
吉林省天然黄檗种群遗传多样性ISSR分析
2016-08-29 03:29:22李绍臣李凤明张立民林玉梅
生态学报 2016年13期
李绍臣, 李凤明, 张立民, 任 军, 林玉梅
吉林省林业科学研究院, 长春 130033
吉林省天然黄檗种群遗传多样性ISSR分析
李绍臣, 李凤明*, 张立民, 任军, 林玉梅
吉林省林业科学研究院, 长春130033
黄檗(PhellodendronamurenseRupr.)是吉林省长白山林区珍贵用材树种和主要建群树种,由于过度采伐和利用,其资源数量和质量明显下降。依据其资源自然分布现状,选择了10个具有代表性的天然黄檗分布种群,应用ISSR标记技术对其进行了遗传多样性的分析,以期为黄檗种群资源的收集、保存和保护提供依据和支持。研究结果表明:从60条ISSR引物中筛选出扩增99条带,多态性条带数为54条,多态性比率为54.5%。10个种群的多态位点比率分布在18.52%—37.96%范围内,其中珲春种群的多态位点比率最高,为37.96%,吉林省露水河种群的多态位点比率最低为18.52%,种群的平均多态位点百分比为26.02%。利用Shannon指数与Nei指数可较好的估算黄檗种群间的遗传变异,Shannon指数的变化范围在0.1103—0.1949之间,Shannon指数总体平均值为0.1522。 Nei指数的变化范围在0.0759—0.1327之间, 平均为0.1043。根据Nei法计算黄檗10个种群遗传多样性是Dst=0.1586,分化指数Gst=0.6183,基因流系数Nm为0.3086,总的遗传变异中有61.83%的变异存在于群体间,群体内的变异只占38.17%,种群间存在明显分化。黄檗的10个种群可分为两个大群,即:①松江河、露水河、湾沟、集安、辉南②白石山、汪清、安图、延吉、珲春。根据黄檗的遗传结构提出了保护措施:适度引导营造药用或用材林;开展本地黄檗资源的本底调查并进行资源汇总(包括林班、小班,每株的树龄、树高、胸径、枝下高和冠幅等数据),筛选本地的优势群体进行原地保存;迁地保护策略中要增加样本的数量,白山地区迁地保护的种源应选择松江河、露水河种源,通化地区迁地保护的种源应选择集安种源,而延边地区应选择白石山和汪清种源;人工促进黄檗的天然林更新改造,逐步恢复黄檗种群规模,并且进行人工更新的资源登记。
黄檗;遗传多样性;ISSR
黄檗(黄柏,黄菠萝PhellodendronamurenseRupr.),属芸香科黄檗属落叶阔叶乔木,是东北地区地带性植被顶级群落——阔叶红松林的主要伴生树种、主要的用材树种和药用植物[1]。由于过度的利用,目前已成为国家级重点保护植物。黄檗在吉林省主要分布在东部山区,包括延边、白山、通化、吉林等,目前对黄檗的研究多集中于育种造林[2-4]、植物化学[5-6]等方面, 有关遗传多样性方面的工作较少[7-8],闫志峰仅对吉林省的3个种群进行了AFLP分析。迄今对其遗传多样性和遗传结构知之甚少, 以至于无法提出有针对性的保护策略。
ISSR(Inter-Simple Sequence Repeat)技术[9-10]是由Zietkiewicz等于1994年创建的一种简单序列重复区间扩增多态性分子标记,近年来,已被广泛应用于遗传多样性和群体遗传结构的研究[11-14]。本研究采用先进的ISSR技术,运用群体遗传学原理,从DNA水平对黄檗天然种群的遗传多样性进行研究,揭示不同地区黄檗天然种群的遗传关系,探讨黄檗天然种群的遗传分化机制,为黄檗遗传资源的有效保护和合理利用提供理论依据。
1 材料与方法
1.1材料
黄檗在吉林省主要分布在东部山区,根据地理位置和吉林省森林资源调查文献以及种群分布的具体情况确定了10个有代表性的天然种群,采样时间是2013年3月在各采样点进行采样,每个地点采25个样本。为了使样本具有代表性,各个样本间相互距离要在100m以上,林分选择要求立地较好的近熟林分,采无病虫害的枝条拿回实验室水培,待叶子长出后采摘叶片提取DNA。10个天然种群的基本情况见表1。
1.2总DNA的提取
利用Universal genomic DNA extraction kit(Ver.3.0,TaKaRa, 大连)提取黄檗叶片总DNA。
1.3ISSR引物筛选
根据University of British Columbia(UBC)提供的ISSR引物序列,在上海生工公司合成60个2核苷酸重复的引物。每个种群随机选择1个DNA模板进行扩增筛选,从60个ISSR引物中筛选出10个能产生多态、清晰且可重复条带的引物,用于全部10个种群样本分析。各引物序号及序列见表2。
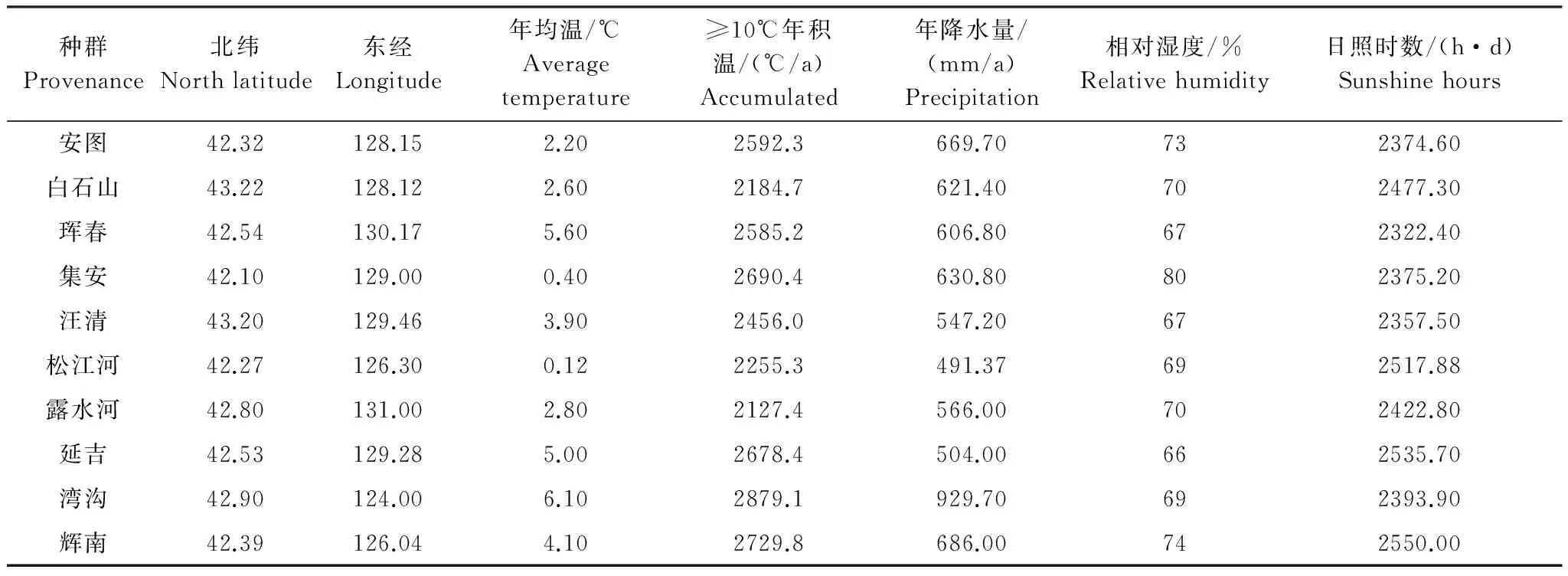
表1 10个种群黄檗的地理气候因子Table 1 Factors of geographical and climatic of 10 Provenance of P. amurense Rupr.

表2 ISSR引物序号与序列Table 2 Primer sequence number and sequence
1.4ISSR-PCR扩增及扩增产物的电泳检测
用筛选出的10个引物,对10个种群的250个DNA样本进行ISSR-PCR扩增,扩增产物用1×TAE电泳缓冲液(pH 8.0)在1.0%的琼脂糖凝胶中(含EB0.5μg/ml)电泳2h,电压为5V/cm,以分离扩增产物。DL2000(宝生物公司生产)作为分子量标准,电泳结果用Gene Genius公司的Bio Imaging System凝胶成像系统照相,记录结果。
1.5ISSR数据的统计分析
ISSR-PCR产物经电泳分离后,对扩增结果进行记录。具体的谱带记录方法目前无明确的标准,本研究参考邹喻平等的方法[15],以Marker产生带亮度为标准,亮的记为1,弱的、无带的记为0;建立0/1数据矩阵。为尽量减少误差,谱带结果的记录始终由试验者一人进行。得到的ISSR数据矩阵用于以下分析:1)将ISSR标记视为表征性状, 应用POPGENE软件(Version 1.3.1)计算多态位点百分率(PPL)和Shannon信息多样性指数(I);2)将ISSR标记视作同一基因座位上的2个等位基因,基于Hardy.Weinberg平衡,删除基因频率小于3/N(N为样本数,N=192)的条带后[16],分别计算Nei′s基因多样度指数(HE)、种群总基因多样度(H′)、种群内基因多样度(Hs)、基因分化系数(Gst)和Nei′s遗传距离(D);3)基于POPGENE软件对种群间Nei′s(1978)无偏遗传距离(unbiased genetic distance)的估算,用Mega2(molecular evolutionary genetics analysis)软件中的UPGMA聚类法以遗传距离对种群进行聚类分析。4)用AMOVA.PREP软件计算个体间欧氏距离所得的输出文件(距离文件、组文件、群体文件)作为输入文件,用WINAMOVA软件进行分子方差分析,并以种群为单位,采用Mantel检验分析10个种群间遗传距离和地理距离的相关性。
2 结果与分析
2.1黄檗地理种群的遗传多样性
2.1.1黄檗扩增产物的多态性
从60条ISSR引物中筛选出扩增条带清晰、重复性高的引物10条(表2)。ISSR扩增的条带在500bp—2500bp之间,图1为813扩增结果。10条引物共扩增出99条带,多态性条带数为54条,多态性比率为54.5%。

图1 引物813对部分材料进行扩增Fig.1 813 primers for amplification part materialM为标准分子量DL2000M is a standard molecular weight DL2000;1—20为部分个体1—20 for part of the individual
由表3可以看出,10个种群的多态位点比率分布在18.52%—37.96%范围内,其中珲春种群的多态位点比率最高,为34.26%,吉林省露水河种群的多态位点比率最低为18.52%,种群的平均多态位点百分比为26.02%。可见,种群间的多态位点百分比差距较大,珲春种群的多态位点百分比是露水河种群的2倍左右,说明珲春种群遗传基础较宽,具有较好的适应性。

表3 黄檗10个种群的多态性及遗传差异统计Table 3 The polymorphic loci and genetic diversity in 10 provenance of P. amurense Rupr.
2.1.2黄檗种群的Shannon信息指数分析和Nei遗传多样性指数分析
利用PopGen 1.32软件对黄檗种群进行遗传多样性分析,获得了黄檗总体的Shannon信息指数以及每个种群的Shannon信息指数。
由表3看出,黄檗总体的Shannon指数为0.1949,各个种群的Shannon多态性信息指数中,珲春种群最大,达到0.1949,露水河种群最小,为0.1103。Shannon指数总体平均值为0.1522。
所研究的黄檗每个种群的Nei指数分布在0.1327—0.0759范围内,平均为0.1043。所有种群根据Nei指数排列的顺序与根据Shannon指数排列的顺序基本一致。与按多态位点百分比排列的顺序有些差距,这主要是由于多态位点百分比居中的几个种群间百分比差距不大的原因。
2.1.3黄檗各种群间的遗传分化分析
根据总的遗传多样性(Ht)和群体内遗传多样性(Hs)计算不同群体间的遗传多样性(Dst,Dst=Ht-Hs)和遗传分化水平(Gst,Gst=Dst/Ht)。10个黄檗种群间的遗传多样性是Dst=0.1586,分化指数Gst=0.6183,基因流系数Nm为0.3086(表4)。

表4 不同种群间的遗传分化分析Table 4 Analysis of genetic differentiation among provenances
由结果可见,总的遗传变异中有61.83%的变异存在于群体间,群体内的变异只占38.17%,种群间的遗传变异占相当大的比重;基因流系数也远小于1,说明种群间存在明显的分化,因此黄檗种群的利用潜力很大。
2.2黄檗种群的聚类分析
为了进一步分析群体之间的遗传分化程度,计算了Nei′s的遗传距离(D)。遗传距离常用来衡量群体间的亲缘关系,遗传距离越大,说明群体间的亲缘关系越远;反之,遗传距离越小,群体间的亲缘关系越近。

图2 黄檗种群间遗传关系UPGMA聚类图 Fig.2 The genetic relationship between Huang Bo provenance UPGMA clustering figure
根据Nei′s遗传距离利用MEGA软件构建的群体遗传关系UPGMA聚类图见图2。聚类图中将黄檗的10个种群分为两个大群,即:①松江河、露水河、湾沟、集安、辉南②白石山、汪清、安图、延吉、珲春。其中,在群①中,又可分为2个小群,其中松江河、露水河和集安种群间的遗传距离较小,较早的聚为一支,湾沟和辉南种群间的遗传距离较小,较早的聚为一支;在群②中,又可分为2个小群,其中延吉、安图和珲春种群间的遗传距离较小,较早的聚为一支,白石山和汪清种群间的遗传距离较小,较早的聚为一支。
3 讨论
3.1黄檗遗传多样性的ISSR分析
遗传多样性是指存在于生物个体内、单个物种内以及物种之间的基因多样性。一个物种的遗传多样性说明了该物种在特定的环境因子作用下,其基因的丰富程度,而这是物种适应和进化的遗传基础。本文所检测到的108条多态位点的平均有效等位基因数目为1.1864,Shannon指数为0.1522,Nei指数为0.1043,低于闫志峰的1.2487,0.1524,0.2374[7]。遗传参数略低的原因是本文所用的种群都是来源于吉林省境内。单就这3个指标来说,黄檗的遗传多样度是中等的。但从多态位点百分率来看, 10个种群的多态位点比率分布在18.52%—37.96%范围内,其中珲春种群的多态位点比率最高,为34.26%,吉林省露水河种群的多态位点比率最低为18.52%,种群的平均多态位点百分比为26.90%。可见,种群内的多态位点比率相对总的多态位点比率小很多,但种群间的多态位点百分比差距较大,珲春种群的多态位点百分比是露水河种群的2倍左右,可以据此选择环境适应性强的种群。遗传变异在物种或是群体的分布形式及其时间上是以非随机方式变化的。遗传变异受突变、基因流、选择和遗传漂变的共同作用,同时还和物种的进化史和生物学特性有关[17-18]。本试验利用多态性位点分析来度量遗传变异水平的高低,而用Nei 法求出基因多样性,反映种群的多态性;利用Nei指数来估测实验获得的shomnon指数的变化范围来估测黄檗种群间及种群内的遗传分化。Nei指数(H*)和Shannon指数(I*)估算所得的10个黄檗种群的遗传变异结果基本是一致的,数据都表明在这10个种群中,遗传变异最高的是珲春种群,其次是湾沟种群,遗传变异最低的是露水河种群。遗传变异最高和最低的种群地理距离很远,遗传距离也较远。Nei基因分化系数为61.83%,Nei种群间基因多样性为15.86%,说明黄檗种群遗传差异较大,具有丰富的遗传基础。在本文中,应用ISSR分子标记技术,通过研究黄檗种群之间的遗传距离和聚类树形图,根据Nei′s遗传距离利用MEGA软件构建的群体遗传关系UPGMA聚类图。在0.20的遗传距离上将黄檗的10个种群分为两个大群,地理种群近的都很 好的聚在了一起,种群间遗传距离越小,说明这几个种群在遗传组成上越相似,亲缘较近。
3.2黄檗种群的保护策略
黄檗为名贵中药材和优质木材,其应用范围较广。黄檗的优势群体主要分布在河谷两侧的含腐殖质丰富的中性或微酸性冲积土,因此,其分布区较为狭窄,加之长期的认为干扰,造成生境的间隔化和种群数量变小,相互隔离的小种群使黄檗的遗传基础越来越窄, 而且本研究也证实了不同种群间的亲缘关系较近。根据黄檗的生存现状及遗传结构特点,提出保护策略:(1)结合禁止采伐的同时,应加强宣传教育力度, 严禁盗伐黄檗林木的行为,此外,可以适度引导营造药用或用材林,满足其供应和需求。(2)开展本地黄檗资源的本底调查并进行资源汇总(包括林班、小班号,每株的树高、胸径、枝下高和冠幅等数据),筛选本地的优势群体进行原地保存,既重视保护黄檗的高水平遗传多样性种群, 又兼顾低水平地区稀有基因的存在,保护黄檗种群的完整性。 (3)加强迁地保护,在迁地收集时应结合自然林和人工林的特殊群落特征,既保护其资源,又能丰富其遗传多样性,同时建立种质资源收集圃, 根据遗传结构进行合理取样,丰富现有种质资源。在迁地保护策略中要增加样本的数量,而且吉林省白山和通化地区迁地保护的种源应选择松江河、露水河和集安种源,而延边地区应选择白石山和汪清种源。(4)为防止该物种的遗传退化, 应积极创造适宜条件促进黄檗种子的原地萌发, 人工促进天然更新,逐步恢复黄檗种群规模,并且进行人工更新的资源登记。
[1]任宪威. 树木学. 北京: 中国林业出版社, 1997: 437-438.
[2]沙广义, 兰士波, 苏明炎, 李朝毅. 黄菠萝母树林改建技术及经营效果分析. 林业科技开发, 2007, 21(4): 54-57.
[3]鲁长虎, 常家传, 许青. 黄檗的更新特点及食果实鸟类对其种子的传播. 生态学杂志, 2004, 23(1): 24-29.
[4]刘军. 黄檗药用林栽培技术研究. 吉林林业科技, 2007, 36(1): 45-47.
[5]李霞, 王洋, 阎秀峰. 光强对黄檗幼苗三种生物碱含量的影响. 生态学报, 2009, 29(4): 1655-1660.
[6]张煊, 崔征, 周海燕, 董娥. 高效液相色谱法测定关黄柏不同采收期及黄檗不同部位的小檗碱、巴马汀含量. 沈阳药科大学学报, 2003, 20(3): 194-197.
[7]闫志峰, 张本刚, 张昭, 于俊林. 珍稀濒危药用植物黄檗野生种群遗传多样性的AFLP分析. 生物多样性, 2006, 14(6): 488-497.
[8]闫志峰, 张本刚, 张昭. 迁地保护黄檗群体的遗传多样性评价. 中国中药杂志, 2008, 33(10): 1121-1125.
[9]Ewa Zietkiewicz, Antoni Rafalski, Damian Labuda. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 1994, 20(2): 176-183.
[10]Wolfe A D, Liston A. Contributions of PCR-based methods to plant systematics and evolutionary biology // Soltis D E, Soltis P S, Doyle J J, eds. Molecular Systematics of Plants II: DNA Sequencing. New York: Kluwer Academic Publishers, 1998: 43-86.
[11]Harris S A. RAPDs in systematics——a useful methodology? // Hollingsworth P M, Bateman R M, Gornall R J, eds. Molecular Systematics and Plant Evolution. London: Taylor and Francis, 1999: 221-228.
[12]Tsumura Y, Ohba K, Strauss S H. Diversity and inheritance of inter-simple sequence repeat polymorphisms in Douglas-fir (Pseudotsugamenziesii) and sugi (Cryptomeriajaponica). Theoretical and Applied Genetics, 1996, 92(1): 40-45.
[13]Martins M, Tenreiro R, Oliveira M M. Genetic relatedness of Portuguese almond cultivars assessed by RAPD and ISSR markers. Plant Cell Reports, 2003, 22(1): 71-78.
[14]Panda S, Martin J, Aguinagalde I. Chloroplast and nuclear DNA studies in a few members of theBrassicaoleraceaL. group using PCR-RFLP and ISSB-PCR markers: a population genetic analysis. Theoretical and Applied Genetics, 2003, 106(6): 1122-1128.
[15]邹喻苹, 葛颂, 王晓东. 系统与进化植物学中的分子标记. 北京: 科学出版社, 2001.
[16]Lynch M,Miligan B G. Analysis of population genetic structure with RAPD markers.Molecular Ecology,1994,3:91-99.
[17]张杰, 吴迪, 汪春蕾, 屈红军, 邹学忠, 杨传平. 应用ISSR-PCR分析蒙古栎种群的遗传多样性. 生物多样性, 2007, 15(3): 292-299.
[18]祁建民, 周东新, 吴为人, 林荔辉, 吴建梅, 方平平. 用ISSR标记检测黄麻野生种与栽培种遗传多样性. 应用生态学报, 2003, 14(9): 1473-1477.
Analysis of genetic diversity in wild populations ofPhellodendronamurenseRupr. in Jilin Province using inter-simple sequence repeat
LI Shaochen, LI Fengming*, ZHANG Limin, REN Jun, LIN Yumei
JilinAcademyofForestryScience,Changchun130033,China
PhellodendronamurenseRupr. is a valuable timber species and a key species in the plant communities of Changbai Mountain in Jilin province; however, due to over-harvesting and utilization, its quantity and quality have clearly declined.. Therefore, based on its distribution on Changbai Mountain in Jilin province, 10 representative populations ofP.amurensewere selected, and their genetic diversity was investigated using the Inter-Simple Sequence Repeat (ISSR) technique. Using the ISSR data, the Nei and Shannon indices were determined to estimate the genetic variation of the 10 populations. This study aims to provide foundational data for preservation and protection ofP.amurense. Using 60 ISSR primers to screen the 10 representative populations ofP.amurense, 99 bands were amplified; 54 bands were polymorphic (54.5%). The polymorphism of the 10 populations ranged from 18.52% to 37.96%; the Hunchun population showed the greatest polymorphism, at 37.96%, the Lushuihe population showed the least, at 18.52%, and the average was 26.02%. The Shannon index ranged from 0.1103 to 0.1949, with an average of 0.1522. The Nei index ranged from 0.0759 to 0.1327, with an average of 0.1043. According to Nei′s method calculation of Phellodendron amurense 10 population genetic diversity is the DST = 0.1586, the differentiation index (GST = 0.6183, gene flow coefficient nm is 0.3086... The total genetic variation of 61.83% of the variation existed among populations, and the variation within population was only 38.17%..A cluster map of the unweighted pair-group method with arithmetic means(UPGMA) relationships, using the ISSRs as molecular markers, was constructed. The 10 populations could be divided into two groups: 1) Songjianghe, Lushuihe, Wangou, Ji-an, and Huinan provinces, and 2) Baishi Shan, Wangqing, Antu, Yanji, and Hunchun provinces. Based on the genetic structure ofP.amurense, protection measures were put forward, including guidance to create a medicinal or timber forestry. The local plant resources background investigation and summary of resources (including compartments, sub-compartments, each plant′s age, height, diameter, height under crown and crown). To select the local dominant groups to carry on in situ conservation. Ex situ conservation strategies should be used to increase the number of individuals; in the Baishan area, the Songjianghe, Lushuihe provinces should be selected, in the Tonghua Area,the Ji-an provinces should be selected,in the Yanbian area, the Wangqing and Baishi Shan provinces should be selected. Artificial propagation ofP.amurenseshould be used to promote the gradual recovery of populations, and resource manual registration should be established.
P.amurense; genetic diversity; ISSRs (Inter-Simple Sequence Repeats)
10.5846/stxb201411142255
国家“十二五”科技支撑计划项目资助(2012BAD22B04)
2014-11-14; 网络出版日期:2015-10-30
Corresponding author.E-mail: lkylfm@126.com
李绍臣, 李凤明, 张立民, 任军, 林玉梅.吉林省天然黄檗种群遗传多样性ISSR分析.生态学报,2016,36(13):4006-4012.
Li S C, Li F M, Zhang L M, Ren J, Lin Y M.Analysis of genetic diversity in wild populations ofPhellodendronamurenseRupr. in Jilin Province using inter-simple sequence repeat.Acta Ecologica Sinica,2016,36(13):4006-4012.
猜你喜欢
蜜蜂杂志(2022年8期)2022-09-14 14:29:36
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
系统工程学报(2021年4期)2021-12-21 06:21:08
传奇·传记文学选刊(2020年12期)2020-12-23 06:56:35
新世纪智能(高一语文)(2020年12期)2020-06-01 08:14:20
西南农业学报(2016年6期)2016-04-16 05:12:47
法医学杂志(2015年4期)2016-01-06 12:36:36
中国医药导报(2015年27期)2015-02-28 22:08:01
河南医学研究(2014年7期)2014-02-27 14:53:42
中国林副特产(2013年4期)2013-08-07 10:15:48
