
嗜麦芽窄食单胞菌产氨基糖苷类修饰酶基因APH(3′′)-Ⅰ和AAC(2′)-Ⅰ的克隆与表达
2016-08-18 01:32关小珊关锐梨钟华敏邓秋连谢永强周珍文
国际检验医学杂志 2016年15期
关小珊,关锐梨,钟华敏,邓秋连,谢永强,周珍文
(广州市妇女儿童医疗中心儿童院区检验科 510120)
·论著·
嗜麦芽窄食单胞菌产氨基糖苷类修饰酶基因APH(3′′)-Ⅰ和AAC(2′)-Ⅰ的克隆与表达
关小珊,关锐梨,钟华敏,邓秋连,谢永强,周珍文△
(广州市妇女儿童医疗中心儿童院区检验科510120)
目的对嗜麦芽窄食单胞菌临床分离株gzch810(SM gzch810)携带的产氨基糖苷类修饰酶基因APH(3′′)-Ⅰ和AAC(2′)-Ⅰ进行扩增、测序及原核表达,为下一步功能试验的开展提供基础材料。方法提取SM gzch810基因组染色体,PCR扩增APH(3′′)-Ⅰ和AAC(2′)-Ⅰ全基因并克隆入pMD18-T载体进行核苷酸序列分析;将APH(3′′)-Ⅰ和AAC(2′)-Ⅰ基因克隆至表达载体pGEX-4T-1,转化大肠杆菌BL21,SDS-PAGE分析融合蛋白表达情况。结果以染色体DNA为模板,成功扩增800 bp APH(3′′)-Ⅰ基因和550 bp AAC(2′)-Ⅰ基因;序列比对分析显示其与相关报道序列核苷酸和氨基酸一致性分别达91%及95%;SM gzch810 APH(3′′)-Ⅰ和AAC(2′)-Ⅰ序列已登录GenBank(登录号:HQ315852和HQ315853);SDS-PAGE显示,融合基因表达的蛋白相对分子质量分别约为56×103和46×103。结论从SM gzch810中成功克隆及表达了APH(3′′)-Ⅰ和AAC(2′)-Ⅰ基因,为下一步检测上述两种重组体大肠杆菌对相应抗菌药物的耐药性及其功能评价做好准备。
嗜麦芽窄食单胞菌;产氨基糖苷类修饰酶基因;克隆;原核表达
嗜麦芽窄食单胞菌(SM)为医院内获得性感染的重要病原菌之一,中国CHINET监测网2013年度耐药监测数据表明,该菌占所有革兰阴性菌的3.96%,非发酵菌的10.82%[1];国外文献报道,SM引起肺部感染的归因死亡率高达20%~30%[2]。SM耐药机制复杂,对大多数抗菌药物耐药,是临床抗感染治疗中的一道难题,其中对β-内酰胺类、喹诺酮类药物的耐药机制已有较多研究,但对氨基糖苷类药物的耐药研究则较少。本文从临床儿童先天性心脏病患者血液分离菌株gzch810中PCR扩增出APH(3′′)-Ⅰ和AAC(2′)-Ⅰ基因并将其在大肠杆菌中进行表达,为下一步检测上述两种重组体大肠杆菌对氨基糖苷类抗菌药物的耐药性及其功能评价做好准备。
1 材料和方法
1.1材料
1.1.1SM gzch810分离株来源于2012年5月分离自本院CICU心脏病患者血液,患儿男性,日龄8 d。
1.1.2试剂与仪器pGEX-4T-1质粒载体购自法国马西亚公司;pMD18-T质粒载体,Ex-Taq DNA聚合酶,EcoRⅠ、XhoⅠ限制性内切酶,T4 DNA连接酶等购自Takara公司。IPTG、丙烯酰胺、Tris碱、甘氨酸等购自鼎国公司。德国Biometra公司PCR仪,英国Uvitec公司GAS7508-T20紫外凝胶成像分析系统,德国 Sorvall公司Micro 17R台式高速冷冻离心机。
1.2方法
1.2.1染色体DNA的提取及扩增根据GenBank提供的Smlt2336(YP001972129.1)和Smlt1669(YP001971501.1)序列,设计特异性引物两对:APH(3′′)-Ⅰ P1:5′-CGC GAA TTC ATG AGC GAA CCC TAC CTG AG-3′,APH(3′′)-Ⅰ P2:5′-AAA CTC GAG TCA CTT CTC CGC CAG CGC CT-3′;AAC(2′)-Ⅰ P1:5′-CGC GAA TTC ATG GGC CTG GAC ACA CAT TCA-3′,AAC(2′)-Ⅰ P2:5′-AAA CTC GAG TCA GAC CGT TTC GCC CTG CA-3′,引物序列分别加入EcoRⅠ、XhoⅠ酶切位点。以试剂盒提取的染色体DNA为模板进行PCR扩增,反应条件为预变性94 ℃ 5 min后,94 ℃ 45 s、60 ℃ 45 s、72 ℃ 1.5 min,循环35次,最后72 ℃延伸15 min。
1.2.2pMD18-T-APH(3′′)-Ⅰ/AAC(2′)-Ⅰ的构建及序列分析PCR产物纯化后与pMD18-T载体16 ℃连接2 h,连接产物转化大肠杆菌DH5α。挑取经氨苄西林筛选的阳性菌落提取质粒并经EcoRⅠ和XhoⅠ双酶切鉴定;将鉴定含目的基因的质粒菌送Invitrogen公司进行测序并将测序结果进行BLAST比对分析。
1.2.3pGEX-4T-1-APH(3′′)-Ⅰ/AAC(2′)-Ⅰ的构建及表达纯化PCR产物与pGEX-4T-1载体经EcoRⅠ和XhoⅠ酶切后连接过夜,转化大肠杆菌BL21;挑取经氨苄筛选的阳性菌落提取质粒并进行双酶切鉴定。将含有重组质粒的BL21接种于含氨苄西林(100 μg/mL)的LB培养基,37 ℃培养过夜后挑单菌落于含氨苄西林LB液体培养基中振荡培养至OD600值为0.4左右,加 IPTG至终浓度0.5 mmol/L,30 ℃诱导表达5 h,8 000 r/min 4 ℃离心15 min,收集菌体,SDS-PAGE电泳鉴定。
2 结 果
2.1pMD18-T-APH(3′′)-Ⅰ/AAC(2′)-Ⅰ重组质粒的测序结果测序结果显示,SM gzch810 APH(3′′)-Ⅰ基因全长813 bp;预测的相对分子质量为30 113.43,等电点为5.85。其序列已登录GenBank,登录号为HQ315852,与SM K279a株氨基酸同源性达91%(图1)。SM gzch810 AAC(2′)-Ⅰ基因全长549 bp;预测的相对分子质量为2 0183.02,等电点为5.96。其序列已登录GenBank,登录号为HQ315853,与SM K279a株氨基酸同源性达95%(图2)。

图1 SM gzch810 APH(3′′)-Ⅰ与SM K279a株氨基酸同源性比较
2.2SDS-PAGE检测Smqnr蛋白的表达重组菌经IPTG诱导后可见GST和APH(3′′)-Ⅰ、AAC(2′)-Ⅰ的融合蛋白表达带,相对分子质量分别约为56×103和46×103,含空质粒菌经诱导后则在26×103位置上出现GST表达带(图3、4)。
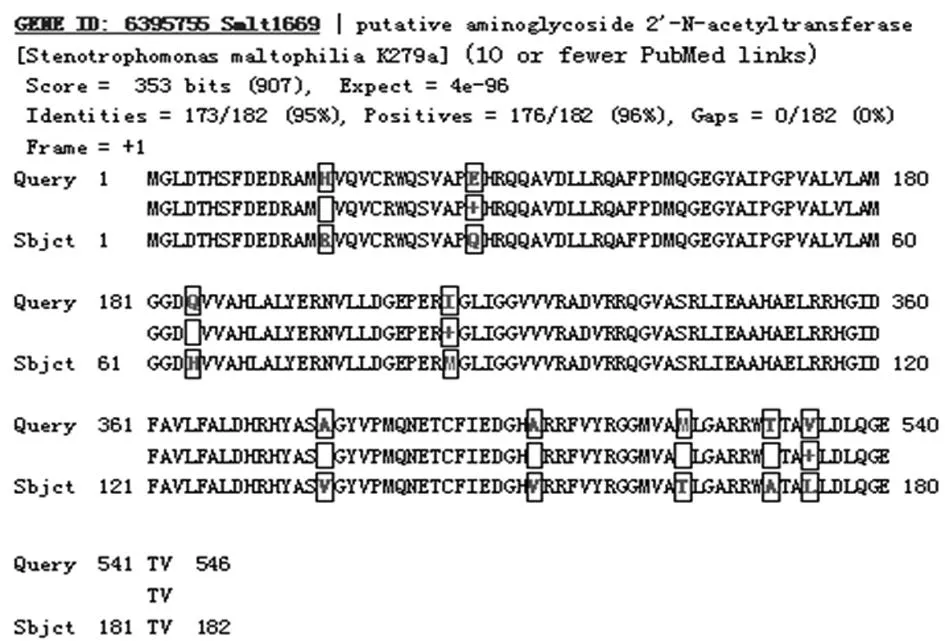
图2 SM gzch810 AAC(2′)-Ⅰ与SM K279a株氨基酸同源性比较

注:Lane1为预染蛋白marker;lane2为pGEX-4T-1-APH(3′′)-Ⅰ/BL21经诱导;Lane3为pGEX-4T-1/BL21经诱导。
图3重组蛋白表达产物SDS-PAGE电泳结果

注:Lane1为预染蛋白 marker;lane2为pGEX-4T-1-AAC(2′)-Ⅰ/BL21 经诱导;Lane3为pGEX-4T-1/BL21经诱导。
图4重组蛋白表达产物SDS-PAGE电泳结果
3 讨 论
SM是医院内感染的重要致病菌之一,对氨基糖苷类药物的耐药机制主要有:(1)膜屏障的改变或主动外排作用使药物的吸收减少;(2)核糖体结合位点的改变;(3)氨基糖苷类修饰酶(AMEs)对药物的修饰钝化作用,其中以AMEs的作用为主要原因。常见的AMEs 按功能可分为磷酸转移酶(APH)、乙酰转移酶(AAC) 和核苷转移酶(ANT) 3类,已在美国NCBI登录的AMEs基因有30余种[3],目前在临床分离革兰阴性杆菌中,最常见的有AAC(3)-Ⅰ、AAC(3)-Ⅱ、AAC(6′)-Ⅰb、AAC(6′)-Ⅱ、ANT(2′′)-Ⅰ、ANT(3′′)-Ⅰ[4-5]共6种,但在SM中除AAC(6′)-Ⅰz[6]、APH(3′)-Ⅱc[7]外,仅有少量检出ANT(2′′)-Ⅰ、AAC(6′)-Ⅱ 和AAC(3) -Ⅰ/Ⅱ[8-9]的报道。此类酶作用于氨基糖苷类抗菌药物特定的氨基或羟基,使抗菌药物发生钝化,降低或丧失对靶位核糖体的亲和力而不能进入下一阶段发挥抗菌作用。由AMEs所致的耐药性在不同的微生物或不同菌株之间有很大差异,取决于酶产生的数量、酶的催化活性和作用于何种抗菌药物等因素;在众多能产生足够活性而使细菌达到耐药标准的AMEs中,APHs能造成高水平耐药[10]。
APH是一种利用ATP作为第二底物,且能磷酸化所有氨基糖苷类抗菌药物的羟基酶。目前,临床上分离到的APH有7种,其中APH(3′′)-Ⅰ的作用底物只有链霉素,修饰其3′′位羟基。本次实验检出的APH(3′′)-Ⅰ基因经核苷酸及氨基酸序列分析,与GenBank上登记注册的Smlt 2336、Smlt 1923氨基酸同源性分别达到91%和89%,是国内首次从临床分离SM中检出APH(3′′)-Ⅰ基因,该基因序列已登录GenBank,登录号为HQ315852。SDS-PAGE结果显示,在IPTG的诱导下,重组表达质粒表达出了相对分子质量约56×103的融合蛋白,与核苷酸序列推导的蛋白质分子量大小相符。
AAC 有4 种同工酶:AAC(1)、AAC(3)、AAC(2′)和AAC(6′),主要以乙酰辅酶A作为乙酰基的供体,分别作用于氨基糖苷类抗菌药物的2-脱氧链霉胺环的1位和3 位、6-氨基己糖环的2′位和6′位。AAC(6′)和AAC(3)多数由转座子、整合子和质粒等可移动遗传元件介导,在抗菌药物的选择压力下能在微生物间迅速播散[11-12],而目前所发现的AAC(2′)则均由染色体编码。本次实验检出的AAC(2′)-Ⅰ基因经测序及同源性分析,与GenBank上登记注册的Smlt 1669氨基酸同源性达95%,与从结核分枝杆菌中检出的AAC(2′)-Ⅰc 同源性仅30%,推测其具种属特异性,不同菌属间不能传播。
本实验从临床分离株 gzch810 中经PCR检出两种AMEs基因APH(3′′)-Ⅰ和AAC(2′)-Ⅰ,并对其进行了克隆及表达,为下一步检测上述两种重组体大肠杆菌对相应抗菌药物的耐药性及其功能评价做好准备。
[1]胡付品,朱德妹,汪复,等.2013年中国CHINET细菌耐药性监测[J].中国感染与化疗杂志,2014,14(5):365-374.
[2]Falagas ME,Kastoris AC,Vouloumanou EK,et al.Attributable mortality of Stenotrophomonas maltophilia infections:a systematic review of the literature[J].Future Microbiol,2009,4(9):1103-1109.
[3]Ramirez MS,Tolmasky ME.Aminoglycoside modifying enzymes[J].Drug Resist Updat,2010,13(6):151-171.
[4]Ho PL,Leung LM,Chow KH,et al.Prevalence of aminoglycoside modifying enzyme and 16S ribosomal RNA methylase genes among aminoglycoside-resistant Escherichia coli isolates[J].J Microbiol Immunol Infect,2014,14:1-4.
[5]Haldorsen BC,Simonsen GS,Sundsfjord A,et al.Increased prevalence of aminoglycoside resistance in clinical isolates of Escherichia coli and Klebsiella spp.in Norway is associated with the acquisition of AAC(3)-Ⅱ and AAC(6′)-Ⅰb[J].Diagn Microbiol Infect Dis,2014,78(1):66-69.
[6]Li XZ,Zhang L,Mckay GA,et al.Role of the acetyltransferase AAC(6′)-Iz modifying enzyme in aminoglycoside resistance in Stenotrophomonas maltophilia[J].J Antimicrob Chemother,2003,51(4):803-811.
[7]Okazaki A,Avison MB.Aph(3′)-Ⅱc,an aminoglycoside resistance determinant from Stenotrophomonas maltophilia[J].Antimicrob Agents Chemother,2007,51(1):359-360.
[8]时东彦,付洁,李扬.嗜麦芽窄食单胞菌中氨基苷类修饰酶基因的研究[J].医药导报,2009,28(4):427-429.
[9]刘治敏,潘亚萍,吴桂华,等.189株嗜麦芽寡养单胞菌耐药性检测与氨基糖苷类修饰基因的分布[J].中华医院感染学杂志,2011,21(2):231-233.
[10]Vakulenko SB,Mobashery S.Versatility of aminoglycosides and prospects for their future[J].Clin Microbiol Rev,2003,16(3):430-450.
[11]Díaz MA,Cooper RK,Cloeckaert A,et al.Plasmid-mediated high-level gentamicin resistance among enteric bacteria isolated from petturtles in Louisiana[J].Appl Environ Microbiol,2006,72(1):306-312.
[12]邓笑伟,张丹梅,刘长庭,等.嗜麦芽寡氧氨基糖苷类修饰酶与Ⅰ类整合子酶基因的研究[J].中华医院感染学杂志,2009,19(17):2233-2235.
Clone and expression of APH(3′′)-Ⅰand AAC(2′)-Ⅰ gene of Stenotrophomonas maltophilia
GUAN Xiaoshan,GUAN Ruili,ZHONG Huamin,DENG Qiulian,XIE Yongqiang,ZHOU Zhenwen△
(GuangzhouMunicipalWomenandChildren′sMedicalCenter,Guangzhou,Guangdong510120,China)
ObjectiveTo perform the amplification,sequencing and prokaryotic expression of APH(3′′)-Ⅰand AAC(2′)-Ⅰ genes from the clinically isolated gzch810 strain(SM gzch810)of Stenotrophomonas maltophilia to provide the basic materials for the next step functional test.MethodsThe SM gzch810 genome chromosome was extracted,the APH(3′′)-Ⅰ,AAC(2′)-Ⅰ whole genes were amplified by PCR and sequenced after being cloned into pMD18-T vector.The recombination were subcloned into pGEX-4T-1 vector and the expression of the recombinant APH(3′′)-Ⅰand AAC(2′)-Ⅰ were analyzed by SDS-PAGE.ResultsThe 800bp and 550bp DNA fragments of APH(3′′)-Ⅰ,AAC(2′)-Ⅰ gene were amplified from SM gzch810 by PCR and sequenced;the sequence comparison analysis showed that DNA and amino acid sequence identities of APH(3′′)-Ⅰand AAC(2′)-Ⅰ genes with other strains were 91% and 95% respectively.The sequence of APH(3′′)-Ⅰand AAC(2′)-Ⅰ of SM gzch810 were submitted to GenBank(accession number:HQ315852 and HQ315853);two major protein bands corresponding to the expected recombinant GST-TP fusion proteins (56×103and 46×103respectively) were identified by SDS-PAGE.ConclusionAPH(3′′)-Ⅰand AAC(2′)-Ⅰgene of SM gzch810 are successfully cloned and expressed,which lays a good foundation for further detecting corresponding antibiotic resistance and functional evaluation of above two kinds of recombinant E.coli.
Stenotrophomonas maltophilia;AMEs;clone;prokaryotic expression
2016-01-18修回日期:2016-05-05)
关小珊,女,主管技师,主要从事临床微生物检验工作。△
,E-mail:zhouzhenwen28@126.com。
10.3969/j.issn.1673-4130.2016.15.018
A
1673-4130(2016)15-2099-03
猜你喜欢
中草药(2022年9期)2022-05-06
现代畜牧科技(2021年9期)2021-10-13
趣味(作文与阅读)(2021年12期)2021-04-19
童话王国·文学大师班(2021年12期)2021-01-14
特别健康(2018年3期)2018-07-20
商周刊(2017年10期)2017-08-23
中国马铃薯(2015年5期)2016-01-09
现代检验医学杂志(2014年4期)2014-02-02
江西理工大学学报(2013年1期)2013-03-20
郑州大学学报(理学版)(2013年3期)2013-03-11
