimasuperbaandCunninghamialanceolatasoil seed Banks′ total distribution shows the tendency of gradually reduce from top to bottom.The total amount of litterSchimasuperbaseed bank were more than 70% of the sampling number of seed collection,0 to 5 cm in soil seed bank amount per unit area in between 4 to 11 grain·m-2,every time sampling,seed regardless of number or no life force ratio is higher.In three times sampling,the average amounus of schima superba seed bank are 49.6,24.8,23 grain·m-2.The soil seed bank ofCunninghamialanceolata(Lamb.)Hook litter layer and soil layer 0~5 cm and 5~10 cm three levels mean life vigor seed density respectively is 8.77,1.23,3.50 grain·m-2,accounts for the proportion of the total average of the seed bank 53.02%,14.82% and 45.05%
Low-efficiency forests ofPinusmassonianaLamb.;transformation;plant diversity;seed rain;soil seed banks
2015-04-13;
2015-05-15
江西省林业科学院博士启动基金项目(江西省典型阔叶林修复类型的种子雨与土壤种子库动态研究,2011520802);江西省林业财政专项资金项目(江西省红壤低丘低效针叶林改造技术集成与示范,2011510801);国家科技支撑项目(鄱阳湖生态经济区建设生态环境保护关键技术研究及示范子课题:人工针叶林结构优化技术集成研究,2012BAC11B05-1)
王丽艳(1980—),女,山西文水人,江西省林业科学院助理研究员,博士,从事森林生态方面的研究。E-mail:wangliyan052@163.com。
10.13428/j.cnki.fjlk.2016.01.001
S756.5;S718.5
A
1002-7351(2016)01-0001-08
猜你喜欢
中国林副特产(2022年4期)2022-08-08 05:50:30
散文诗世界(2022年5期)2022-05-23 02:02:55
水资源开发与管理(2019年11期)2019-12-02 09:09:16
绿色科技(2019年5期)2019-11-29 13:17:37
防护林科技(2019年12期)2019-04-14 04:01:18
河北林业科技(2016年5期)2016-11-08 03:12:58
河北林业科技(2016年3期)2016-10-27 08:02:44
花卉(2016年14期)2016-07-31 18:46:24
广西林业科学(2016年1期)2016-03-20 05:33:00
广西林业科学(2016年1期)2016-03-20 05:32:59
