
缺钾油菜叶片光合速率下降的主导因子及其机理
2016-08-24 10:33陆志峰鲁剑巍李小坤丛日环潘勇辉李凯旭
植物营养与肥料学报 2016年1期
陆志峰, 任 涛, 鲁剑巍, 李小坤, 丛日环, 潘勇辉, 李凯旭
(华中农业大学资源与环境学院,农业部长江中下游耕地保育重点实验室,湖北武汉 430070)
缺钾油菜叶片光合速率下降的主导因子及其机理
陆志峰, 任 涛, 鲁剑巍*, 李小坤, 丛日环, 潘勇辉, 李凯旭
(华中农业大学资源与环境学院,农业部长江中下游耕地保育重点实验室,湖北武汉 430070)
【目的】钾是油菜(Brassica napusL.)生长发育所必需的矿质元素,缺钾胁迫可导致油菜叶片光合功能的衰退,最终影响籽粒产量。本文通过比较田间条件下蕾薹期油菜叶片(长柄叶、 短柄叶和无柄叶)对缺钾胁迫的反应,探讨缺钾导致叶片光合速率下降的主导因子及其作用机理,为合理施用钾肥提高油菜产量提供理论依据。【方法】采用大田试验,研究不施钾(-K)和施钾(+K)条件下华油杂9号和中双9号单株重,蕾薹期不同类型叶片单叶重、 叶面积、K含量和叶绿素含量。利用Li-6400XT便携式光合测定系统测定各类叶片的光合速率(Pn)和气孔导度(gs)等气体交换参数,以及PSⅡ最大光化学量子效率(Fv/Fm)和实际光化学量子效率(ΦPSⅡ)等叶绿素荧光参数。并利用胞间CO2浓度(Ci)和气孔限制值(Ls)的变化,分析叶片光合作用的限制因子。【结果】1) 缺钾胁迫使成熟期华油杂9号和中双9号籽粒产量分别下降13.9%和27.2%。2)缺钾显著抑制了蕾薹期中双9号的生长,单株重下降了12.4%; 长柄叶的干重比正常供钾处理降低了19.2%,而对华油杂9号无影响。3) 缺钾主要影响蕾薹期长柄叶的光合生理特性,对短柄叶和无柄叶无影响。缺钾条件下,油菜长柄叶钾含量和叶绿素含量明显降低,其Pn也显著低于正常供钾处理。但缺钾对不同品种Pn的限制原因并不同,缺钾胁迫导致华油杂9号Ci显著降低,Ls增加了16.6%,即气孔因素是华油杂9号Pn下降的主要原因; 而中双9号Ci显著增加,Ls却下降了14.0%,Pn的下降是由非气孔因素引起的。4) 缺钾胁迫下中双9号长柄叶的Fv/Fm、ΦPSⅡ、 qP和ETR均显著降低,说明PSⅡ反应中心受损,NPQ显著上升,间接表明缺钾条件下中双9号Pn下降的原因与非气孔因素有关; 而华油杂9号各荧光参数均未受缺钾胁迫的影响。油菜收获期籽粒产量与各叶片钾素含量以及长柄叶气体交换和叶绿素荧光参数关系密切。【结论】缺钾胁迫导致蕾薹期油菜长柄叶光合功能加速衰退,影响收获期籽粒产量。中双9号长柄叶光合性能受缺钾胁迫的影响大于华油杂9号,这与品种自身对缺钾胁迫的耐受能力有关。
油菜; 叶片; 蕾薹期; 缺钾胁迫; 光合气体交换; 叶绿素荧光

钾素作为植物生长发育所必须的矿质元素,在维持细胞渗透压[4],调节气孔运动[5],保障酶活性、 增加光合作用[6],促进体内糖分的运输、 提高产量上发挥着重要作用[7]; 其中光合器官光合速率的降低是缺钾导致作物减产的重要原因[8]。蕾薹期油菜同时具备长柄叶、 短柄叶和无柄叶三类叶片光合器官,当根系吸收的钾素无法满足植株需求时,便会出现钾素从长柄叶向无柄叶和短柄叶快速转移,继而伴随着叶片失绿黄化焦枯,光合功能衰退,甚至萎缩死亡的现象。各类叶片生长发育特性的不同直接决定了其对缺钾胁迫的响应差异,弄清各叶片对缺钾胁迫的响应表现,尤其是气体交换和叶绿素荧光特性上的异同,有助于系统性分析缺钾胁迫对蕾薹期油菜光合性能的影响。
钾对植物光合作用的影响主要包括气孔因素和非气孔因素。彭海欢等研究发现缺钾条件下,水稻叶片净光合速率随着气孔导度的降低而下降,认为气孔因素是限制光合速率的主要原因[9]; 孙骏威等的研究结果表明,缺钾胁迫加剧了光合速率的下降,但对气孔导度没有影响,而胞间CO2浓度显著升高,暗示光合速率下降是由非气孔因素引起的[10]。同样,棉花[11]、 玉米[12]和小麦[13]等作物在缺钾条件下,光合速率和气孔导度均下降,但是缺钾导致光合速率下降的原因却不尽相同。此外,缺钾使水稻叶片光合机构受损,光合电子传递能力降低,发生光抑制[9,14]。因此,本研究立足于田间试验,以华油杂9号和中双9号为材料,通过测定三类叶片的气体交换和叶绿素荧光参数,探讨缺钾导致油菜叶片光合速率下降的主导因子及其作用机理,以期为改善油菜光合作用、 提高产量提供理论依据。
1 材料与方法
1.1试验点概况
试验于2013年9月至2014年5月在湖北省武穴市梅川镇(30°06′N,115°36′E)进行。试验田土壤为花岗片麻岩母质发育的红壤性水稻土,其理化性状为:pH5.3,有机质30.5g/kg,全氮1.7g/kg,全钾18.6g/kg,速效氮(碱解氮)123.4mg/kg,速效钾32.5mg/kg,速效磷5.7mg/kg。试验点前茬作物为水稻。
1.2试验设计
试验采用品种和钾肥两因素试验设计,品种包括华油杂9号和中双9号,钾肥处理包括不施钾(-K)和施钾(+K)两个处理,施钾处理钾肥用量为120kgK2O/hm2。每个处理3次重复,完全随机区组设计,小区面积为20m2。
各处理氮磷硼肥用量均为N180kg/hm2、P2O590kg/hm2、 硼砂15kg/hm2。供试肥料品种分别为尿素(含N46%)、 过磷酸钙(含P2O512%)、 氯化钾(含K2O60%)和硼砂(含B11%)。氮肥分3次施用,分别是基肥60%、 越冬肥和薹肥各占20%,其他肥料均一次性施用。油菜于2013年9月27日育苗,11月1日移栽,密度为11.25×104plants/hm2。
1.3测定项目与方法
1.3.1 土壤样品的采集与分析土壤基础样品均为前茬水稻收获后、 油菜基肥施用前采集。以整个试验田块为采样单元,在试验田块内以“S”形均匀布点15个,取0—20cm耕作层土壤,实验室风干磨细过 20 目和100目筛,供理化分析用。土壤基本理化性质按常规方法进行测定[15]。土壤pH按水土比2.5 ∶1,pH计测定; 有机质采用重铬酸钾容量法; 全氮采用半微量开氏法; 全钾采用NaOH熔融—火焰光度计法; 速效氮采用碱解扩散法; 速效钾采用1mol/LNH4OAc浸提—火焰光度计法; 速效磷采用0.5mol/LNaHCO3浸提—钼锑抗比色法测定。
1.3.3 气孔限制值气孔限制值(Ls)参照Berry和Downton[16]的方法计算:
(1)
式中,Ca为设定CO2浓度。
另外,许大全[17]介绍了辨别叶片Pn降低的主要原因的两个可靠判据,即Ci和Ls的变化方向。Ci降低和Ls升高表明气孔导度降低是主要原因; 而Ci增加和Ls降低则表明主要原因是非气孔因素。
1.3.4 叶绿素荧光参数利用配备好荧光叶室的Li-6400XT测定标记叶片的荧光参数。测定前用暗适应夹子处理叶片30min后测定初始荧光(F0)和最大荧光(Fm),再将样品置于活化光 [1200μmol/(m2·s)]下照射0.5h,在数据稳定后,得到稳态荧光(Fs),接着用饱和光照射,得到光适应下的最大荧光(Fm′),然后关闭活化光,再照射远红光测量光适应下的F0′。PSII最大光化学量子效率(Fv/Fm)、PSII实际光化学效率(ΦPSII)、 天线色素转换效率(Fv′/Fm′)和PSII的电子传递速率(ETR)均由仪器计算给出。光化学淬灭系数(qP)和非光化学淬灭系数(NPQ)按照Kramer等[18]的公式计算:
(2)
(3)
实产统计在收获期每个小区选取8株有代表性的植株进行籽粒产量的测定。
1.4数据分析方法
试验数据均采用MicrosoftExcel2013软件进行计算处理,利用SPSS18.0软件进行统计分析,采用OriginPro8.5软件作图。
2 结果与分析
2.1缺钾对油菜籽粒产量和蕾薹期干物质积累的影响
不同品种油菜对钾肥的反应不同(图1),华油杂9号籽粒产量高于中双9号,缺钾显著降低了两个品种油菜的籽粒产量,尤其是中双9号,与施钾处理相比下降了27.2%。缺钾胁迫导致蕾薹期中双9号的单株干重比施钾处理降低了12.4%,而华油杂9号则对钾肥的反应不敏感,施钾和不施钾处理两者干重并无显著性差异。

图1 不同钾肥处理油菜籽粒产量和蕾薹期单株干重Fig.1 Seed yield and dry matter content in different K treatments at stem elongation stage
[注(Note):K—钾肥处理Ktreatment;C—品种Cultivar. 方柱上不同小写字母表示同一品种不同钾处理间差异达5%显著水平Differentsmalllettersabovethebarsmeansignificantdifferenceat5%levelbetweentwoKtreatmentswithinsamecultivar; *和**分别表示钾肥处理、品种和两者交互作用对籽粒产量和单株干重的影响达5%和1%显著水平,ns表示影响不显著IndicatethatseedyieldanddrymattercontentaresignificantlyinfluencedbyKtreatment,cultivarandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.]
2.2缺钾对油菜叶片形态生理的影响
对不同品种和钾肥处理油菜蕾薹期叶片形态生理指标的分析(表1)可以看出,中双9号单个叶片干重、 叶面积以及叶绿素a/b明显高于华油杂9号,叶绿素含量却差别不大。缺钾对华油杂9号叶片干重、 叶面积均无影响,但显著降低了无柄叶和长柄叶的叶绿素含量和叶绿素a/b; 缺钾显著降低了中双9号长柄叶的叶片干重、 叶绿素含量和叶绿素a/b,但对叶面积的影响不显著。
两个品种不同叶片K含量对缺钾胁迫的反应存在明显差异(图2)。华油杂9号各叶片的钾含量均高于中双9号,缺钾使中双9号长柄叶和短柄叶的K含量分别下降了59.6%和39.6%,对无柄叶的影响较小。缺钾也同样降低了华油杂9号长柄叶和短柄叶的K含量,但降幅小于中双9号。在施钾和不施钾条件下,华油杂9号三种叶片K含量差异显著; 在施钾条件下,中双9号短柄叶和长柄叶K含量显著大于无柄叶,而在不施钾时,无柄叶和短柄叶的钾含量却显著大于长柄叶。
2.3缺钾对油菜叶片气体交换参数的影响
从蕾薹期不同品种和钾肥处理下油菜叶片气体交换参数可以看出,两个品种间Pn、 Ci和Tr明显不同,而Gs并无显著性差异,华油杂9号的Pn和Tr高于中双9号,而Ci则较低(表2)。缺钾条件下,华油杂9号长柄叶的Pn、 Gs、 Ci和Tr分别下降了13.0%、 20.9%、 7.7%和14.1%; 中双9号的Pn、 Gs和Tr在缺钾时的下降幅度大于华油杂9号,而Ci却显著增加; 缺钾胁迫对短柄叶和无柄叶的气体交换参数无显著影响。
2.4缺钾对油菜叶片气孔限制值的影响
两个品种油菜各类叶片的气孔限制值如图3所示,缺钾不影响无柄叶和短柄叶的Ls,但使华油杂9号长柄叶的Ls增加了16.6%; 相反,使中双9号长柄叶的Ls降低了14.0%。因此,华油杂9号长柄叶Ci降低(表3),而Ls显著升高,说明气孔因素是其Pn下降的主要原因; 中双9号长柄叶Ci显著降低,而Ls升高,说明非气孔因素是引起Pn下降的主导因素。
2.5缺钾对油菜叶片叶绿素荧光参数的影响
从不同品种和钾肥处理油菜蕾薹期叶片叶绿素荧光特性的分析可以看出,缺钾胁迫对华油杂9号各类型叶片以及中双9号无柄叶和短柄叶的PSⅡ最大光化学量子效率(Fv/Fm,反应PSⅡ利用光能的能力,也作为光抑制的重要指标[20])无影响,但显著降低了中双9号长柄叶的Fv/Fm(表3)。实际光化学量子效率(ΦPSⅡ,反映PSⅡ反应中心在部分关闭情况下的实际原初光能捕获效率[20])在各品种不同钾处理间的反映和Fv/Fm一致,中双9号长柄叶的ΦPSⅡ在缺钾时下降了23.0%。缺钾条件下,PSⅡ电子传递速率(ETR,其值越低,表明光抑制程度越重[21])的变化也与上述两个参数一致,中双9号长柄叶的ETR比施钾处理下降了26.1%。以上结果说明蕾薹期油菜长柄叶利用光能的能力下降,光抑制程度增加,缺钾进一步加剧了光的抑制,且不同品种对缺钾的耐性有差异。

表1 蕾薹期钾肥处理油菜叶片形态生理特征Table 1 Leaf morphological and physiological characteristics in different K treatments at stem elongation stage
注(Note): 数据后不同字母表示同种叶片不同钾处理间差异达5%显著水平Valuesfollowedbydifferentlettersaresignificantdifferenceat5%levelbetweenKtreatmentswithinsameleaftype, *、 **和ns分别表示差异达5%、 1%显著水平和差异不显著Meansignificantdifferencesat5%, 1%levelandnosignificantdifference,respectively.

图2 蕾薹期不同钾肥处理油菜叶片钾含量Fig.2 Leaf K content in different K treatments at stem elongation stage
[注(Note):K—钾肥处理Ktreatment;L—叶片Leaf. 方柱上不同小写字母表示同种叶片不同钾处理间差异达5%显著水平Differentsmalllettersabovethebarsmeansignificantdifferenceat5%levelbetweentwoKtreatmentswithinsameleaftype; *和**分别表示钾肥处理、叶片和两者交互作用对气孔限制值的影响达5%和1%显著水平,ns表示影响不显著IndicatethatleafstomatallimitationvalueissignificantlyinfluencedbyKapplicationrate,leafandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.]
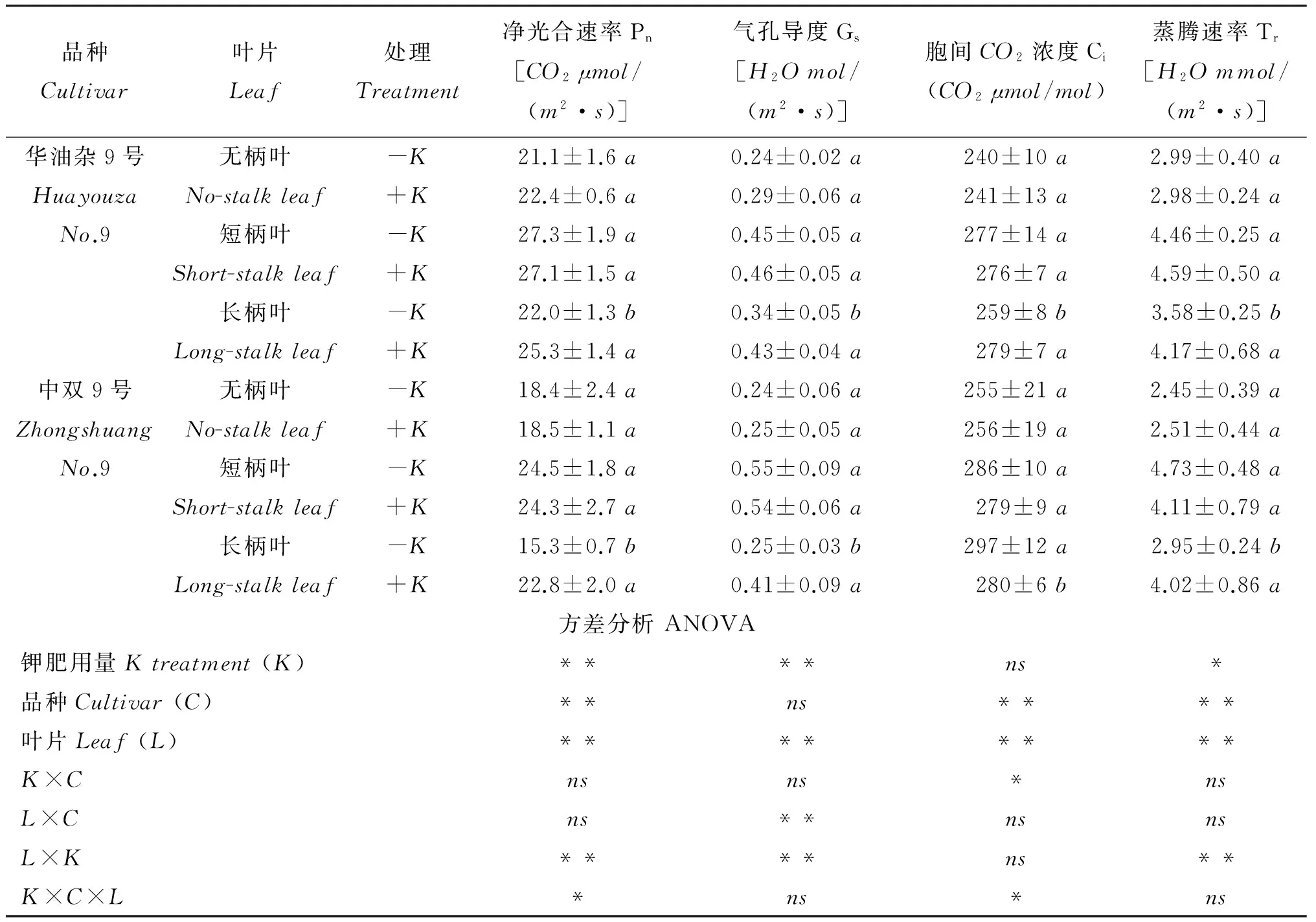
表2 蕾薹期不同钾肥处理油菜叶片气体交换参数Table 2 Leaf gas exchange parameters in different K treatments at stem elongation stage
注(Note):K—钾肥用量Kapplicationrate;C—品种Cultivar;L—叶片Leaf. 不同字母表示同种叶片不同钾处理间差异达5%显著水平Differentlettersmeansignificantdifferenceat5%levelbetweenKtreatmentswithinsameleaftype. *和**分别表示钾肥用量、 叶片和两者交互作用对叶片钾含量的影响达5%和1%显著水平,ns表示影响不显著IndicatethatleafKcontentissignificantlyinfluencedbyKapplicationrate,leafandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.
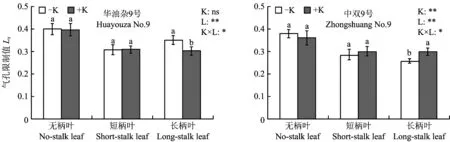
图3 蕾薹期不同钾肥处理油菜叶片气孔限制值Fig.3 Limitations due to stomatal resistance in different K treatments at stem elongation stage
[注(Note):K—钾肥处理Ktreatment;L—叶片Leaf. 方柱上不同小写字母表示同种叶片不同钾处理间差异达5%显著水平Differentsmalllettersabovethebarsmeansignificantdifferenceat5%levelbetweentwoKtreatmentswithinsameleaftype; *和**分别表示钾肥处理、叶片和两者交互作用对气孔限制值的影响达5%和1%显著水平,ns表示影响不显著IndicatethatleafstomatallimitationvalueissignificantlyinfluencedbyKapplicationrate,leafandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.]
缺钾显著影响了中双9号长柄叶的光化学淬灭系数(qP,反映了PSⅡ原初电子受体QA的氧化还原状态[21]),但并未影响其他类型叶片。华油杂9号施钾和不施钾处理间各叶片的qP也未受到影响。缺钾胁迫使中双9号长柄叶的非光化学淬灭系数(NPQ,指PSⅡ天线色素吸收但不能用于光合电子传递而以热能的形式耗散掉的光能部分[22])增加了18.3%,而未对华油杂9号造成影响。以上结果说明蕾薹期油菜长柄叶PSⅡ反应中心的开放程度较低,QA的还原状态增加,电子传递活性较低,缺钾胁迫进一步加剧了反应中心的关闭,降低了电子的传递,却通过增加热耗散来减轻光抑制的损伤。

表3 蕾薹期不同钾肥处理油菜叶片叶绿素荧光参数Table 3 Leaf chlorophyll fluorescence parameters in different K treatments at stem elongation stage
注(Note): Fv/Fm—PSⅡ最大光化学量子效率MaximumquantumefficiencyofPSⅡphotochemistry;ΦPSⅡ—PSⅡ实际光化学量子效率ActualphotochemicalefficiencyofPSⅡ;ETR—电子传递速率Electrontransportrate; qP—光化学淬灭系数Photochemicalquenchingcoefficient;NPQ—非光化学淬灭系数Non-photochemicalquenchingcoefficient. 数值后不同字母表示同种叶片不同钾处理间差异达5%显著水平Valuesfollowedbydifferentlettersmeansignificantdifferenceat5%levelbetweenKtreatmentswithinsameleaftype. *和**分别表示钾肥用量、 叶片和两者交互作用对叶片钾含量的影响达5%和1%显著水平,ns表示影响不显著IndicatethatleafKcontentissignificantlyinfluencedbyKapplicationrate,leafandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.
2.6籽粒产量及蕾薹期干物质积累与叶片光合生理参数的相关性
对比分析收获期单株籽粒产量与蕾薹期叶片光合生理参数的相关性(表4),可以发现收获期籽粒产量与蕾薹期各叶片的钾素含量显著相关。籽粒产量与长柄叶的叶绿素含量、 净光合速率以及叶绿素荧光参数(除Fv/Fm外)显著相关,而与无柄叶和短柄叶的叶绿素含量和光合气体交换以及叶绿素荧光参数(除qP外)无显著相关性。

表4 籽粒产量与不同类型叶片光合生理参数的相关性(r)Table 4 Correlations between seed yield and photosynthetic physiological characteristics of different leaves
注(Note):K—叶片钾含量LeafKcontent;Chl. —叶绿素含量Leafchlorophyllconcentration; Pn—净光合速率Netphotosyntheticrate; Gs—气孔导度Stomatalconductance; Ci—胞间CO2浓度IntercellularCO2concentration; Fv/Fm—PSⅡ最大光化学量子效率MaximumquantumefficiencyofPSⅡphotochemistry;ΦPSⅡ—PSⅡ实际光化学量子效率ActualphotochemicalefficiencyofPSⅡ;ETR—电子传递速率Electrontransportrate; qP—光化学淬灭系数Photochemicalquenchingcoefficient;NPQ—非光化学淬灭系数Non-photochemicalquenchingcoefficient. *和**分别表示钾肥用量、 叶片和两者交互作用对叶片钾含量的影响达5%和1%显著水平,ns表示影响不显著IndicatethatleafKcontentissignificantlyinfluencedbyKapplicationrate,leafandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.
3 讨论
3.1缺钾对蕾薹期油菜叶片光合性能的影响
本研究结果表明,缺钾主要影响蕾薹期油菜长柄叶的光合功能和叶绿素荧光特性,而未对短柄叶和无柄叶产生限制,这与油菜植株生长发育规律和叶片钾素含量有关。3类叶片中无柄叶的发生时间较晚,其光合速率和叶绿素含量处于上升期[23-24],短柄叶的发生早于无柄叶,处于光合功能的稳定期,叶片中钾素含量较高,光合功能相对较强。而长柄叶的发生发育时间最早,蕾薹期时处于植株最下部,当出现钾素亏缺时,长柄叶
的钾素快速向短柄叶和无柄叶转移[25],含量显著下降,加快了叶绿素的分解。而叶片叶绿素含量是反映作物衰老状况和光合能力的一个重要指标[26],可见进入蕾薹期后长柄叶开始衰老,光合能力下降,缺钾加速了长柄叶的衰退。长柄叶衰退过程中约有90%的钾素输向其他器官[25],对保证缺钾条件下上部叶片的光合功能有一定益处。
此外,长柄叶叶绿素含量和光合参数(Ci除外)与叶片钾含量显著相关,缺钾是导致光合能力下降的原因。缺钾条件下,Gs和叶绿素荧光参数的变化与叶片叶绿素含量无关(表5),这与Lu等[27]的研究结果相一致,即在小麦旗叶衰老的过程中,叶绿素含量的适当下降并不会导致叶绿素荧光参数的剧烈变化。

表5 长柄叶钾含量和叶绿素含量与光合参数的相关性(r)Table 5 Correlations between K, chlorophyll content and photosynthetic parameters of Long-stalk leaf (r value)
注(Note): Pn—净光合速率Netphotosyntheticrate;K—叶片钾含量LeafKcontent; Gs—气孔导度Stomotalconductance; Ci—胞间CO2浓度IntercellularCO2concentration; Tr—蒸腾速率Transpirationrate; *和**分别表示钾肥用量、 叶片和两者交互作用对叶片钾含量的影响达5%和1%显著水平,ns表示影响不显著IndicatethatleafKcontentissignificantlyinfluencedbyKapplicationrate,leafandtheirinteractionat5%and1%levels,respectively,andnsmeansnotsignificant.
3.2缺钾引起油菜长柄叶光合速率下降的主导因素及其作用机制
本研究中,华油杂9号长柄叶Pn下降的主导因素来自气孔,气孔导度的下降导致光合作用能够利用的CO2减少,光合速率下降; 同时测定的叶绿素荧光参数差别不大,说明光能的吸收和电子传递等都正常。而非气孔限制是中双9号长柄叶Pn下降的主导因素,其中CO2从细胞间隙向叶绿体传递阻力的增加以及碳同化过程CO2的利用减少(包括碳同化酶活性和能力合成等过程)可能是胞间CO2升高,光合速率下降的原因[23]。缺钾胁迫使中双9号长柄叶Fv/Fm和ΦPSⅡ下降,表明缺钾对PSⅡ造成了伤害,光能的利用能力下降。与此同时ETR显著降低,说明源于光合磷酸化和电子传递过程相偶联所生成的ATP和NADPH减少,碳同化过程也正因此而受到限制,这与孙骏威等[14]在水稻上的研究结果一致。缺钾显著降低了叶片的qP,说明较大部分反应中心处于关闭状态,QA处于较高的还原态,无法快速有效的传递H2O光解产生的电子,这也是ETR降低的原因。PSⅡ反应中心功能的下降,往往会导致光合机构吸收的光能进入光化学过程的量显著减小,使植物产生光抑制[10,14]。在缺钾条件下,中双9号长柄叶通过增加热耗散(NPQ),在一定程度上减轻了光合机构所遭受的光破坏。
缺钾引起两个品种长柄叶光合速率下降主导因素的不同,和品种自身对缺钾胁迫耐受能力的差异有关。华油杂9号比中双9号具有更强的吸钾能力,在缺钾时体内还能够保持较高的钾含量,使各光合参数所受的影响小。当钾素亏缺时,中双9号长柄叶钾素快速转移,面临着严重缺钾威胁的长柄叶加快进入早衰阶段[25]。相反,由于华油杂9号吸收了更多的钾,长柄叶面临的缺钾威胁较轻,其光合功能较中双9号更加稳定。Bednarz等[28]研究认为,气孔限制是轻度缺钾条件下棉花叶片光合速率下降的主要原因,但随着缺钾胁迫的加重,非气孔限制开始占主导地位。因此,此种差异的出现可能与叶片K含量有关。李向东等[29]的研究指出,花生叶片衰老过程中,气孔限制不断降低,而非气孔限制逐渐增强,说明光合作用的限制因素还可能与叶片的衰老程度相关。此外,叶片在缺钾及衰老过程伴随着Rubisco酶活性的降低[30],气孔导度和叶肉导度的下降[23],叶绿体结构的破坏[29],光系统活性衰退,活性氧清楚系统功能的减弱[10]等,都可能导致光合作用限制因素的转变。本研究中,长柄叶钾含量和衰老程度对光合作用的影响表现趋同,均和油菜对缺钾的耐受能力有关。
4 结论
1)缺钾主要抑制蕾薹期油菜下部长柄叶的光合能力,对中部短柄叶和上部无柄叶影响不显著。缺钾导致长柄叶光合性能的加速衰退与油菜产量的降低关系密切。
2)缺钾条件下,华油杂9号和中双9号长柄叶光合速率下降分别受气孔因素和非气孔因素的主导。此种差异的出现可能和叶片钾含量以及衰老程度有关,而这又都取决于品种自身对缺钾胁迫耐受能力的不同,华油杂9号对缺钾胁迫的耐受能力大于中双9号。
[1]RenT,LuJW,LiH, et al.Potassium-fertilizermanagementinwinteroilseed-rapeproductioninChina[J].JournalofPlantNutritionandSoilScience, 2013, 176(3): 429-440.
[2]李银水, 鲁剑巍, 廖星, 等.钾肥用量对油菜产量及钾素利用效率的影响[J].中国油料作物学报, 2011, 33(2): 152-156.
LiYS,LuJW,LiaoX, et al.Effectofpotassiumapplicationrateonyieldandfertilizer-potassiumutilizationefficiencyinrapeseed[J].ChineseJournalofOilCropSciences, 2011, 33(2): 152-156.
[3]刘晓伟.冬油菜养分吸收规律及不同养分效率品种特征比较研究[D].武汉: 华中农业大学硕士学位论文, 2011.
LiuXW.Studyonnutrientabsorptionofoilseedrape(Brassica napusL.)andcharacteristicscompareindifferentnutrientefficiencytypes[D].Wuhan:MSThesis,HuazhongAgriculturalUniversity, 2011.
[4]PervezH,AshrafM,MakhdumMI.Influenceofpotassiumnutr-
itionongasexchangecharacteristicsandwaterrelationsincotton(Gossypium hirsutumL.) [J].Photosynthetica, 2001, 42: 251-255.
[5]LebaudyA,VavasseurA,HosyE, et al.Plantadaptationtofluc-
tuatingenvironmentandbiomassproductionarestronglydependentonguardcellpotassiumchannels[J].ProceedingsoftheNationalAcademyofSciences, 2008, 105(13): 5271-5276.
[6]HanQ.Height-relateddecreasesinmesophyllconductance,leafphotosynthesisandcompensatingadjustmentsassociatedwithleafnitrogenconcentrationsinPinus densiflora [J].TreePhysiology, 2011, 31: 976-984.
[7]PettigrewWT.Potassiuminfluencesonyieldandqualityproduct-
ionformaize,wheat,soybeanandcotton[J].Physiologiaplantarum, 2008, 133(4): 670-681.
[8]刘高洁, 逄焕成, 李玉义.长期施肥对潮土夏玉米生长发育和光合特性的影响[J].植物营养与肥料学报, 2010, 16(5): 1094-1099.
LiuGJ,PangHC,LiYY.Effectsoflong-termfertilizationongrowthandphotosynthesisofsummermaizeinfluvo-aquicsoil[J].PlantNutritionandFertilizerScience, 2010, 16(5): 1094-1099.
[9]彭海欢, 翁晓燕, 徐红霞, 等.缺钾胁迫对水稻光合特性及光合防御机制的影响[J].中国水稻科学, 2006, 20(6): 621-625.
PengHH,WengXY,XuHX, et al.Effectsofpotassiumdeficiencyonphotosynthesisandphoto-protectionmechanismsinriceplants[J].ChineseJournalofRiceScience, 2006, 20(6): 621-625.
[10]孙骏威, 李素芳, 付贤树, 等.低钾对水稻不同叶位叶片光合特性及抗氧化系统的影响[J].核农学报, 2006, 21(4): 404-408.
SunJW,LiSF,FuXS, et al.Effectsoflowpotassiumstressonphotosyntheticcharacteristicsandantioxidantsystemsindifferentpositionleavesofriceplants[J].JournalofNuclearAgriculturalSciences, 2006, 21(4): 404-408.
[11]WangN,HuaH,EgrinyaEnejiA, et al.Genotypicvariationsinphotosyntheticandphysiologicaladjustmenttopotassiumdeficiencyincotton(Gossypium hirsutum) [J].JournalofPhotochemistryandPhotobiologyB:Biology,B, 2012, 110: 1-8.
[12]陈爱珠, 杨杰文.钾素对甜玉米苗期光合特性的影响[J].中国农学通报, 2010, 26(9): 230-233.
ChenAZ,YangJW.Effectsofpotassiumonphotosyntheticcharactersofsweetcorninseedlingphasing[J].ChineseAgriculturalScienceBulletin, 2010, 26(9): 230-233.
[13]于振文, 张炜, 岳寿松, 等.钾营养对冬小麦光合作用和衰老的影响[J].作物学报, 1996, 22(3): 305-312.
YuZW,ZhangW,QiuSS, et al.Effectofpotassiumonphotosynthesisandsenescenceinwinterwheat[J].ActaAgronmicaSinica, 1996, 22(3): 305-312.
[14]孙骏威, 翁晓燕, 李峤, 等.缺钾对水稻不同品种光合和能量耗散的影响[J].植物营养与肥料学报, 2007, 13(4): 577-584.
SunJW,WenXY,LiQ, et al.Effectsofpotassium-deficiencyonphotosynthesisandenergydissipationindifferentricecultivars[J].PlantNutritionandFertilizerScience, 2007, 13(4): 577-584.
[15]鲍士旦.土壤农化分析(第3版)[M].北京: 中国农业出版社, 2000. 263-271.
BaoSD.Soilandagrochemistryanalysis(3rdEd) [M].Beijing:ChinaAgriculturalPress, 2000: 30-107, 270.
[16]BerryJA,DowntonWJS.Environmentalregulationofphotosy-
nthesis[J].Photosynthesis, 1982, 2: 263-343.
[17]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯, 1997, 33 (4): 241-244.
XuDQ.Someproblemsinstomatallimitationanalysisofphotosynthesis[J].PlantPhysiologyCommunications, 1997, 33(4): 241-244.
[18]KramerDM,JohnsonG,KⅡratsO, et al.NewfluorescenceparametersforthedeterminationofQAredoxstateandexcitationenergyfluxes[J].PhotosynthesisResearch, 2004, 79(2): 209-218.
[19]舒展, 张晓素, 陈娟, 等.叶绿素含量测定的简化[J].植物生理学通讯, 2010, 46(4): 399-402.
ShuZ,ZhangXS,ChenJ, et al.Thesimplificationofchlorophyllcontentmeasurement[J].PlantPhysiologyCommunications, 2010, 46(4): 399-402.
[20]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报, 1999, 16(4): 444-448.
ZhangSR.Adiscussiononchlorophyllfluorescencekineticsparametersandtheirsignificance[J].ChineseBulletionofBotany, 1999, 16(4): 444-448.
[21]王菲, 曹翠玲.磷水平对不同磷效率小麦叶绿素荧光参数的影响[J].植物营养与肥料学报, 2010, 16(3): 758-762.
WangF,CaoCL.Effectsofphosphoruslevelsonchlorophyllfluorescenceparametersofwheat(Triticum aestivumL.)withdifferentphosphorusefficiencies[J].PlantNutritionandFertilizerScience, 2010, 16(3): 758-762.
[22]BilgerW,BjörkmanO.Roleofthexanthophyllcycleinphotop-
rotectionelucidatedbymeasurementsoflight-inducedabsorbancechanges,fluorescenceandphotosynthesisinleavesofHederacanariensis[J].PhotosynthesisResearch, 1990, 25(3): 173-185.
[23]Battie-LaclauP,LaclauJP,BeriC, et al.PhotosyntheticandanatomicalresponsesofEucalyptus grandisleavestopotassiumandsodiumsupplyinafieldexperiment[J].Plant,CellandEnvironment, 2014, 37(1): 70-81.
[24]赵懿.油菜光合功能衰退研究[D].中国农业科学院, 2006.
ZhaoY.Studyonphotosyntheticfunctionaldeclineforoilseedrape(Brassica napusL.) [D].Chineseacademyofagriculturalsciences, 2006.
[25]冷锁虎, 单玉华, 李德权.油菜叶片衰老与NPK含量变化[J].中国油料作物学报, 2001, 23(1): 38-40.
LengSH,ShanYH,LiDQ.ChangesofNPKcontentsindecrepitleavesofrapeseed[J].ChineseJournalofOilCropSciences, 2001, 23(1): 28-40.
[26]OhSA,ParkJH,LeeGI, et al.IdentificationofthreegeneticlocicontrollingleafsenescenceinArabidopsisthaliana[J].ThePlantJournal, 1997, 12(3): 527-535.
[27]LuQT,LuCM,ZhangJH,KuangTY.Photosynthesisandchlorophyllafluorescenceduringflagleafsenescenceoffield-grownwheatplants[J].JournalofPlantPhysiology, 2002, 159(11): 1173-1178.
[28]BednarzCW,OosterhuisDM,EvansRD.Leafphotosynthesisandcarbonisotopediscriminationofcottoninresponsetopotassiumdeficiency[J].EnvironmentalandExperimentalBotany, 1998, 39(2): 131-139.
[29]李向东, 王晓云, 余松烈, 等.花生叶片衰老过程中光合性能及细胞微结构变化[J].中国农业科学, 2002, 35(4): 384-389.
LiXD,WangXY,YuSL, et al.Thechangesofphotosyntheticpropertiesandcellmicrostructureinpeanutleavesduringleafsenescence[J].ScientiaAgriculturaSinica, 2002, 35(4): 384-389.
[30]郑炳松, 程晓建, 蒋德安, 翁晓燕.钾元素对光合速率、Rub-
isco和RCA的影响[J].浙江林学院学报, 2002, 19(1): 104-108.
ZhengBS,ChenXJ,JiangDA,WengXY.EffectsofpotassiumonRubisco,RCAandphotosyntheticrateofplant[J].JournalZhejiangForestryCollege, 2002, 19(1): 104-108.
Mainfactorsandmechanismleadingtothedecreaseofphotosyntheticefficiencyofoilseedrapeexposuretopotassiumdeficiency
LUZhi-feng,RENTao,LUJian-wei*,LIXiao-kun,CONGRi-huan,PANYong-hui,LIKai-xu
[College of Resources and Environment, Huazhong Agricultural University/Key Laboratory of Arable Land Conservation (Middle and Lower Reaches of Yangtse River), Ministry of Agriculture, Wuhan 430070, China]
【Objectives】Potassium(K)isessentialforoilseedrape(Brassica napusL.),insufficientsupplywilldeclinetheleafphotosyntheticcapacity,aswellasseedyield.Thepresentstudywasconductedtocomparetheresponseofdifferentleaves(namelylong-stalkleaf,short-stalkleafandno-stalkleaf)toKdeficiencyatstemelongationstageunderfieldconditions.Additionally,weattemptedtofindoutthedominantlimitationsandrevealtheunderlyingmechanismsduetoKdeficiencywhichrestrainedtheleafphotosynthesis,andtoprovidetheoreticalbasisforthereasonableapplicationofpotashfertilizeronthepurposeofimprovingseedyield. 【Methods】AfieldexperimentwithnoKsupply(-K)andnormalKsupply(+K)wasconductedusingtworapeseedcultivars(HuayouzaNo.9andZhongshuangNo.9).Theplantdrymatterbiomass,leafdrymattercontent,leafarea,leafKconcentrationandchlorophyllcontentwereconcurrentlydetermined.Theleafgasexchangeparameters,includingnetphotosynthesis(Pn)andstomatalconductance(gs),andleafchlorophyllfluorescenceparameters,includingmaximumquantumyieldofPSⅡ(Fv/Fm)andeffectivequantumefficiencyofPSⅡ (ΦPSⅡ),thechangesofCiandstomatallimitations(Ls)weredetected.【Results】1)Potassiumdeficiencyleadedtoasignificantreductionofseedyield,inHuayouzaNo.9thedecreasewas13.9%andinZhongshuangNo.9was27.2%. 2)ThegrowthofZhongshuangNo.9wassignificantlysuppressedunderinsufficientpotassiumsupply,with12.4%and19.2%decreasesofplantdrymattercontentandleafdrymattercontentinlong-stalkleaf,respectively,whilethoseinHuayouzaNo.9wereunaffected. 3)Potassiumdeficiencymainlyaffectedthephotosyntheticphysiologicalcharacteristicsoflong-stalkleaf,butnotonthoseofshortandno-stalkleaves.UnderKdeficient,theKcontent,chlorophyllcontentandPnoflong-stalkleafweresharplydeclined.ThedominantfactorsinvolvedinPndeclineweredifferentbetweenthetwocultivars.PotassiumdeficiencyledtosharplydeclineofCibut16.6%increaseinLsinHuayouzaNo.9,indicatingthatPnwasprimarilyaffectedbystomatallimitations.Conversely,KdeficiencyledtothedramaticincreaseinCiand14.0%ofdecreaseinLsinZhongshuangNo.9,whichsuggestedthatPnwasprimarilylimitedbynon-stomatallimitations. 4)Moreover,KdeficiencycausedsharpdeclineintheleafFv/Fm,ΦPSⅡ, qPandETRofZhongshuangNo.9,butnotinthoseofHuayouzaNo.9,indicatingthatthePSⅡreactioncenterwassufferedofdamage.Meanwhile,theobviousincrementofNPQhintedthatnon-stomatallimitationsinducedthedeclineofPn. 【Conclusions】Thedeclineofphotosyntheticefficiencyinthelong-stalkleafofrapseedisexacerbatedunderKdeficiency,whichresultesinthedecreaseofseedyield.TheZhongshuangNo.9wasmoresensitivetoK-starvationthanHuayouzaNo.9,whichisassociatedwithitstolerancetoKdeficiency.
rapeseed;leaf;stemelongationstage;potassiumdefficiency;gasexchange;chlorophyllfluorescence
2014-08-28接受日期: 2015-02-04网络出版日期: 2015-07-02
国家油菜产业技术体系建设专项(CARS-13); 公益性行业(农业)科研专项(201203013); 华中农业大学博士研究生创新研究工程项目(2014bs17)资助。
陆志峰(1989—),男,浙江衢州人,博士研究生,主要从事现代施肥技术研究。E-mail:luzhifeng@webmail.hzau.edu.cn
E-mail:lunm@mail.hzau.edu.cn
S565.4;Q945.11
A
1008-505X(2016)01-0122-10
猜你喜欢
疯狂英语·新读写(2022年4期)2022-11-22
疯狂英语·读写版(2022年4期)2022-04-08
花火·慧阅读(2022年4期)2022-03-18
江苏农业科学(2022年4期)2022-03-11
干旱地区农业研究(2022年1期)2022-01-28
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
农家科技下旬刊(2018年8期)2018-10-16
农业科技与装备(2016年9期)2017-03-10
花样盛年(2016年6期)2016-06-07
