
小果叶下珠花气味组成及其两性异型性
2016-08-29 02:39:05黄代红陈国平杨晓飞李后魂石福臣
生态学报 2016年13期
黄代红,陈国平,杨晓飞,李后魂,石福臣
南开大学生命科学学院,天津 300071
小果叶下珠花气味组成及其两性异型性
黄代红,陈国平,杨晓飞,李后魂,石福臣*
南开大学生命科学学院,天津300071
大戟科(Euphorbiaceae)植物小果叶下珠(Phyllanthusmicrocarpus)由种特异性细蛾科(Gracillariidae)昆虫头细蛾(Epicephala)专门为其传粉,具体包括:头细蛾在雄花上积极采粉,然后为雌花授粉并在其内产卵等极其不同的传粉行为。花气味在维持小果叶下珠与传粉头细蛾专性传粉互利共生关系中起着至关重要的作用。采用动态顶空吸附法分别收集小果叶下珠雌花和雄花气味,利用气相色谱-质谱(gas chromatography-mass spectrometry, GC-MS)联用技术分析鉴定其成分,并用峰面积归一法与内标法进行定量,最后通过主成分分析法比较两者间的差异性。结果表明:(1)小果叶下珠花气味中共分离出17种化合物,主要以萜类和脂肪族物质为主;顺-β-罗勒烯在雌花和雄花中含量均最高,为主要气味成分;(2)雌花气味释放量显著高于雄花;(3)雌花和雄花之间气味化学成分存在明显的差异,即具有两性异形性。初步推测花气味出现两性异形性是植物为适应传粉头细蛾极其高度特异的传粉行为(雄花采粉,雌花传粉并产卵)而选择进化的结果。
专性传粉;互利共生关系;小果叶下珠;头细蛾;花气味;两性异型性
植物与昆虫之间互利共生关系中最为特化的例子就是植物-种子寄生性传粉昆虫专性传粉互利共生关系[1-5]。在此类传粉系统中,昆虫为植物传粉同时,并在其繁殖器官内产卵[6]。最经典的模式有榕树-榕小蜂[7-9]和丝兰-丝兰蛾系统[2,5]。此外,细蛾科(Gracillariidae)头细蛾属(Epicephala)昆虫与大戟科(Euphorbiaceae)算盘子属(Glochidion),叶下珠属(Phyllanthus)和黑面神属(Breynia)植物也存在类似的专性传粉互利共生关系[3,10-12]。
花气味在植物-昆虫专性传粉互利共生体系中具有非常重要的生态功能,它作为宿主植物释放的一种化学信号,可以强烈地吸引种特异性传粉者,介导传粉昆虫定位到宿主,促使双方相遇,进而寻求各自所需的报酬[13]。小果叶下珠(Phyllanthusmicrocarpus)为雌雄同株,单性花。在夜晚,由种特异性头细蛾专门为其传粉:首先,头细蛾在雄花上采集花粉,然后为雌花传粉,之后在授过粉的雌花中产卵。孵化的幼虫仅消耗一部分种子即可完成发育,剩余的种子可确保宿主能够繁育后代。这样双方之间形成了专性传粉互利共生关系[10]。然而,小果叶下珠花小且颜色不鲜艳(图1),加之头细蛾一般在夜间活动,进而推测花气味可能作为一种化学信号,在夜晚来吸引传粉头细蛾定位到小果叶下珠植株,是维持小果叶下珠与传粉头细蛾之间高度的种特异性所必须的。因此,分析小果叶下珠花气味的化学组成是非常有必要的。
两性异型性在动物界普遍存在,主要指两性间所表现出的差异,包括个体大小、形态以及颜色间的不同,有时甚至包括生理、行为和生活史等方面的差异[14-15]。在被子植物中,雌雄异花植物均由两性花植物进化而来,将近占被子植物的30%[16]。在雌雄异花的虫媒植物中,雌花和雄花产生的气味通常都是均质的,不具有两性异型性。其主要原因有两点:(1)雌花和雄花均吸引同一传粉者,两性间相似的花气味可以确保传粉者携带花粉顺利完成雄花到雌花的种内转移,成功完成传粉[17-19];(2)在一般的传粉系统中,传粉仅仅是一个副产品,传粉者为植物传粉同时,主要是从雌花和雄花间所寻求相同的报酬,比如取食花蜜[20]。然而,在大戟科植物-传粉头细蛾育幼传粉系统中发现一种很有趣的现象,传粉头细蛾在宿主植物上表现出明显不同的传粉行为(报酬):在雄花上积极采粉,然后为雌花传粉并在其内产卵[3,10-12]。宿主植物为了确保头细蛾在其上完成这两种高度特异且完全不同的传粉行为,雌花和雄花可能会选择进化出不同的气味。因此,推测小果叶下珠雌花和雄花的气味之间可能存在差异,即两性异型性。
基于以上假设,本研究利用动态顶空吸附法分别收集小果叶下珠雌花和雄花气味挥发物,运用GC-MS技术分析吸引传粉头细蛾的特异性化学信号的组成,并且运用主成分分析法比较雌花和雄花的气味之间化学成分的差异性,来初步探讨维持小果叶下珠和传粉头细蛾专性传粉互利共生关系的化学生态机制。
1 材料和方法
1.1实验材料
2013年3月底至4月初于广西省凭祥市夏石镇梢平林场附近公路两旁的小果叶下珠植株 (图1)上分别采集盛花期的雌花和雄花:雌花500—600朵;雄花800朵。

图1 小果叶下珠Fig.1 P. microcarpus
1.2方法
1.2.1花气味收集
(1)花气味收集采用动态顶空吸附法进行花气味收集[21]。主要工作流程:将采集好的花装入无味透明的聚氟乙烯采集袋中(2 L,Teflon®FEP,大连德霖气体包装有限公司),袋两端分别留有进气口和出气口。首先,用空气泵(FAY4002,成都气海机电制造有限公司)将空气通入活性碳管(外径6 mm,长75 mm,Sigma-Aldrich公司,美国)内进行净化,之后经由采集袋进气口进入袋中,促使袋内花朵气味挥发,最后用空气泵将挥发的气味经由采集袋出气口抽出,导入Tenax TA(100 mg /50mg, 60/80目,Sigma-Aldrich公司,美国)吸附管(外径6 mm,长75 mm)中进行吸附。通气端和出气端空气流量分别用玻璃转子流量器(LZB-4,天津流量仪表有限公司)控制,流速均为400 mL/min。连接装置器件之间均用无味的特氟隆管(PTEE,VICI Jour公司,瑞士)连接。花气味收集于18:30—23:30进行,持续5 h。在采样过程中,用空气作为对照,与花气味收集同步进行。采集完毕后,用锡箔纸包裹吸附管并装入密封袋内,置于超低温冰盒带回实验室。
(2)花气味洗脱及浓缩用正己烷(色谱纯,Fisher公司,美国)反复洗脱吸附管3次,收集约0.5 mL洗脱液于2 mL棕色进样瓶中。为进行样品的定量分析,加入 500 ng正二十烷(色谱纯,Sigma-Aldrich公司,美国)作为内标化合物,然后氮吹浓缩至100L,最后将样品置于冰箱内(-18 ℃)保存,直至进行仪器分析时取出。
1.2.2花气味化学分析
采用美国安捷伦科技公HP6890 (GC)-5973(MS)气-质联用仪进行气味挥发物的化学分析。
(1)GC条件色谱柱为HP-5MS石英毛细管柱(30 m×0.25 mm×0.25 μm);进样量2 μL,采用分流模式,分流比4∶1;柱箱程序升温40 ℃ 保持 5 min,然后以3 ℃/min升温至 150 ℃,最后以10 ℃/min升至200 ℃,保持5 min;进样口温度250 ℃;载气为高纯He,流速1.0 mL/min。
(2)MS条件离子轰击源设为扫描功能,扫描范围35—500 amu,电离方式EI;电子能量70 eV;接口温度280 ℃;离子源温度 230 ℃;四级杆温度150 ℃。
(3)挥发物的鉴定采用仪器自带的标准谱库(NIST 08)检索定性确认挥发物的化学成分。运用峰面积归一化法计算气味各组分的相对百分含量。
1.3数据统计分析
所有数据均表示为平均值±标准差。采用Graphad Prism 5. 0软件,对雌花和雄花气味释放量利用非配对样本t检验比较两者之间的差异性;采用SPSS17.0软件,对小果叶下珠雌花和雄花气味化学成分进行主成分分析,并利用非参数检验 Kruskal-Wallis test对两者主成分进行差异显著性检验。图片均采用 Graphad Prism 5. 0软件制作完成。
2 结果与分析
2.1花气味化合物的组成
运用GC-MS对小果叶下珠花气味成分进行化学分析,共鉴定出17种化合物(表1),其中反-β-罗勒烯在雌花和雄花中含量均最高,分别挥发20.8 pg 和3.76 pg,各占相对百分含量的26.77%和17.97%,是花气味的主要成分。此外,雌花气味中还主要含有α-金合欢烯(16.61%)、顺乙酸-3-己烯酯(15.97%)以及顺-3-己烯-1-醇(10.92%)等物质;雄花气味中还主要含有顺乙酸-3-己烯酯(11.23%)、苯甲醚 (10.79%)以及β-榄香烯 (9.98%)等物质。
在检测到的17中挥发物中,雌花和雄花各自所特有的挥发物仅3种,雌花:1-己醛、乙酸己酯和顺-异戊酸-3-己烯酯;雄花:顺-β-罗勒烯、β-榄香烯和香树烯。两者共同含有的多达12种。因此,雄花和雌花气味化合物的组成模式基本相似。
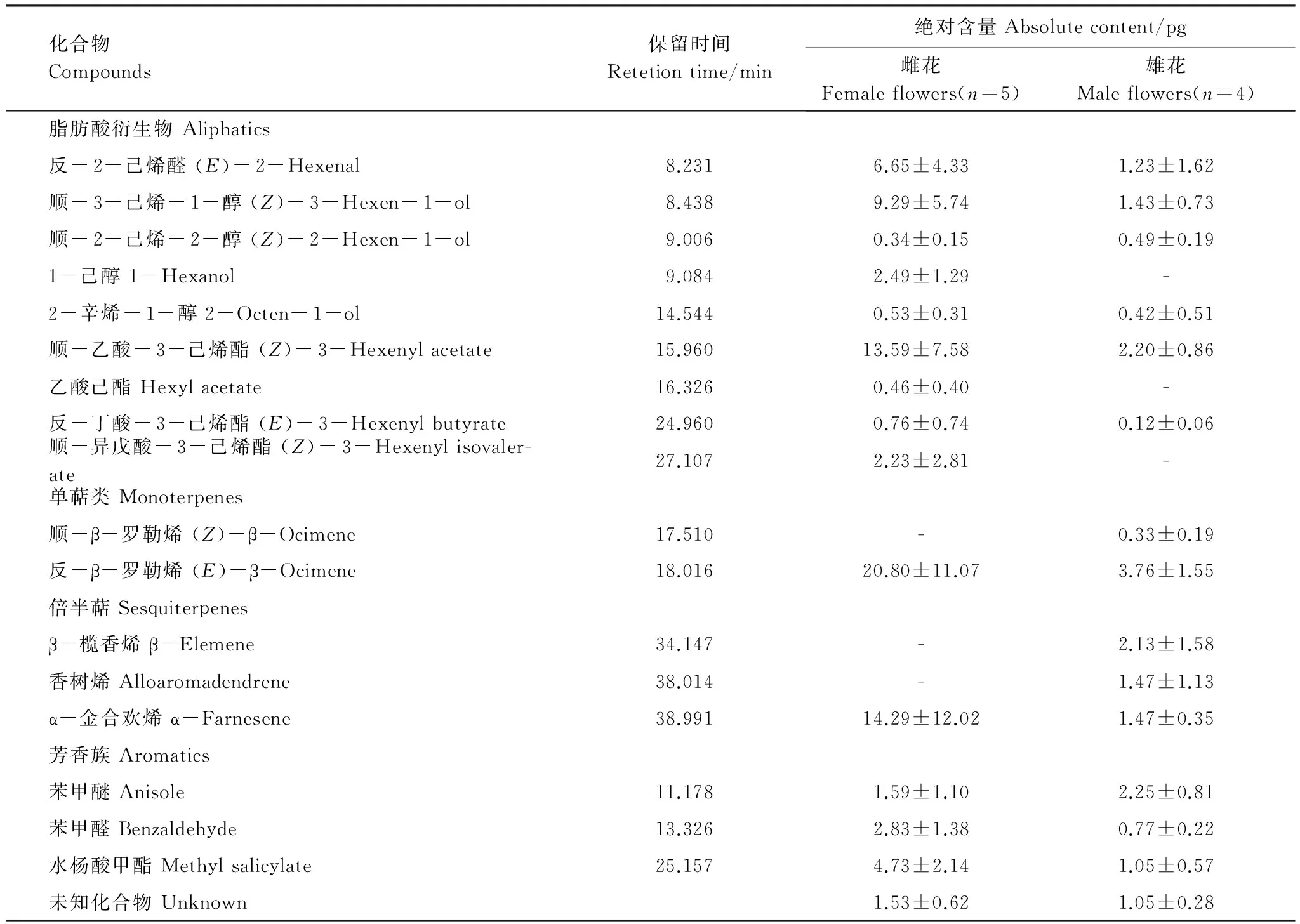
表1 小果叶下珠花气味挥发物的化学组成Table 1 The chemical composition of floral volatiles of both female and male flowers inP.microcarpus
2.2花气味的化合物类别
小果叶下珠花气味中共包含3种类型的化合物,包括脂肪酸衍生物、萜类(单萜类和倍半萜类)和芳香族化合物。从图2可以看出:雌花和雄花中,均主要以萜类和脂肪族化合物为主;两大类挥发物相对含量高达:87.81%和75.1%。此外,芳香族化合物在两种花气味中含量均最少。
2.3花气味挥发物的释放量
用内标法分别对小果叶下珠雌花和雄花气味中各个组分进行定量,得到单位时间内单朵花挥发物的总挥发量,如图3所示。雌花的挥发量高达(81.11±22.66)pg/h;而雄花仅为(20.68±3.10)pg/h,雌花的释放量显著高于雄花(t=5.21,P<0.005)。结果表明小果叶下珠雌花和雄花气味存在明显的量上的差异。
2.4雌花和雄花气味差异性分析
对小果叶下珠花气味顶空样品(雄花4;雌花5)中17种化合物进行主成分分析,其结果如图 4 所示。主成分1和2的累计方差累计贡献率达到 64%,可认为这两主成分包含了小果叶下珠花气味中主要信息。小果叶下珠雄花和雌花气味顶空样品各自聚合在一起,两者之间互相分离,没有交叉或重叠。对两个主成分进行非参数Krushal-Wallis 检验,结果显示,用主成分1和2确实可以将雄花和雌花的气味明显区分开 (χ2=6,P=0.014),以上结果说明小果叶下珠雄花和雌花的气味化学成分存在明显的差异,即具两性异型性。

图2 小果叶下珠雌花和雄花气味不同类别化合物的相对含量Fig.2 Relative content of different chemical classes of the floral scents from female and male flowers in P. microcarpus
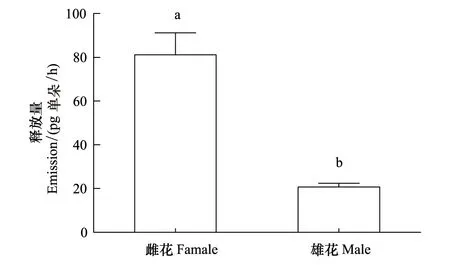
图3 小果叶下珠雄花和雌花气味挥发物总释放量Fig.3 Total emission of scent compounds from male flower and female flower of P. microcarpus图中不同小写字母代表差异达到显著水平,P<0.05

图4 小果叶下珠雌花和雄花气味化合物的主成分分析 Fig.4 Principal component analysis (PCA) of the scent compounds from the female and male flowers in P. microcarpus
3 讨论
3.1小果叶下珠花气味特异性信号及其组成特征
花气味是由植物花瓣及腺体释放的一类具有香气的挥发性物质,作为一种古老媒介,在植物与昆虫的相互关系中至关重要[22]。比如,传粉昆虫可通过识别植物气味来寻找“特异”植株[23]。本研究从小果叶下珠花气味中分离出17种挥发物。其中,顺-β-罗勒烯在雌花和雄花中含量均最高,是主要成分(表1)。与头细蛾传粉相关的大戟科植物种类繁多[4,24],但目前已鉴定出花气味化学成分的仅2科6种植物[25-26]。已有研究显示,小叶黑面神(Breyniavitis-idaea)花气味主要物质是2-苯乙腈和2-苯乙醇;香港算盘子(Glochidion.zeylanicum)中主要含有反-β-罗勒烯,而披针叶算盘子(G.lanceolatum)中以芳樟醇为主,台闽算盘子(G.rubrum),倒卵叶算盘子(G.obovatum)以及里白算盘子(G.acuminatum)主要为顺-β-罗勒烯和有芳樟醇[25-26]。除顺-β-罗勒烯外,以上4种主要气味物质均在小果叶下珠花中没有检测到。此外,在鉴定的17种化合物中,多达9种物质为小果下珠特有,在以上6种植物花中均无发现。因此,尽管到目前为止报道花气味的植物数量有限,但通过以上对比发现,小果叶下珠花气味与已报道的6种同科植物花气味化学组成之间存在着一定的差异,具有种特异性。然而,还需进一步开展触角电生理分析和昆虫行为学实验来鉴定特异性吸引小果叶下珠传粉头细蛾的活性物质。
Visser 提出,植物挥发物都通常以两种方式实现其特异性:(1)依靠一些常见的化合物,通过不同种类,依照不同比例混合形成,即称为“泛化”策略。(2)依赖一种或者某几种稀有化合物实现特异性,即称为“特化”策略[27]。Knudsen等归纳总结了利用顶空动态法收集的38目90科991物种,共发现1700 多种花香化合物[28]。Tomoko等研究表明,5种算盘子植物花气味中42种化合物,高达30种存在其他开花植物中[25,28]。Svensson等研究显示,吸引小叶黑面头细蛾的主要物质2-苯乙醇和2-苯乙腈也分别广泛存在于263种和68种植物中[26,28]。因此,已报道花气味的与传粉头细蛾相关的大戟科植物均采取了“泛化”策略实现其气味特异性,来吸引各自的传粉者。然而,到目前为止,鸡嗉子榕(Ficussemicorda)是植物与昆虫专性传粉互利共生体系中唯一种通过稀有化合物4-苯甲基醚,通过建立“私密通道”吸引其种特异性传粉者Ceratosolengravelyi为其进行传粉的植物[29]。在小果叶下珠花气味中共分离出17中化合物(表1),雌雄气味花的主要物质顺-β-罗勒烯不但存在于同科植物算盘子属植物和黑面神中[26-27],而且还广泛存在于其他43科的植物花中[25]。另外,其它挥发物也存在于其他植物中[25]。因此,认为小果叶下珠花气味主要是利用一些常见化合物,通过一定的比例混合形成其特异性“化学指纹图谱”,来区别同科其他植物,介导传粉头细蛾找到宿主,这样就维持了小果叶下珠和传粉头细蛾之间专性传粉互利共生关系的高度种特异性。
3.2小果叶下珠花气味两性异型性
在一般的动物-植物(雌雄异花)传粉系统中,植物雌花和雄花释放的气味通常相似,主要是由于它俩均吸引同一传粉者。为确保传粉者携带花粉在雌花和雄花之间的顺利完成转移,成功为植物授粉,植物会选择进化出相似的雌花与雄花气味信号[17-19]。此外,当植物花一方(通常为雄花)可为传粉者提供报酬(取食花粉)或者繁殖场所,而另一方(通常为雌花)则不提供任何报酬。此时,无报酬方雌花通常会化学模拟释放与雄花相似的气味,来欺骗引诱传粉者为其授粉[30]。两性间气味化学模拟现象在榕树[19,28,31-32 ]和苏铁科植物[33-35]中普通存在。然而,本研究发现,小果叶下珠雌花与雄花间气味存在着明显差异,即花气味具有两性异型性(图4)。同样,由头细蛾专性传粉的小叶黑面神雌花与雄花之间气味化学成分也存在分歧[27]。就雌雄异花的小果叶下珠而言,雌花和雄花在形态结构上存在明显的差异(图1),两性间花形态结构的这种差异可能会导致雌花与雄花产生的气味不均一[29]。然而,Svensson等[27]提出一种适应性假设:传粉头细蛾在宿主小叶黑面神雌花和雄花上表现出明显不同且高度程序化的传粉行为:先在雄花采粉;然后为雌花传粉并在其内产卵。推测宿主植物为了适应传粉头细蛾这种如此高度特异的传粉行为,可能会选择进化产生不同的花香,分别吸引头细蛾完成极其不同的传粉行为。头细蛾在小果叶下珠和黑面神上的所表现出相似的传粉行为[10-11]。因此,本文也支持以上适应性假设,认为小果叶下珠雌花和雄花气味存在两性异型性很可能是为适应头细蛾这种高度特异的传粉行为所致。然而,还需要进一步对非细蛾传粉的大戟科植物花气味两异型性分析以及相关头细蛾行为学等实验对于此假设加以验证。
此外,大多数昆虫为植物传粉同时,主要是为了从中寻求一定的报酬。植物雌花和雄花所提供的报酬不同可促使花信号两性异型性的产生[20]。传粉头细蛾在小果叶下珠雄花上可以取食花粉作为食物报酬,然而,雌花可以为其提供产卵场所,可供其后代发育。因此,小果叶下珠雄花和雌花为头细蛾提供的报酬不同,导致其花气味产生两性异型性。
3.3小果叶下珠花气味释放量的差异
本实验结果显示,单位时间内,小果叶下珠单朵雌花释放量显著高于雄花(图3)。通过野外观测,小果叶下珠雌花尺寸明显大于雄花,推测雌雄间花朵尺寸大小的差异是可能导致小果叶下珠雌花气味释放量高于雄花的原因之一[30]。然而,Waelti等研究显示,在白玉草(Silenelatifolia)-夜蛾(Hadenabicruris)育幼传粉系统中,白玉草雄花虽小,但释放的气味明显高于雌花,导致传粉夜蛾更偏好选择雄花,雄花可以吸引更多的传粉者[36]。因此,还需要对小果叶下珠花气味吸引传粉头细蛾生物学行为学实验来进一步分析导致小果叶下珠花气味释放量的差异的原因。
4 结论
采用动态顶空吸附法收集小果叶下珠花气味,运用GC-MS分析鉴定其化学成分,并利用主成分分析法比较雄花和雌花气味之间的差异:
(1)小果叶下珠花气味中共鉴定17种挥发物,以萜类和脂肪族物质为主,雌花与雄花主要成分均为顺-β-罗勒烯,这些化合物形成吸引小果叶下珠头细蛾特异性的化学信号,与已报道的6种头细蛾相关植物的花气味之间存在明显的差异,这样维持了小果叶下珠与头细蛾专性传粉互利共生关系的高度种特异性。
(2)小果叶下珠花气味具有两性异型性,雌花和雄花释放不同的气味,推测是为了适应头细蛾在宿主上的高度特异的传粉行为进化而产生,确保头细蛾先在雄花上采粉,然后转移至在雌花上传粉并产卵,从而保证了两者之间专性互利关系的稳定性。
研究表明:小果叶下珠花气味及其两性异型性对维持其与传粉头细蛾专性传粉互利共生体系具有重要的化学生态学意义。
致谢:西南林业大学李宗波老师对实验给予帮助,中国林业科学研究院热带林业研究中心梢平林场工作人员对野外采集工作提供支持,特此致谢。
[1]Daniel H Janzen. How many babies do figs pay for babies?. Biotropica, 1979, 11(1):48-50.
[2]Olle Pellmyr, James Leebens-Mack, Chad J Huth. Non-mutualistic yucca moths and their evolutionary consequences. Nature, 1996, 380(6570):155-156.
[3]George D Weiblen. How to be a fig wasp. Annual Review of Entomology, 2002, 47:299-230.
[4]Makoto Kato, Atsushi Takimura, Atsushi Kawakita. An obligate pollination mutualism and reciprocal diversification in the tree genusGlochidion(Euphorbiaceae). Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(9):5264-5267.
[5]Olle Pellmyr. Yuccas, yucca moths, and coevolution:a review. Annals of the Missouri Botanical Garden, 2003, 90(1):35-55.
[7]Marie Charlotte Anstett, Martine Hossaert-McKey, Finn Kjellberg. Figs and fig pollinators:evolutionary conflicts in a coevolved mutualism. Trends in Ecology & Evolution, 1997, 12(3):94-99.
[8]James M Cook, Jean-Yves Rasplus. Mutualists with attitude:coevolving fig wasps and figs. Trends in Ecology & Evolution, 2003, 18(5):241-248.
[9]Edward Allen Herre, K Charlotte Jandér, Carlos Alberto Machado. Evolutionary ecology of figs and their associates:recent progress and outstanding puzzles. Annual Review of Ecology, Evolution and Systematics, 2008, 39:439-458.
[10]Atsushi Kawakita, Makoto Kato. Evolution of obligate pollination mutualism in New CaledonianPhyllanthus(Euphorbiaceae). American Journal of Botany, 2004, 91(3):410-415.
[11]Atsushi Kawakita, Makoto Kato. Obligate pollination mutualism inBreynia(Phyllanthaceae):further documentation of pollination mutualism involvingEpicephalamoths (Gracillariidae). American Journal of Botany, 2004, 91(9):1319-1325.
[12]Zhang J, Wang S X, Li H H, Hu B B, Yang X F, Wang Z B. Diffuse coevolution between twoEpicephalaspecies (Gracillariidae) and twoBreyniaspecies (Phyllanthaceae). PLoS ONE, 2012, 7(7):e41657.
[13]Martine Hossaert-McKey, Catherine Soler, Bertrand Schatz, Magali Proffit. Floral scents:their roles in nursery pollination mutualisms. Chemoecology, 2010, 20(2):75-88.
[14]Charles Darwin. The Descent of Man, and Selection in Relation to Sex. Princeton:Princeton University Press, 1871.
[15]Fairbairn D J, Roff D A. The quantitative genetics of sexual dimorphism:assessing the importance of sex-linkage. Heredity, 2006, 97:319-328.
[16]David G Lloyd, C J Webb. Secondary sex characters in plants. The Botanical Review, 1977, 43(2):177-216.
[17]Charles B Fenster, W Scott Armbruster, Paul Wilson, Michele R Dudash, James D Thomson. Pollination syndromes and floral specialization. Annual Review of Ecology, Evolution and Systematics, 2004, 35:375-403.
[18]Mary F Willson, Jon Agren. Differential floral rewards and pollination by deceit in unisexual flowers. Oikos, 1989, 55(9):23-29.
[19]Catherine C L Soler, Magali Proffit, Jean-Marie Bessière, Martine Hossaert-McKey, Bertrand Schatz. Evidence for intersexual chemical mimicry in a dioecious plant. Ecology Letters, 2012, 15(9):978-985.
[20]Åsa M Hemborg, William J Bond. Different rewards in female and male flowers can explain the evolution of sexual dimorphism in plants. Biological Journal of the Linnean Society, 2005, 85(1):97-109.
[21]Robert A Raguso, Olle Pellmyr. Dynamic headspace analysis of floral volatiles:a comparison of methods. Oikos, 1998, 81(Fasc. 2):238-254.
[22]Olle Pellmyr. Three pollination morphs inCimicifugasimplex:incipient speciation due to inferiority in competition. Oecologia, 1986, 68(2):304-307.
[23]Eran Pichersky, Jonathan Gershenzon. The formation and function of plant volatiles:perfumes for pollinator attraction and defense. Current Opinion in Plant Biology, 2002, 5(3):237-243.
[24]Atsushi Kawakita. Evolution of obligate pollination mutualism in the tribe Phyllantheae (Phyllanthaceae). Plant Species Biology, 2010, 25(1):3-19.
[25]Tomoko Okamoto, Atsushi Kawakita, Makoto Kato. Interspecific variation of floral scent composition inGlochidion(Phyllanthaceae) and its association with host-specific pollinating seed pasrasite (Epicephala; Gracillariidae). Journal of Chemical Ecology, 2007, 33(5):1065-1081.
[26]Glenn P Svensson, Tomoko Okamoto, Atsushi Kawakita, Ryutaro Goto, Makoto Kato. Chemical ecology of obligate pollination mutualisms:testing the ‘private channel’ hypothesis in theBreynia-Epicephalaassociation. New phytologist, 2010, 186(4):995-1004.
[27]Visser J H. Host Odor Perception in phytophagous insects. Annual Review of Entomology, 1986, 31:121-144.
[28]Jette T Knudsen, Roger Eriksson, Jonathan Gershenzon, Bertil Ståhl. Diversity and distribution of floral scent. The Botanical Review, 2006, 72(1):1-20.
[29]Chen C, Song Q S, Magali Proffit, Jean-Marie Bessière, Zongbo Li, Martine Hossaert-McKey. Private channel:a single unusual compound assures specic pollinator attraction inFicussemicordata. Functional Ecology, 2009, 23(5):941-950.
[30]Tia-Lynn Ashman. Snifng out patterns of sexual dimorphism in floral scent. Functional Ecology, 2009, 23(5):852-862.
[32]李宗波, 杨培, 彭艳琼, 杨大荣. 木瓜榕隐头果传粉前后挥发性化合物构成及其变化规律. 云南大学学报(自然科学版), 2012, 34(1):90-98.
[33]Irene Terry, Paul I Forster, Chris J Moore, Robert B Roemer, Peter J Machin. Demographics, pollination syndrome and conservation status ofMacrozamiaplatyrhachis(Zamiaceae), a geographically restricted Queensland cycad. Australian Journal of Botany, 2008, 56(4):321-332.
[34]Serban Proches, Steven D Johnson. Beetle pollination of the fruit-scented cones of the South African cycadStangeriaeriopus. American Journal of Botany, 2009, 96(9):1722-1730.
[35]Terence N Suinyuy, John S Donaldson, Steven D Johnson. Variation in the chemical composition of cone volatiles within the African cycad genusEncephalartos. Phytochemistry, 2013, 85:82-91.
[36]Maelti O Waelti, Paul A Page, Alex Widmer, Florian P Schiestl. How to be an attractive male:floral dimorphism and attractiveness to pollinators in a dioecious plant. BMC Evolutionary Biology, 2009, 9:190.
Analysis of the composition and sexual dimorphism of floral scent inPhyllanthusmicrocarpus
HUANG Daihong, CHEN Guoping, YANG Xiaofei, LI Houhun, SHI Fuchen*
CollegeofLifeScience,NankaiUniversity,Tianjin300071,China
Obligate pollination mutualisms between plants and their seed-parasitic pollinators are known to be one of the most specialized plant-insect interactions. The obligate pollination mutualism betweenEpicephalamoths (Gracillariidae) andPhyllanthusmicrocarpusplants (Phyllanthaceae) has been documented.P.microcarpusspecies need to rely on species-specificEpicephalafemale moths for obligate pollination. At night, a female moth actively collects pollen grains from the host male flower by using unique proboscises equipped with numerous hair, and then deposits them on the female flower; subsequently, the female moth lays an egg by using the specialized acicular ovipositor. The hatched larva consumes only a subset of the maturing seeds within a single fruit to complete its development and drills out of the fruit. The rest of the seeds ensure the reproduction of the host′s offspring. The floral scent has been considered to be the key signal guiding obligate pollinators to locate host plants in nursery pollination mutualisms; however, studies on floral volatiles ofP.microcarpusare lacking. In this study, the chemical composition of floral volatiles of both female and male flowers fromP.rmicrocarpuswas analyzed using the dynamic headspace technique and gas chromatography-mass spectrometry (GC-MS). Whether there are qualitative differences in floral scent between the sexes ofP.microcarpuswas determined by conducting principal component analysis (PCA). The results showed that (1) a total of 17 compounds were detected and identified from the floral scent ofP.microcarpus, which were dominantly terpenoid and aliphatic compounds. (E)-β-Ocimene was found in the highest amounts in both female (26.77%) and male flowers (17.97%) and was thus considered as the major scent component ofP.microcarpus. Of the 17 identified volatiles, 12 occurred in both female and male flowers, indicating similar chemical composition patterns of floral scent between both the sexes ofP.microcarpusis obviously different from other Phyllanthaceae plants whose floral scent has been identified in the chemical composition of floral scent. (3) Female flowers emitted significantly more amounts of odor than male flowers. (4) The PCA revealed thatP.microcarpusspecies showed qualitative differences in chemical composition of floral scent between male and female flowers, indicating sexual dimorphism in floral scent. These results indicated that the species-specific floral scent ofP.microcarpusmight mediate the obligate pollinatorEpicephalamoth to find the host in order to maintain in the high species specificity ofP.microcarpus-Epicephalamutualism. The sexual dimorphism in floral scent ofP.microcarpusmight be an important reason for the host to adapt to the highly specializedEpicephalapollination.
obligate pollination; mutualism;Phyllanthusmicrocarpus;Epicephala; floral scent; sexual dimorphism
10.5846/stxb201411212307
国家自然科学基金重点资助项目(30930014)
2014-11-21; 网络出版日期:2015-10-29
Corresponding author.E-mail:fcshi@nankai.edu.cn
黄代红,陈国平,杨晓飞,李后魂,石福臣.小果叶下珠花气味组成及其两性异型性.生态学报,2016,36(13):4013-4020.
Huang D H, Chen G P, Yang X F, Li H H, Shi F C.Analysis of the composition and sexual dimorphism of floral scent inPhyllanthusmicrocarpus.Acta Ecologica Sinica,2016,36(13):4013-4020.
猜你喜欢
作文周刊·小学二年级版(2023年48期)2024-01-01 21:25:24
农业科技通讯(2023年1期)2023-02-12 07:09:08
中国科学探险(2022年8期)2022-12-31 07:27:54
金沙江文艺(2022年4期)2022-04-26 14:14:20
蚕桑通报(2022年3期)2022-04-10 03:16:00
疯狂英语·新悦读(2022年12期)2022-03-08 02:37:12
花火B(2021年3期)2021-06-25 02:38:31
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25 01:40:26
上海农业学报(2017年3期)2017-04-10 12:39:22
天然产物研究与开发(2016年6期)2016-06-05 10:29:27
