
川西高山林线交错带凋落叶分解初期转化酶特征
2016-08-30 06:02:13陈亚梅和润莲邓长春宋小艳刘军伟
生态学报 2016年13期
陈亚梅, 和润莲, 刘 洋, 张 健, 邓长春, 宋小艳, 杨 林, 刘军伟
长江上游林业生态工程重点实验室,四川农业大学生态林业研究所,高山森林生态系统定位研究站,长江上游生态安全协同创新中心,成都 611130
川西高山林线交错带凋落叶分解初期转化酶特征
陈亚梅, 和润莲, 刘洋*, 张健, 邓长春, 宋小艳, 杨林, 刘军伟
长江上游林业生态工程重点实验室,四川农业大学生态林业研究所,高山森林生态系统定位研究站,长江上游生态安全协同创新中心,成都611130
胞外酶对于有机质的降解具有重要的作用。在凋落物分解过程中,酶活性不仅受到凋落物种类或基质质量的影响,还受到环境因素的影响。转化酶催化蔗糖水解为葡萄糖和果糖,因此在凋落物分解早期,转化酶比降解难分解物质的酶具有更重要的作用。以川西高山林线交错带12种代表性凋落叶为研究对象,对林线交错带不同植被类型下的凋落叶转化酶活性以及物种和环境因子对转化酶活性的影响进行了研究。结果表明:同一植被类型下,12个物种转化酶活性具有极显著差异(P<0.01)。物种、环境因子及其交互作用对转化酶活性有极显著的影响(P<0.01)。初始纤维素含量与转化酶活性极显著正相关(P<0.01)。初始木质素和总酚含量与转化酶活性极显著负相关(P<0.01),能够共同解释转化酶活性变异的50.8%。不同植物生活型中,禾草类转化酶活性均为最高,这可能与禾草类较高的初始纤维素含量、较低的木质素和总酚含量有关。多元线性回归分析表明,凋落叶含水量能单独解释转化酶活性变量的62.1%,是环境因子中最重要的变量。从植被类型来看,大多数物种的转化酶活性在针叶林中均极显著高于高山草甸和灌丛(P<0.01),这可能与针叶林中凋落叶的含水量最高且雪被最厚有关。历经一个雪被期分解后,凋落叶初始质量与环境因子的综合作用能够解释转化酶活性变异的79.1%,表明川西高山林线交错带凋落叶分解前期转化酶活性主要受初始木质素含量、总酚含量和含水量的调控。在全球气候变化情景下,凋落物水分含量的变化将会强烈的影响凋落叶分解前期的转化酶活性。
转化酶;林线交错带;凋落物质量;含水量;凋落物分解
凋落物分解过程中的胞外酶对于有机质的降解具有至关重要的作用[1]。胞外酶主要来源于微生物、土壤动物以及植物活体组织(主要为植物的根系)分泌以及死亡细胞的释放[2-3]。一般而言,凋落物分解过程与胞外酶尤其是与难分解物质降解相关的纤维素酶、木聚糖酶和木质素酶有密切的关联[4]。凋落物分解前期,主要是凋落物易分解组分的分解[5], 因此参与简单碳化合物的分解的转化酶对于凋落物早期分解能发挥更重要的作用[6]。凋落物分解过程中的酶活性不仅受到凋落物种类或基质质量的影响,还受到环境因子的影响[7-9]。
研究发现,不同种类植物的凋落物在其分解过程中酶的活性具有较大差异[10-11]。一方面,不同凋落物初始质量的差异,尤其是木质素、纤维素和半纤维素以及果胶相对含量的差异,通过酶-基质的相互作用导致酶活性差异[1]。另外,基质质量影响微生物群落[12],诱导其释放特定的胞外酶,胞外酶对基质作用后又改变基质质量[1]。
大量研究表明,酶活性与环境中的温度和水分有关[13-15]。一方面,在一定范围内, 酶的活性与温度存在极强的正相关关系,水热条件会直接影响到酶活性[13,16];另一方面,温度和水分可通过影响微生物进而影响酶活性[17]。研究表明,酶活性与真菌生物量及活性有很好的相关性[18]。在高山区域,雪被是最重要的环境因子之一,覆盖时间长达半年以上[19-20]。总体来看,在高山地区雪被梯度生成了温度和水分梯度[21-22]。除此之外,土壤的冻融循环也在凋落物的冬季分解过程中扮演着重要的角色[20]。
高山-亚高山是研究气候变化的关键区域,监控这些敏感和脆弱的生态系统对反映气候变化有重要价值[23]。高山林线交错带从暗针叶林过渡到高山草甸,不仅凋落物种类发生很大变化,且不同植被类型对降雪分配的差异可能造成系统内温度、湿度的急剧变化,从而很大程度上影响凋落物的分解过程[24]。邓长春等研究发现,高山林线交错带不同植被类型下的雪被差异是导致冬季凋落物可溶性有机碳淋失的主要环境因素[25]。杨文彬等对漓江水陆交错带不同植被类型土壤酶活性进行了研究,发现植被类型影响土壤酶活性,且土壤水分含量显著影响土壤酶活性的变化[26]。由于影响转化酶活性的因素较多,因此在凋落物分解的初期,不同种类或生活型的凋落物的转化酶活性特征和影响因素还存在不确定性。经历一个雪被期的分解后,高山林线交错带的凋落物转化酶活性在不同植被类型下有何不同,哪些因素是影响酶活性的重要因子?目前还缺乏对林线交错带凋落物分解早期转化酶活性的研究,这不利于人们全面理解凋落物分解过程。因此,本文以川西高山林线交错带12种代表性凋落叶为研究对象,研究林线交错带不同植被类型下一个雪被期分解后的凋落叶转化酶活性特征及其影响因素,以期为深入理解高山生态系统凋落物分解过程及其影响机制提供基础数据。
1 研究区域
研究区位于四川省阿坝州理县米亚罗林区,地理位置为31°43′N—31°51′N,102°40′E—103°02′E,位于青藏高原东缘,杂谷脑河上游,是大渡河与岷江的分水岭,为重要的江河源区。鹧鸪山位于杂谷脑河北侧,山体海拔3200—4800 m,具有明显的山地垂直地带性,自河谷至山顶依次分布的植被类型有针阔混交林、暗针叶林、高山疏林灌丛、高山草甸,4500 m以上为高山荒漠和积雪带。气候属于冬寒夏凉的高寒气候,年平均气温6—12 ℃,1月平均气温-8 ℃,7月平均气温12.6 ℃,年积温1200—1400 ℃,年降水量600—1100 mm,年蒸发量1000—1900 mm。冬季雪被期明显,从10月到次年4月,长达6—7个月。林线交错带的植被类型变化主要为针叶林、高山灌丛和高山草甸。从暗针叶林到高山灌丛和草甸,光照逐渐增强,蒸发量逐渐增大,地被物厚度逐渐降低。暗针叶林内植被发育良好,完全郁闭,生境阴冷潮湿,以高山杜鹃(Rhododendronlapponicum)-岷江冷杉(Abiesfargesiivar.faxoniana)林为主要景观,土壤类型为山地假灰化棕色针叶林土,土壤腐殖质含量高,但多系粗腐殖质,未很好矿质化[27]。疏林灌丛和高山草甸的土壤类型为高山草甸土,地表石砾含量较高,灌木优势种有红毛花楸(Sorbusrufopilosa)、柳叶忍冬(Loniceralanceolata)、康定柳(Salixparaplesia)、越桔叶忍冬(Loniceramyrtillusvar.myrtillus)、峨眉蔷薇(Rosaomeiensis)、大理杜鹃(Rhododendrontaliense)、华西小檗(Berberissilva-taroucana)、金露梅(Potentillafruticosa);草本优势种主要有禾本科、菊科和龙胆科植物[28]。
2 研究方法
2.1样地设置和样品采集
研究样地位于米亚罗鹧鸪山,地理位置为31°51′428″ N, 102°41′230″ E。采用GPS定位,沿垂直于等高线方向,据高山林线交错带的植被类型变化分别设置3个海拔梯度的固定样方——针叶林3900 m,高山灌丛4000 m,高山草甸4200 m。2012年10月初,于高山林线交错带植物凋落高峰期收集12种代表性植物(康定柳Salixparaplesia、红毛花楸Sorbusrufopilosa、高山杜鹃Rhododendronlapponicum、圆叶筋骨草Ajugaovalifolia、藏羊茅Festucawallichanica、柳兰Epilobiumangustifolium、糙野青茅Deyeuxiascabrescens、草血竭Polygonumpaleaceum、冷地早熟禾Poacrymophila、高山冷蕨Cystopterismoupinensis、锦丝藓Actinothuidiumhookeri、草甸马先蒿Pedicularisroylei)新鲜凋落叶,于自然条件下风干后65 ℃烘干至恒量。称取12种烘干后的凋落叶10 g装入尼龙网材质、大小20 cm×20 cm,网孔大小为底部(贴地面)0.5 mm表面层为1 mm[25,29]的凋落物袋中,每个物种制备12袋,共制备144袋备用。每个物种取3袋,共36袋,磨碎并分析凋落叶初始基质质量。2012年10月底将凋落物袋放回林线交错带的3个样带的3个固定样方,凋落袋平铺于样地地表,各凋落袋间隔至少5 cm。每个固定样方中选择一个凋落物袋,将一个纽扣式温度记录器(iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, USA)放置于选择的凋落物袋中,以便记录凋落叶的分解温度。设定每3h记录1次温度。于2013年4月30日在3个植被类型下收取每个物种的分解袋3个,供酶活性和含水量的测定,采集的凋落袋用封口袋封装,装入伴有冰袋的保鲜盒中,翌日带回实验室,于4 ℃冰箱保存,一周内进行相关分析测试。采样同时读取温度计记录数据,日平均分解温度见图 1。雪被覆盖时期,每个月定期对每个样方(5个点)内雪被厚度进行测量,计算每个海拔的平均雪被厚度。冻融循环频次计算参考Zhu等[30],具体为按照持续时间≥3 h的零上温度交替到零下温度并持续3 h及以上,或持续时间≥3 h的零下温度交替到零上温度并持续3 h及以上,计为冻融1次。

图1 分解期间高山草甸、高山灌丛和针叶林的凋落叶分解日平均温度(共分解184 d)Fig.1 Dynamic of daily decomposition temperature of alpine meadow, alpine shrub and coniferous forest from October 29, 2012 to April 30, 2013 (totaling 184 days of exposure)
2.2分析方法
取部分样品(约凋落袋样品中的三分之一)于烘箱中,在65 ℃烘干至恒重,进行凋落物含水量的测定,剩余样品进行酶活性测定。酶活性采用先提取后测定的方法,由于转化酶不仅来源于微生物的分泌,还可能部分来源于植物本身[31],因此本研究提取的是凋落叶中的所有转化酶,不区分来源。
粗酶液的制备参考张瑞清等[6]修改的Criquet等[32]的方法,并做了部分修改,具体方法如下: 称取4.00 g被剪碎的样品(<0.5 mm)于冰浴研钵中,加入20 mL预冷(4 ℃, 12 h) 的提取液(1 mol/L CaCl2溶液,其中含有0.5 mL tween 80),迅速研磨成匀浆,转移至50 mL 离心管中,加入0.40 g预冷的交联聚乙烯吡咯烷酮(PVPP),摇动后放置过夜,12000 g 4 ℃冷冻离心20 min,将上清液转移到预处理过的透析袋(截留分子量为10 kD)中,置于2 mmol/L的Bis-Tris缓冲液(pH 6. 0)中,在4 ℃透析48 h,期间,每12 h更换1次透析液。 将粗酶液转移到刻度为15 mL试管中定容,4 ℃保存备用。
转化酶活性的测定参考张瑞清等[6]的方法。具体方法如下:0.4ml 粗酶液和0.6%的蔗糖的醋酸缓冲液(pH 5.5, 0.2 mol/L)于50℃水浴3h。生成的还原糖量采用3, 5-二硝基水杨酸(DNS)还原法测定。设失活酶对照,重复测3次。酶活力单位表达为μmol 葡萄糖 g-1干重 h-1。
凋落叶全碳含量采用重铬酸钾氧化-外加热法测定。全氮含量采用半微量凯氏定氮法测定,全磷含量采用钼锑钪比色法测定。纤维素和木质素含量采用改进的范氏酸性洗涤纤维法[33]测定。凋落叶可溶性有机碳含量采用王春阳等[34]的测定方法,总酚的测定参考福林-酚法[35]。凋落叶初始质量的测定结果见表1。

表1 高山林线交错带12个物种凋落叶初始质量特征(平均值±标准差)Table 1 Initial litter quality of 12 typical plants in alpine timberline ecotone (mean±SD)

续表
2.3统计分析
采用SPSS 17.0对不同植被类型同一物种的转化酶活性和同一植被类型下不同物种转化酶活性进行单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)检验。采用双因素方差分析(Two-way ANOVA)检验物种和植被类型及其交互作用对转化酶活性的影响。采用Pearson相关分析检验转化酶活性与初始凋落叶质量和环境因子的相关关系。用多元线性回归(逐步进入法)分析初始凋落叶质量和环境因子对转化酶活性的影响。
3 结果与分析
3.1凋落叶转化酶活性特征
同一植被类型类型下,12物种转化酶活性差异极显著(P<0.01)。从不同生活型来看,禾本科草本转化酶活性在3个植被类型下均为最高(表2)。不同植被类型下,锦丝藓、柳兰、糙野青茅、藏羊茅的转化酶活性均为针叶林>高山草甸>高山灌丛(P<0.01),圆叶筋骨草、冷地早熟禾、草甸马先蒿和草血竭的转化酶活性为针叶林>高山草甸和高山灌丛(P<0.01),高山冷蕨、高山杜鹃转化酶活性为高山灌丛>高山草甸和针叶林(P<0.01)。不同植被类型下,红毛花楸的转化酶活性为高山草甸>针叶林>高山灌丛(P<0.01)(图2)。不同生活型凋落叶中,锦丝藓、阔叶草本和禾本科草本的转化酶活性均为针叶林>高山草甸和高山灌丛(P<0.01)。木本植物的转化酶活性在不同植被类型下无显著差异(P>0.05) (表2)。

表2 不同植物生活型凋落叶转化酶活性Table 2 Invertase activity of different life form leaf litter
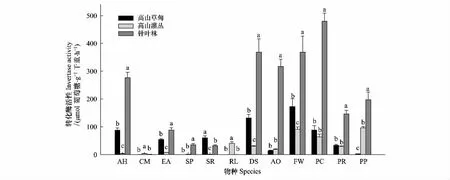
图2 12个物种凋落叶的转化酶活性Fig.2 Leaf litter invertase activity of 12 species level among vegetation typesAH: 锦丝藓Actinothuidium hookeri; CM: 高山冷蕨Cystopteris moupinensis; EA: 柳兰 Epilobium angustifolium; SP: 康定柳 Salix paraplesia; SR:红毛花楸 Sorbus rufopilosa; RL:高山杜鹃Rhododendron lapponicum; DS:糙野青茅Deyeuxia scabrescens; AO: 圆叶筋骨草Ajuga ovalifolia; FW: 藏羊茅Festuca wallichanica; PC:冷地早熟禾Poa crymophila; PR: 草甸马先蒿Pedicularis roylei; PP: 草血竭Polygonum paleaceum; 不同小写字母表示同一物种在不同植被类型下差异显著 Different lowercase letters of the same species meant significant difference at 0.05 among vegetation types
3.2物种和植被类型对转化酶活性的影响
物种和植被类型对转化酶的双因素方差分析结果显示(表3):植被类型、物种及其交互作用均对转化酶活性有极显著的影响(P<0.01)。

表3 植被类型和物种及其交互作用对12种凋落叶转化酶活性的双因素方差分析Table 3 Two-way ANOVA of vegetation type, species and their interaction on the leaf litter invertase activity of 12 species
凋落叶初始C含量与针叶林和整个交错带的转化酶活性负相关(P<0.01)。除了高山草甸外,初始纤维素含量和木质素/纤维素分别与转化酶活性极显著正相关和负相关(P<0.01)。初始木质素含量与不同植被类型下的转化酶活性均为负相关关系(P<0.01,P<0.05)。除高山灌丛外,转化酶活性随着初始可溶性有机碳和总酚含量的升高而降低(P<0.01,P<0.05)(表4)。多元线性回归分析结果显示(表5),总酚和木质素含量极显著影响转化酶活性,能够共同解释酶活性变异的50.8%。

表4 转化酶活性与12种凋落叶初始质量的相关性分析Table 4 Pearson correlation analysis of invertase activity and leaf litter quality of 12 species

表5 初始凋落叶质量和环境因子对转化酶活性的多元线性回归分析Table 5 Regressions of invertase activity with initial litter quality variables and environmental factors

表6 高山林线交错带雪被期凋落叶含水量及主要环境参数Table 6 The average moisture content of litter and main environmental parameters in alpine timberline ecotone
环境因子显著影响转化酶活性。针叶林凋落叶平均含水量显著高于高山灌丛和高山草甸(P<0.05)。针叶林中雪被最厚,冻融循环次数最少且凋落叶平均分解温度显著低于高山灌丛和高山草甸(P<0.05)(表6)。凋落叶平均分解温度和冻融循环次数与转化酶活性极显著负相关(P<0.01),凋落叶含水量和平均雪被厚度与转化酶活性极显著正相关(P<0.01)(表7)。多元线性回归分析结果表明(表5),含水量能单独解释酶活性变异的62.1%,含水量与平均分解温度的共同作用能解释转化酶活性64.4%的变异,初始凋落叶质量(木质素、纤维素和总酚的含量)与环境因子(含水量和平均分解温度)的综合作用能够解释酶活性变异的79.1%。

表7 转化酶活性与环境因子的相关性分析Table 7 Pearson correlation analysis of invertase activity and environmental factors
4 讨论
4.1物种/不同生活型凋落叶对转化酶活性的影响
研究表明,物种强烈的影响凋落物酶活性的总体水平[10,36]。本研究中物种对转化酶活性有极显著的影响(P<0.01),同一植被类型下,12个物种转化酶活性差异极显著(P<0.01)(图2),与Kshattriya等[37]的研究结果一致。研究表明,转化酶部分地来源于植物体本身[31],因此转化酶活性差异可能与凋落叶质量差异有关。此外,不同凋落物初始质量的差异,尤其是木质素、纤维素和半纤维素以及果胶相对含量的差异,通过酶-基质的相互作用导致酶活性差异[1]。
凋落叶质量因子中,纤维素含量、木质素含量、木质素/纤维素、可溶性有机碳和总酚含量能显著影响转化酶活性,其中木质素和总酚含量能共同解释转化酶活性变异的50.8%,这与Criquet[38]、Sinsabaugh等[39]和Fioretto等[40]的研究结果基本一致。凋落叶中总酚含量越高,转化酶活性越低,这是由于酚类物质对转化酶有抑制作用[41]。凋落物的木质素代表了最丰富的芳族聚合物,也是最难分解的物质之一[13,42]。Jiang等[43]研究也发现凋落物中的木质素是影响凋落物分解的关键组分。相较于木质素而言,纤维素更易分解[44],较高的木质素含量(木质素/纤维较高),表示凋落物的可分解性程度越低,进而影响分解初期微生物对基质的利用,导致转化酶活性较低。研究表明,在凋落物分解过程中纤维素是最初被分解的化合物之一[40]。在纤维素分解过程中,纤维素分解酶能够从葡聚糖链的末端移除单体和二聚体[45],而转化酶能够参与纤维素降解过程,催化蔗糖水解为葡萄糖和果糖。因此在凋落叶分解早期纤维素含量越高,转化酶活性越高。这与张瑞清[6]和季晓燕等[46]的研究结果一致,凋落物的分解并不是通过某个酶单独起作用的,而是多种酶联合作用的结果。本研究中同一植被类型下,禾草类转化酶活性均为最高(表2),这可能与禾本科草本较高的初始纤维素含量、较低的木质素和总酚含量有关。
4.2环境因子对酶活性的影响
凋落物分解过程中的胞外酶活性受到温度、水分等环境因子的影响[15,40,47],尤其在寒冷生态系统,温度和水分是凋落物分解的主要驱动力[48]。在高山地区,土壤温度、水分以及土壤冻结与雪被的厚度和持续时间紧密相关[49]。高山林线交错带在短距离内存在着不同植被类型的交错分布[50],不同植被类型导致的雪被差异使得凋落物分解的水热条件在较短的距离内具有明显的差异。通过对环境因子的分析发现,湿度比温度对转化酶活性的影响更大,凋落叶含水量能单独解释酶活性变异的62.1%。具体来看,凋落叶含水量与转化酶活性极显著正相关,平均分解温度与转化酶活性极显著负相关,与徐振锋等[51]模拟增温(+0.61℃和+0.56℃)增加了土壤转化酶活性结果不一致。本文中,转化酶活性随着冻融循环次数的增加和雪被厚度的降低而降低。这是由于雪被厚度的降低导致雪被隔热能力下降和冻融循环次数的增加,使雪下生态系统经历更严苛冬季环境[52],从而影响土壤生物的增殖以及分泌的胞外酶活性[52-53]。研究表明,土壤水分含量被证明是影响微生物生物量和胞外酶空间分布最重要的因素之一[17],本研究也证明了这一点。土壤水分含量不仅可以影响微生物生物量和土壤呼吸作用,同时能够影响酶催化过程[13,38],因此含水量对于转化酶的影响较大。本研究中大多数物种和生活型在针叶林中的转化酶活性均极显著高于高山草甸和灌丛(P<0.01),这可能是由于针叶林中湿度最大且雪被最厚,为凋落叶的酶解过程提供了一个稳定的环境。
5 结论
物种和植被类型及其交互作用极显著影响凋落叶转化酶活性。凋落叶初始纤维素含量、木质素含量和总酚含量对转化酶的影响较大。含水量为环境因子中最重要的变量,能单独解释变量的62.1%。凋落叶质量与环境因子的综合作用能够解释酶活性变异的79.1%,表明川西高山林线交错带凋落叶分解前期转化酶活性主要受到凋落叶本身的质量和环境因子的调控。
从不同植被类型来看,大多数物种的转化酶活性在针叶林中均极显著高于高山草甸和灌丛,这可能与针叶林中凋落叶的含水量最高且雪被最厚有关。可以推测,在全球气候变化情景下凋落物水分含量的变化将会强烈的影响凋落叶分解前期的转化酶活性。
[1]Fioretto A, Papa S, Pellegrino A, Fuggi A. Decomposition dynamics ofMyrtuscommunisandQuercusilexleaf litter: Mass loss, microbial activity and quality change. Applied Soil Ecology, 2007, 36(1): 32-40.
[ 2]杨万勤, 张健. 土壤生态研究. 成都: 四川科学技术出版社, 2008.
[ 3]Sinsabaugh R L, Antibus R K, Linkins A E. An enzymic approach to the analysis of microbial activity during plant litter decomposition. Agriculture, Ecosystems & Environment, 1991, 34(1-4): 43-54.
[ 4]Stemmer M, Gerzabek M H, Kandeler E. Invertase and xylanase activity of bulk soil and particle-size fractions during maize straw decomposition. Soil Biology and Biochemistry, 1998, 31(1): 9-18.
[ 6]张瑞清, 孙振钧, 王冲, 袁堂玉. 西双版纳热带雨林凋落叶分解的生态过程. III. 酶活性动态. 植物生态学报, 2008, 32(3): 622-631.
[ 7]Allison S D, Vitousek P M. Extracellular enzyme activities and carbon chemistry as drivers of tropical plant litter decomposition. Biotropica, 2004, 36(3): 285-296.
[ 8]张东来, 毛子军, 张玲, 朱胜英. 森林凋落物分解过程中酶活性研究进展. 林业科学, 2006, 42(1): 105-109.
[ 9]Sinsabaugh R L, Lauber C L, Weintraub M N, Ahmed B, Allison S D, Crenshaw C, Contosta A R, Cusack D, Frey S, Gallo M E, Gartner T B, Hobbie S E, Holland K, Keeler B L, Powers J S, Stursova M, Takacs-Vesbach C, Waldrop M P, Wallenstein M D, Zak D R, Zeglin L H. Stoichiometry of soil enzyme activity at global scale. Ecology Letters, 2008, 11(11): 1252-1264.
[10]Linkins A E, Sinsabaugh R L, McClaugherty C A, Melills J M. Cellulase activity on decomposing leaf litter in microcosms. Plant and Soil, 1990, 123(1): 17-25.
[11]Kourtev P S, Ehrenfeld J G, Häggblom M. Exotic plant species alter the microbial community structure and function in the soil. Ecology, 2002, 83(11): 3152-3166.
[12]Knapp B A, Rief A, Seeber J. Microbial communities on litter of managed and abandoned alpine pastureland. Biology and Fertility of Soils, 2011, 47(7): 845-851.
[13]Criquet S, Farnet A M, Tagger S, Le Petit J. Annual variations of phenoloxidase activities in an evergreen oak litter: influence of certain biotic and abiotic factors. Soil Biology and Biochemistry, 2000, 32(11-12): 1505-1513.
[14]Butenschoen O, Scheu S, Eisenhauer N. Interactive effects of warming, soil humidity and plant diversity on litter decomposition and microbial activity. Soil Biology and Biochemistry, 2011, 43(9): 1902-1907.
[16]徐振锋, 尹华军, 赵春章, 曹刚, 万名利, 刘庆. 陆地生态系统凋落物分解对全球气候变暖的响应. 植物生态学报, 2009, 33(6): 1208-1219.
[17]Baldrian P, Merhautová V, Cajthaml T, Petránková M,najdr J. Small-scale distribution of extracellular enzymes, fungal, and bacterial biomass inQuercuspetraeaforest topsoil. Biology and Fertility of Soils, 2010, 46(7): 717-726.
[18]Andersson M, Kjøller A, Struwe S. Microbial enzyme activities in leaf litter, humus and mineral soil layers of European forests. Soil Biology and Biochemistry, 2004, 36(10): 1527-1537.
[19]Edwards A C, Scalenghe R, Freppaz M. Changes in the seasonal snow cover of alpine regions and its effect on soil processes: A review. Quaternary International, 2007, 162-163: 172-181.
[20]Walker M D, Walker D A, Welker J M, Arft A M, Bardsley T, Brooks P D, Fahnestock J T, Jones M H, Losleben M, Parson A N, Seastedt T R, Turner P L. Long-term experimental manipulation of winter snow regime and summer temperature in arctic and alpine tundra. Hydrological Processes, 1999, 13(14-15): 2315-2330.
[21]Bryant D M, Holland E A, Seastedt T R, Walker M D. Analysis of litter decomposition in an alpine tundra. Canadian Journal of Botany, 1998, 76(7): 1295-1304.
[22]Seastedt T R, Walker M D, Bryant D M. Controls on decomposition processes in alpine tundra//Bowman W, Seastedt T, eds. Structure and Function of an Alpine Ecosystem. Oxford: Oxford University Press, 2001: 222-236.
[23]刘洋, 张健, 闫帮国, 黄旭, 徐振锋, 吴福忠. 青藏高原东缘高山森林——苔原交错带土壤微生物生物量碳、氮和可培养微生物数量的季节动态. 植物生态学报, 2012, 36(5): 382-392.
[24]Sjögersten S, Wookey P A. Decomposition of mountain birch leaf litter at the forest-tundra ecotone in the Fennoscandian mountains in relation to climate and soil conditions. Plant and Soil, 2004, 262(1-2): 215-227.
[25]邓长春, 陈亚梅, 和润莲, 刘洋, 张健, 宋小艳, 李建平. 雪被期川西高山林线交错带凋落物可溶性有机碳的淋失特征. 生态学杂志, 2014, 33(11): 2921-2929.
[26]杨文彬, 耿玉清, 王冬梅. 漓江水陆交错带不同植被类型的土壤酶活性. 生态学报, 2015, 35(14): 4604-4612.
[27]刘洋, 张健, 杨万勤, 吴福忠, 黄旭, 闫帮国, 文维全, 胡开波. 川西高山树线群落交错带地被物及土壤的水文效应. 林业科学, 2011, 47(3): 1-6.
[28]陈亚梅, 和润莲, 邓长春, 刘洋, 杨万勤, 张健. 川西高山林线交错带凋落物纤维素分解酶活性研究. 植物生态学报, 2014, 38(4): 334-342.
[29]Keane R E. Biophysical controls on surface fuel litterfall and decomposition in the northern Rocky Mountains, USA. Canadian Journal of Forest Research, 2008, 38(6): 1431-1445.
[30]Zhu J X, He X H, Wu F Z, Yang W Q, Tan B. Decomposition ofAbiesfaxonianalitter varies with freeze-thaw stages and altitudes in subalpine/alpine forests of southwest China. Scandinavian Journal of Forest Research, 2012, 27(6): 586-596.
[31]Ross D J, Roberts H S. Biochemical activities in a soil profile under hard beech forest. I. Invertase and amylase activities and relationships with other properties. New Zealand Journal of Science, 1973, 16(1): 209-224.
[32]Criquet S, Tagger S, Vogt G, Iacazio G, Le Petit J. Laccase activity of forest litter. Soil Biology and Biochemistry, 1999, 31(9): 1239-1244.
[33]Rowland A P, Roberts J D. Lignin and cellulose fractionation in decomposition studies using acid-detergent fibre methods. Communications in Soil Science and Plant Analysis, 1994, 25(3-4): 269-277.
[34]王春阳, 周建斌, 王祥, 夏志敏. 黄土高原区不同植物凋落物可溶性有机碳的含量及生物降解特性. 环境科学, 2011, 32(4): 1139-1145.
[35]Makkar H P S, Siddhuraju P, Becker K. Plant Secondary Metabolites. New Jersey: Humana Press, 2007.
[36]Waring B G. Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest. Soil Biology and Biochemistry, 2013, 64: 89-95.
[37]Kshattriya S, Sharma G D, Mishra R R. Enzyme activities related to litter decomposition in forests of different age and altitude in North East India. Soil Biology and Biochemistry, 1992, 24(3): 265-270.
[38]Criquet S. Measurement and characterization of cellulase activity in sclerophyllous forest litter. Journal of Microbiological Methods, 2002, 50(2): 165-173.
[39]Sinsabaugh R L, Carreiro M M, Repert D A. Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss. Biogeochemistry, 2002, 60(1): 1-24.
[40]Fioretto A, Papa S, Curcio E, Sorrentino G, Fuggi A. Enzyme dynamics on decomposing leaf litter ofCistusincanusandMyrtuscommunisin a Mediterranean ecosystem. Soil Biology and Biochemistry, 2000, 32(13): 1847-1855.
[41]Mandels M, Reese E J. Inhibition of cellulases. Annual Review of Phytopathology, 1965, 3(1): 85-102.
[43]Jiang X Y, Cao L X, Zhang R D, Yan L J, Mao Y, Yang Y W. Effects of nitrogen addition and litter properties on litter decomposition and enzyme activities of individual fungi. Applied Soil Ecology, 2014, 80: 108-115.
[44]Kanazawa S, Miyashita K. Cellulase activity in forest soils. Soil Science and Plant Nutrition, 1987, 33(3): 399-406.
[45]Sánchez C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnology Advances, 2009, 27(2): 185-194.
[46]季晓燕, 江洪, 洪江华, 马元丹. 亚热带 3 种树种凋落叶厚度对其分解速率及酶活性的影响. 生态学报, 2013, 33(6): 1731-1739.
[47]Sinsabaugh R L, Linkins A E. Inhibition of theTrichodermaviridecellulase complex by leaf litter extracts. Soil Biology and Biochemistry, 1987, 19(6): 719-725.
[48]Aerts R. The freezer defrosting: global warming and litter decomposition rates in cold biomes. Journal of Ecology, 2006, 94(4): 713-724.
[49]Groffman P M, Hardy J P, Fashu-Kanu S, Driscoll C T, Cleavitt N L, Fahey T J, Fisk M C. Snow depth, soil freezing and nitrogen cycling in a northern hardwood forest landscape. Biogeochemistry, 2011, 102(1-3): 223-238.
[50]刘洋, 张健, 杨万勤. 高山生物多样性对气候变化响应的研究进展. 生物多样性, 2009, 17(1): 88-96.
[51]徐振锋, 唐正, 万川, 熊沛, 曹刚, 刘庆. 模拟增温对川西亚高山两类针叶林土壤酶活性的影响. 应用生态学报, 2010, 21(11): 2727-2733.
[52]Bokhorst S, Phoenix G K, Bjerke J W, Callaghan T V, Huyer-Brugman F, Berg M P. Extreme winter warming events more negatively impact small rather than large soil fauna: shift in community composition explained by traits not taxa. Global Change Biology, 2012, 18(3): 1152-1162.
[53]Orchard V A, Cook F J. Relationship between soil respiration and soil moisture. Soil Biology and Biochemistry, 1983, 15(4): 447-453.
Leaf litter invertase activity in the early stage of litter decomposition in alpine timberline ecotone of western Sichuan, China
CHEN Yamei, HE Runlian, Liu Yang*, ZHANG Jian, DENG Changchun, SONG Xiaoyan, YANG Lin, LIU Junwei
KeyLaboratoryofEcologicalForestryEngineeringinSichuanProvince,InstituteofEcology&Forestry,SichuanAgriculturalUniversity,Long-termResearchStationofAlpineForestEcosystems,CollaborativeInnovationCenterofEcologicalSecurityintheUpperReachesofYangtzeRiver,Chengdu611130,China
Extracellular enzyme plays a crucial role in litter decomposition for the degradation of organic matter. In the course of litter degradation, enzyme activity is not only influenced by the litter species or substrate quality, but also affected by environmental factors. Invertase catalyses the hydrolysis of sucrose to glucose and fructose; thus, invertase is more essential than enzymes related to recalcitrant material degradation in the early litter decomposition period. We investigated the invertase activity of 12 representative leaf litter in alpine timberline ecotone of western Sichuan, China. The effects of species and environmental factors on invertase activity were also studied. The results showed that invertase activities of 12 species were significantly different under the same vegetation type (P< 0.01). The effects of species, environmental factors, and their interactions on invertase activity were highly significant (P< 0.01). In addition, the relationship between the initial cellulose content and invertase activity was positively and extremely significant (P< 0.01). Moreover, there was a significant and negative correlation between invertase activity and initial lignin and total phenol content, respectively (P< 0.01). The lignin and total phenol content can explain the 50.8% variation in invertase activity. Invertase activity of graminoids was the highest among all plant life forms, which likely occurs due to the higher initial cellulose content and lower lignin and total phenol content of graminoids. Multiple linear regression analysis showed that leaf litter moisture can explain the 62.1% variation in invertase activity and was the most important environmental factor variable. From the perspective of vegetation type, the invertase activity of most species in the coniferous forest was significantly higher than that of species in the alpine meadow and shrubs (P< 0.01), which may be caused by the highest litter moisture content and snow depth in the coniferous forest. After a snow cover period decomposition, the comprehensive effect of leaf litter quality and environmental factors on invertase activity can explain the 79.1% variation in invertase activity, indicating that invertase activity was mainly controlled by lignin, total phenol content, and leaf litter moisture in the alpine timberline ecotone of western Sichuan. In the global climate change scenario, the change in moisture content of the litter will strongly influence invertase activity in the early stage of litter decomposition.
invertase; timberline ecotone; litter quality; moisture content; litter decomposition
10.5846/stxb201410242086
国家自然科学基金项目(31570605, 31200345);国家“十二五”科技支撑计划项目(2011BAC09B05);教育部博士点基金项目(20115103120003)
2014-10-24; 网络出版日期:2015-10-30
Corresponding author.E-mail: sicauliuyang@163.com
陈亚梅, 和润莲, 刘洋, 张健, 邓长春, 宋小艳, 杨林, 刘军伟.川西高山林线交错带凋落叶分解初期转化酶特征.生态学报,2016,36(13):4099-4108.
Chen Y M, He R L, Liu Y, Zhang J, Deng C C, Song X Y, Yang L, Liu J W.Leaf litter invertase activity in the early stage of litter decomposition in alpine timberline ecotone of western Sichuan, China.Acta Ecologica Sinica,2016,36(13):4099-4108.
猜你喜欢
上海包装(2019年8期)2019-11-11 12:16:14
现代农业研究(2018年4期)2018-08-11 10:25:02
中国绿色画报(2018年5期)2018-08-07 07:12:26
森林工程(2018年4期)2018-08-04 03:23:16
珠江水运(2017年17期)2017-10-10 08:16:03
天津造纸(2016年1期)2017-01-15 14:03:29
中国造纸学报(2015年1期)2015-12-16 19:35:25
中央民族大学学报(自然科学版)(2015年4期)2015-06-11 03:41:42
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06 07:49:06
中国塑料(2014年4期)2014-10-17 03:00:45
