
春季休牧后放牧强度变化对短花针茅荒漠草原主要植物种群空间分布的影响
2016-08-10 09:15卫智军张爽吕世杰解云刘文亭刘红梅吴艳玲内蒙古农业大学生态环境学院内蒙古呼和浩特0009内蒙古农业大学理学院内蒙古呼和浩特0008内蒙古自治区林业科学研究院内蒙古呼和浩特0000内蒙古师范大学生命科学与技术学院内蒙古呼和浩特000
生态环境学报 2016年5期
关键词:空间分布
卫智军,张爽,吕世杰,解云,刘文亭,刘红梅,吴艳玲. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 0009;. 内蒙古农业大学理学院,内蒙古 呼和浩特 0008;. 内蒙古自治区林业科学研究院,内蒙古 呼和浩特 0000;. 内蒙古师范大学生命科学与技术学院,内蒙古 呼和浩特 000
春季休牧后放牧强度变化对短花针茅荒漠草原主要植物种群空间分布的影响
卫智军1,张爽1,吕世杰2,解云2,刘文亭1,刘红梅3,吴艳玲4
1. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2. 内蒙古农业大学理学院,内蒙古 呼和浩特 010018;
3. 内蒙古自治区林业科学研究院,内蒙古 呼和浩特 010010;4. 内蒙古师范大学生命科学与技术学院,内蒙古 呼和浩特 010022
摘要:为揭示春季休牧后放牧强度变化对短花针茅(Stipa breviflora)荒漠草原主要植物种群空间分布形式、生态位变化和种内个体关系的影响,引入多重分形理论进行分析。结果表明,主要植物种群短花针茅、碱韭(Allium polyrhizum)和无芒隐子草(Cleistogenes songorica)在各处理区的空间分布均存在多重分形特征(计盒维数D0>信息维数D1>关联维数D2)。短花针茅种群空间分布主要受低密度分布区影响(多重分形谱fΔ<0),碱韭受高密度分布区影响(fΔ>0),无芒隐子草空间分布的主导因素由低密度区(fΔ<0)转为高密度区(fΔ>0)。春季休牧+夏季重牧+秋季适牧(SA2)处理下的主要植物种群空间分布较春季休牧+夏季适牧+秋季重牧(SA1)处理复杂(多重分形谱宽αΔ表现为SA2>SA1)。春季休牧能够使短花针茅植物种群生态位宽度、个体间竞争强度增加(分形维数均较CK增大),且增加程度SA2>SA1;SA1有利于碱韭和无芒隐子草生态位宽度、个体间竞争强度增加(分形维数D0、D2更趋近于2)。
关键词:荒漠草原;春季休牧;植物种群;空间分布;多重分形理论
引用格式:卫智军, 张爽, 吕世杰, 解云, 刘文亭, 刘红梅, 吴艳玲. 春季休牧后放牧强度变化对短花针茅荒漠草原主要植物种群空间分布的影响[J]. 生态环境学报, 2016, 25(5): 729-736.
WEI Zhijun, ZHANG Shuang, LV Shijie, XIE Yun, LIU Wenting, LIU Hongmei, WU Yanling. Effect of Grazing Intensity after Spring Banning Grazing on Spatial Distribution of Dominant Populations in Stipa breviflora Desert Steppe [J]. Ecology and Environmental Sciences, 2016, 25(5): 729-736.
春季休牧作为一项草地保护措施,不仅比较经济可行,也可以使草地植被顺利渡过返青后的“敏感期”(李青丰等,2005;魏德平等,2005)。大量研究证实春季休牧可对草地植被高度、盖度和地上生物量产生正效应(魏德平等,2005;赵钢等,2006;朱立博等,2008);优势种群在群落中的作用增强,杂类草减少,群落物种多样性增加(高娃等,2010;王敏等,2014);群落的结构趋于优化、植被稳定性增加(王敏等,2014);休牧时间的阈值一般为40~50 d(赵钢等,2006;和平等,2008;褚文彬等,2008;运向军等,2010)。由此可见,春季休牧可以有效地保护草原生态环境,使草原植被状况明显改善(李青丰等,2005)。
短花针茅(Stipa breviflora)荒漠草原地处亚洲中部草原区与荒漠区的过渡地带,由于其旱生生境及地域过渡性,生态学上显示出独特性和脆弱性(卫智军等,2013;吕世杰等,2014a);尽管有关该区春季休牧的研究比较广泛(和平等,2008;褚文彬等,2008;运向军等,2010;王敏等,2014),但建群种和优势种空间分布对春季休牧的响应研究鲜有报道(吴艳玲,2012;孙世贤,2014),特别是春季休牧后利用强度变化对其空间分布的影响及植物种群空间分布特征表现形式有待进一步研究。
本研究以短花针茅荒漠草原建群种和优势种为研究对象,采用多重分形分析方法,结合春季休牧后放牧强度变化,探讨建群种短花针茅、优势种碱韭 (Allium polyrhizum) 和无 芒隐 子 草(Cleistogenes songorica)空间分布对春季休牧后放牧强度变化的响应过程和机制,拟揭示其空间分布特点及表现形式,以期为荒漠草原主要植物种群空间分布规律及群落特征变化提供理论依据。
1 试验地概况与研究方法
1.1 试验地概况
研究区域位于内蒙古高原荒漠草原亚带南侧呈条状分布的短花针茅草原,行政区划属于锡林郭勒盟苏尼特右旗赛汉塔拉镇哈登呼舒嘎查,地理位置E112°47′16.9″、N42°16′26.2″,海拔1100~1150 m。年平均气温为 4.3 ℃,最高 38.7 ℃,最低-38.8 ℃。无霜期130 d左右,年平均降水量170~190 mm,年蒸发量2700 mm,雨热同期。土壤为淡栗钙土,地表沙化,腐殖质含量为1.0%~1.8%。植被以亚洲中部荒漠草原种占主导地位,短花针茅为建群种,无芒隐子草和碱韭为优势种,构成了短花针茅+无芒隐子草+碱韭的荒漠草原群落类型。主要伴生种有银灰旋花(Convolvulus ammannii)、木地肤(Kochia prostrata)、阿尔泰狗娃花(Heteropappus altaicus)、栉叶蒿(Neopallasia pectinata)和猪毛菜(Salsola collina)等。草地草层低矮,高度一般为10~35 cm,盖度为10%~50%。
1.2 试验设计
试验设3个处理,分别为对照区(CK,禁牧区)、春季休牧+夏季适牧+秋季重牧处理区(SA1)和春季休牧+夏季重牧+秋季适牧处理区(SA2),采用随机区组试验设计,共分3个区组。放牧试验地总面积46.8 hm2,每个试验区组内小区面积为5.2 hm2。禁牧(休牧)、适度放牧和重度放牧分别放牧苏尼特羊0只、6只和9只,折算载畜率分别为0 hm2、0.87、0.58 hm2·sheep-1·a-1。
1.3 取样方法与数据集建立
在每一处理的 3个重复区选择一个代表性样地,面积100 m×100 m,然后每隔10 m选取一个1 m×1 m样方,按照大(基部丛径>10 cm)、中(基部丛径5~10 cm)、小丛(基部丛径≤5 cm)记录建群种短花针茅、优势种碱韭和无芒隐子草密度,在Excel中按1大丛=2中丛,1中丛=3小丛折算为小丛数建立主要植物种群株丛密度(cluster/m2)数据集。
1.4 研究方法
(1)采用箱式图探讨各植物种群空间分布的集中性和离散性,继而分析其复杂性;利用的统计参数有中位数、上四分位数、下四分位数、最小值和最大值。
(2)多重分形分析(管孝艳等,2013)
多重分形(Multifractal)是描述几何形体、某种质量或测度在不规则的分形空间之上质量分布的定量化工具。采用连续的多重分形谱描述不同尺度、不同层次复杂分形结构的特征(Perfect et al.,2006),为探讨各主要植物种群在不同放牧处理区密度空间分布特征,首先根据多重分形分析的需要建立密度空间分布的概率测度(Scheuring et al.,1994)。
概率计算公式如下:

式中,ε代表取样尺度(100 m2),Mi代表取样尺度下第i个样方种群密度,n为取样尺度为100 m2的样方数目。
在本研究中,概率质量分布函数采用矩法进行计算,在计算之前根据种群密度的概率分布构造配分函数,如下:
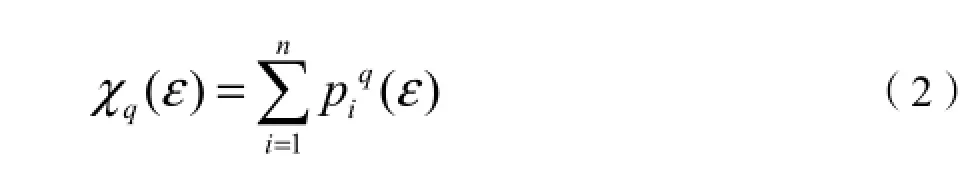
式中,q可取值为-∞<q<+∞(本研究取值为-10≤q≤10),配分函数χq(ε)和取样尺度ε之间满足下式:

式中,τ(q)为q阶质量指数,对于每一个q值对应的质量指数可通过计算logε和log[χq(ε)]之间的拟合曲线的斜率而得到。当q远大于1时,配分函数值主要由较大的数值部分(高密度数据)决定;当q远小于1时,较小值数据(低密度)对于配分函数的贡献率较大。种群密度奇异性指数由 ()qτ曲线的Legendre变换来决定,即:

若研究区域中具有奇异性指数为α的单元个数为 Nα,Nα与取样尺度 ε之间具有幂函数关系Nα=ε-f(α),分形维数f(α)为具有奇异性指数α的分形子集,即:
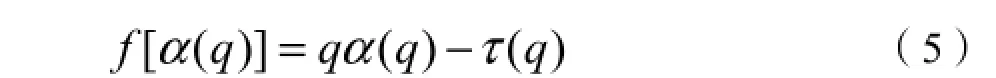
2 结果分析
2.1 主要植物种群变异分析
箱式图不但反应样本空间取样点植物种群密度的集中程度,也反映其分散程度,二者结合可以显示植物种群密度空间分布的复杂程度。箱式图(图1)左、右侧箱线代表其密度数据的分散程度,由图1可知,建群种短花针茅在3个处理区内,其密度变动范围按照CK、SA1和SA2的顺序依次增大,表明短花针茅密度空间分布的变化幅度在增加。从箱体(中间矩形区域)可以看到,按照CK、SA1 和SA2的顺序,箱式图的箱体在逐渐增大,且靠近左端(最小值),表明短花针茅植物种群密度的集中程度在减小,且空间分布以低密度分布为主。
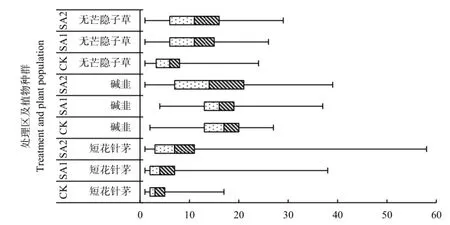
图1 春季休牧对主要植物种群空间分布影响的箱式图Fig. 1 Box diagram of spatial distribution of dominant populations after banning grazing in Spring
优势种碱韭和无芒隐子草植物种群密度空间分布特点总体上与建群种短花针茅相似,即按照CK、SA1和SA2的顺序其密度空间分布的波动范围增大;空间分布的集中区域主要靠近低密度区(碱韭在CK区的空间分布相反)。全年禁牧的CK处理区,主要植物种群空间分布范围的波动性较小,均一化程度较大,且短花针茅和无芒隐子草这种均一化的分布主要由低密度决定,而碱韭由相对的高密度区决定。无论是建群种还是优势种,集中程度SA1均大于SA2处理区,波动程度SA1小于SA2处理区,说明春季休牧+夏季适牧+秋季重牧处理(SA1)能够使建群种和优势种空间分布更趋于均匀化。
2.2 主要植物种群多重分形特征判断
如图2所示,短花针茅植物种群(图2A-1~A-3)logε-log[χq(ε)]双对数曲线图在-10≤q≤10的取值范围内,其配分函数值每一条曲线的直线拟合率均大于90%。其中,当q=-10时,CK、SA1和SA2的直线拟合率分别为95.99%、95.05%和91.44%;当q=10时,CK、SA1和 SA2的直线拟合率分别为95.89%、94.06%和 97.46%。由此说明短花针茅植物种群存在明显的空间分布多重分形特征。
碱韭植物种群(图 2B-1~B-3)在-10≤q≤10的取值范围内,其配分函数值每一条曲线的直线拟合率均大于85%。其中,当q=-10时,CK、SA1和SA2的直线拟合率分别为 89.24%、90.04%和91.72%;当q=10时,CK、SA1和SA2的直线拟合率分别为94.47%、95.43%和95.43%。由此说明碱韭植物种群存在明显的空间分布多重分形特征。
无芒隐子草植物种群(图 2C-1~C-3)在-10≤q≤10的取值范围内,其配分函数值每一条曲线的直线拟合率均大于90%。其中,当q=-10时,CK、SA1和SA2的直线拟合率分别为94.59%、90.17% 和91.14%;当q=10时,CK、SA1和SA2的直线拟合率分别为95.37%、93.51%和95.27%。由此说明无芒隐子草植物种群也存在明显的空间分布多重分形特征。
综合来看,各植物种群在各个处理区内,其logε-log[χq(ε)]关系均呈现随着统计阶距q的增大表现比较一致的变化规律,即在-10≤q≤0的范围内,双对数曲线更逐渐趋于直线,决定系数逐渐趋于1;在 0<q≤10的范围内,双对数曲线逐渐趋于偏离直线的变化趋势,决定系数逐渐小于 1。采用直线进行拟合,其拟合率均大于85%,经统计学显著性检验均达到极显著水平(P<0.01)。由此可见,各植物种群空间分布均存在多重分形特征,可采用多重分形探讨其空间分布特点。
2.3 广义维数谱分析
为进一步探讨植物种群空间分布的多重分形特点,在取样尺度下计算各植物种群在不同处理区的广义多重维数谱q-Dq,结果见图3。短花针茅植物种群广义多重维数谱在-10≤q≤10的范围内,基本呈分散—集中—分散的变化规律。根据多重分形的概率测度pi(ε)∈[0, 1],1<D1<2,其中D1越接近1,说明植物种群密度空间分布在取样尺度的某个区域分布较为集中;D1越接近2,说明其密度空间分布在取样尺度内越趋于均匀分布。短花针茅在取样尺度内,其D1值在CK、SA1和SA2处理区分别为1.766、1.808和1.839。因此,短花针茅受春季休牧的影响,其空间分布趋于均匀化,其均匀程度表现为SA2处理区大于SA1处理区。
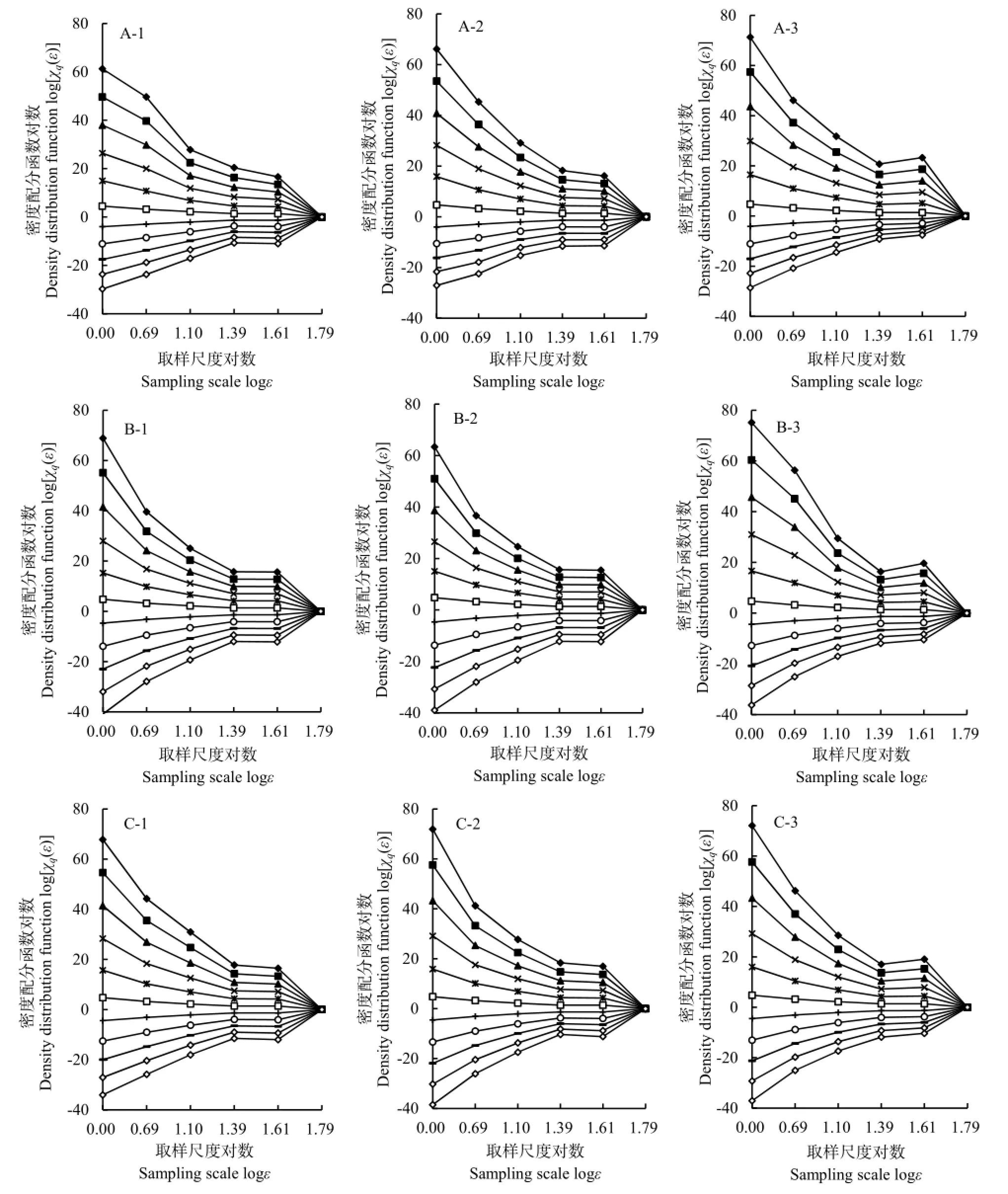
图2 春季休牧条件下取样尺度与植物种群密度的logε-log[χq(ε)]关系Fig.2 logε-log[χq(ε)] relationships between sampling scale and population density after banning grazing in Spring
碱韭植物种群在-10≤q≤10的范围内也呈分散—集中—分散的变化规律,但-10≤q≤0区间的分散程度大于0<q≤10,说明碱韭植物种群低密度区对其空间分布的影响较大;信息维数D1在CK、SA1和SA2处理区分别为1.969、1.971和1.883,表明碱韭在春季休牧条件下,SA2处理区碱韭密度空间分布的均匀程度小于SA1处理区。无芒隐子草植物种群在-10≤q≤10的范围内同样呈分散—集中—分散的变化规律,但-10≤q≤0区间的分散程度小于0<q≤10,这显示出无芒隐子草高密度区对其空间分布影响较低密度区大;D1在CK、SA1和SA2处理区分别为 1.904、1.944和 1.927,表明无芒隐子草在不同放牧处理区主要呈现均匀分布,春季休牧会使无芒隐子草空间分布更为均匀。
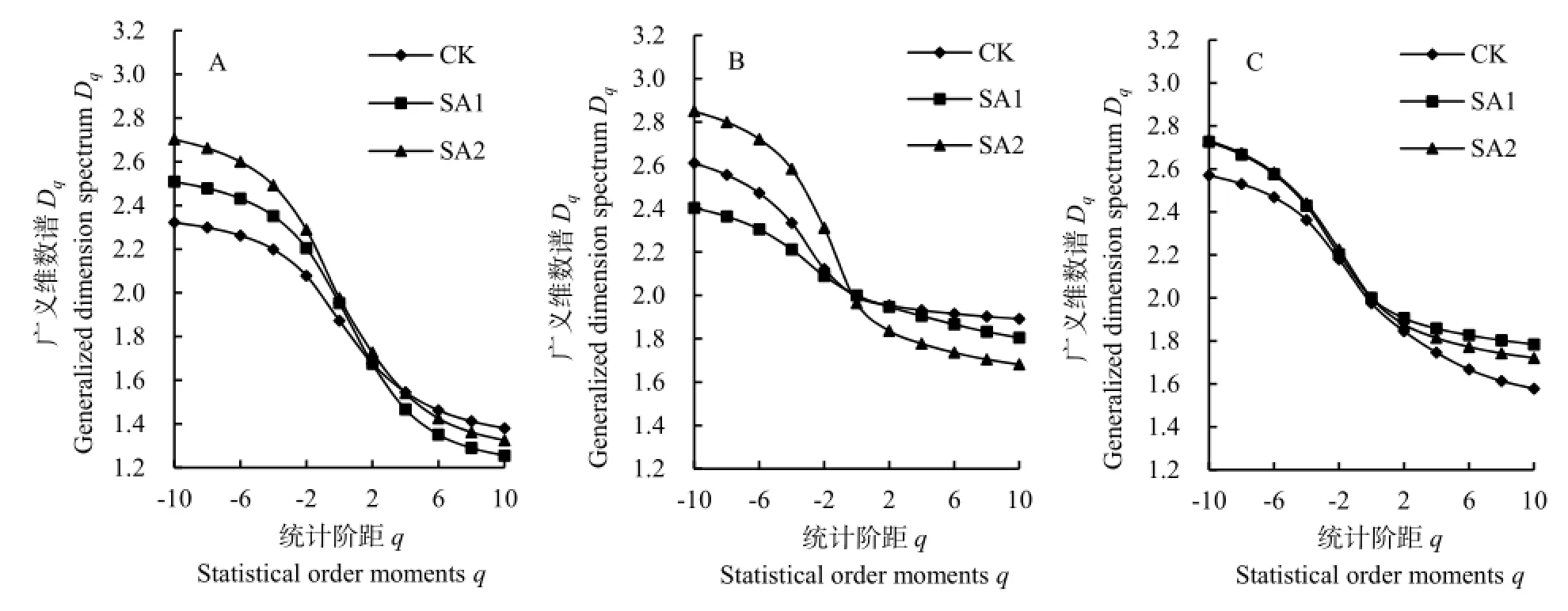
图3 各植物种群在春季休牧下的广义维数谱曲线Fig. 3 Generalized dimension spectrum curve of each plant population after banning grazing in spring
多重维数谱中,当q分别等于0、1和2时,其对应的3个分形维数分别为计盒维数、信息维数和关联维数(见表1)。由表1可知,同一植物种群不同处理区内,计盒维数、信息维数和关联维数尽管存在较大差别,但均存在 D0>D1>D2,这再一次证明3个主要植物种群均存在空间分布的多重分形特征。结合图3不同处理区多重维数谱曲线可知,短花针茅和碱韭空间分布受春季休牧后的放牧强度变化影响大于无芒隐子草,即无芒隐子草的空间分布多重分形特征更稳定。结合多重维数谱和D1/D0的比值来看,春季休牧后无论放牧强度如何变化,短花针茅和碱韭集中在密集区的空间分布形式相对CK区减弱(D1/D0值表现为CK大于SA1 和 SA2);无芒隐子草空间分布集中于密集区的程度SA1> CK>SA2。
2.4 多重分形奇异谱
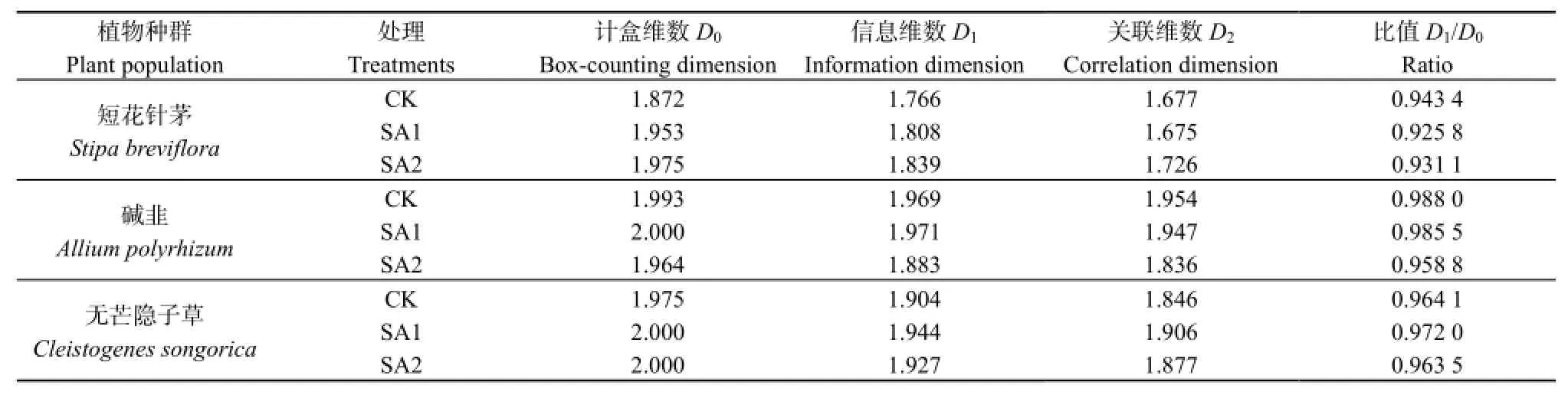
表1 各植物种群在春季休牧下的分形维数和多重分形度Table 1 Fractal dimension and multifractal degree of each plant population after banning grazing in spring
短花针茅多重分形谱曲线 f[α(q)]呈现右钩状(图4),在-10≤q≤10范围内,其表征的是短花针茅空间分布密度较小的样点占主导地位,即其空间变异性更多依赖于密度较小的样点。按照 CK、SA1 和SA2处理区的顺序,短花针茅密度空间分布的变异性依赖低密度分布区的程度减弱。碱韭植物种群密度在CK、SA1和SA2的多重分形谱差别较大,说明其空间分布的变异性存在差别,在CK条件下,其多重分形谱f[α(q)]呈现左钩状,说明碱韭植物种群空间分布密度较大的样点占主导地位,其空间变异性更多依赖于密度较大的样点。SA1和SA2碱韭植物种多重分形谱的对称性也可反映出其空间分布变异性更多依赖于空间分布密度较大的样点(由顶点向 α(q)坐标引垂线,多重分形谱左侧更贴近该锤线)。无芒隐子草密度空间分布多重分形谱基本成开口向下的抛物线形式,但是CK处理区多重分形谱呈右钩状,SA1处理区呈左钩状,因此,CK处理区无芒隐子草空间分布变异性更多依赖于空间分布密度较小的样点,而SA1处理区反之。
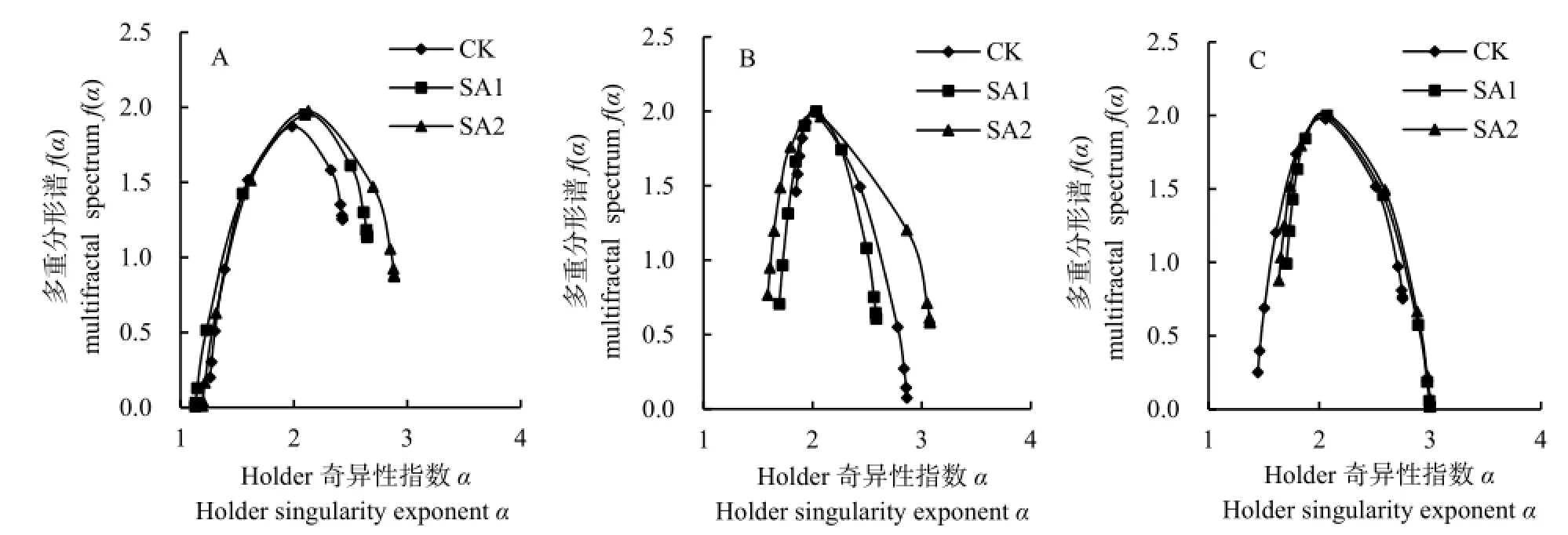
图4 各植物种群在春季休牧下的多重分形奇异谱曲线Fig. 4 Multifractal singular spectrum curve of each plant population after banning grazing in spring
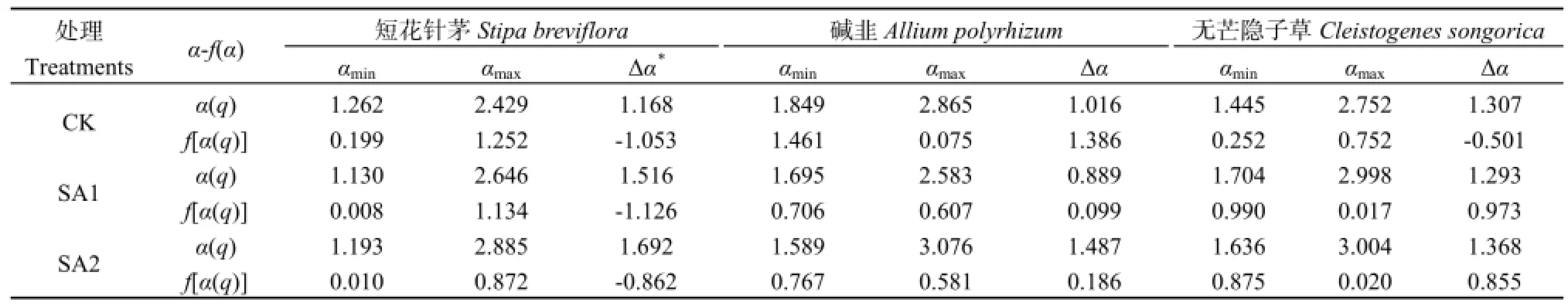
表2 各植物种群在春季休牧下的多重奇异谱参数Table 2 Multiple singular spectrum parameters of each plant population after banning grazing in spring
根据多重分形谱宽Δα=αmax-αmin可知(表 2),短花针茅植物种群Δα表现为SA2>SA1>CK,碱韭植物种群Δα表现出SA2>CK>SA1,无芒隐子草植物种群Δα为SA2>CK>SA1。可见春季休牧后,由于夏秋季放牧强度不同导致不同植物种群Δα存在差异,Δα越大表明其密度空间分布越不均匀。短花针茅春季休牧后,无论后期放牧强度如何,其空间分布的非均匀程度增加。春季休牧+夏季重牧+秋季适牧(SA2)导致碱韭和无芒隐子草植物种群密度空间分布的复杂性较 CK处理区增加,春季休牧+夏季适牧+秋季重牧(SA1)使其密度空间分布较CK处理区更加均匀。
短花针茅Δf[Δf=f(αmin)-f(αmax)]表现为 SA1<CK<SA2<0,短花针茅植物种群多重分形谱主要呈右钩状,其密度空间分布变异性主要由密度较小的分布区域引起。碱韭Δf表现为 0<SA1<SA2<CK,碱韭植物种群主要呈左钩状,说明其空间分布变异性主要由密度较大分布区引起的。无芒隐子草Δf表现为CK<0<SA2<SA1,因此无芒隐子草在CK处理区多重分形谱呈右钩状,其密度空间分布变异性主要由密度较小的分布区域引起;SA1和SA2处理区多重分形谱呈左钩状,其密度空间分布变异性主要由密度较大的分布区域引起。
3 讨论
建群种短花针茅和优势种碱韭、无芒隐子草是荒漠草原的主要植物种群,其地上现存量占群落现存量的比例可达80%以上,3个植物种群数量消涨、时空变化及结构的位移,构成了短花针茅+碱韭+无芒隐子草的群落类型(吕世杰等,2014b),且各植物种的空间分布具有特异性(黄琛等,2014)。本研究结果显示,建群种短花针茅、优势种碱韭和无芒隐子草的空间分布受春季休牧后放牧强度差异的影响变化规律不同,因此植物种空间分布对春季休牧后的放牧强度响应具有特异性。同时3个植物种群地上现存量较大,受放牧家畜采食、排泄以及践踏的影响,不同试验处理导致群落表现出的种群结构和比例均会发生明显变化,从而导致种群间的种间关系及其自身的生态位发生变化(吕世杰,2014a,2014b)。
箱式图可定性反映数据序列的集中程度和离散程度,但存在局限性(刘红梅等,2013;希吉日塔娜等,2013)。在本研究结果中,不同植物种群由于空间分布特点存在差异,导致箱式图不能完全揭示3个主要植物种群空间分布特点。在广义多重维数谱分析中,植物种群多重维数谱因放牧处理不同而不同,说明春季休牧时间相同但后期放牧强度变化,导致植物种群空间分布发生变化。计盒维数(D0)、信息维数(D1)和关联维数(D2)是变量空间分布主要探讨的分形维数(李鹏远等,2008;刘红梅等,2011;茹豪等,2015)。一般认为(Posadas et al.,2003;茹豪等,2015),D0>D1>D2,变量存在空间多重分形特征;当 D0=D1=D2,变量具有空间自相关或均匀分布特征。因此,本研究主要植物种群均存在明显的多重分形特征。D1/D0比值越接近1,植物种群空间分布越集中于密集区域;D1/D0比值越接近于 0,植物种群空间分布越集中于稀疏区域(卞正富,2004;吕春娟等,2011;王金满等,2014)。本研究结果D1/D0比值均大于0.9,接近于1,说明主要植物种群空间分布集中于密集区。3个分形维数具有不同的理论意义(倪红伟,2000a,2000b;张金屯,2011),计盒维数 D0反映植物种群空间占有能力(反映生态位大小),信息维数D1反映植物种群空间分布的均匀性(反映格局强度),关联维数反映植物种群空间分布的自相关强度(反映种群个体竞争强弱)。因此,建群种短花针茅按照CK、SA1和SA2处理区的顺序,其生态位宽度增加,空间分布均匀性增大,个体间的竞争强度增加;碱韭植物种群在中SA1处理区较CK和SA2相比,生态位宽度最宽,空间分布的格局简单,个体间的竞争强度大于SA2而小于CK处理区;无芒隐子草植物种群空间分布特点在SA1处理区具有与SA2同样大的生态位宽度,但SA1处理区无芒隐子草空间分布更加均匀,个体间的竞争强度更大。
多重分形分析的前提首先是检验变量是否具有多重分形特征,文中已经采用了 logε-log[χq(ε)]关系进行判定(管孝艳等,2013)。采用τ″(1)=τ(2)-2τ(1)+τ(0)同样能够判定变量是否存在多重分形特征(陈志军,2007),根据τ(q)和D(q)的关系,由于τ(0)=-D0,τ(1)=0,τ(2)=D2,因此 τ″(1)=τ(2)+τ(0)= D2-D0。由于τ″(1)<0表征变量具有多重分形特征,τ″(1)=0表征变量为单分形或非分形(Cheng,1999;陈志军,2007),这一判定方法与D0>D1>D2的判定方法具有相似之处。因此,无论采用哪一种方法进行判断,短花针茅荒漠草原主要植物种群空间分布均具有多重分形特征。根据多重分形奇异谱极值,多重分形谱宽度Δα=αmax-αmin反映整个分形结构研究变量的非均匀程度和受概率测度大的区域影响程度(王金满等,2014),所以在春季休牧+夏季重牧+秋季适牧处理区短花针茅、碱韭和无芒隐子草植物种群分布的非均匀程度较大,但短花针茅植物种群受低密度区影响较大,而碱韭和无芒隐子草受高密度区的影响较大。这与采用分形维数 D1的分析结果存在偏差,因此有研究者(刘继龙等,2010)认为,多重分形更能揭示变量空间分布特征。本研究认为,这是由于两种分析方法阐释问题的角度不同而导致的,单一分形主要是揭示研究尺度变量空间分布整体分布形式,而多重分形探讨的是某一尺度概率分布随统计阶距的变化规律,可以刻画整体中局部或小尺度特征。多重分形谱形状特征由Δf=f(αmin)-f(αmax)反映,Δf<0表征f[α(q)]呈右钩状,小概率子集占主导地位;反之成立(王金满等,2014)。由此可见,短花针茅植物种群空间分布主要由低密度分布区决定,且SA1处理低密度分布区对其空间分布的主导作用最大;碱韭植物种群Δf>0,f[α(q)]形状呈左钩状,其空间分布主要由高密度分布区起主导作用,无芒隐子草植物种群空间分布的主导因素由低密度区(CK)转为高密度区(SA1和SA2),这再一次证明植物种群空间分布因种群不同而存在特异性(黄琛等,2014)。
综上所述,首先,无论放牧强度改变与否,主要植物种群的空间分布多重分形特征均存在;其次,广义维数谱中的计盒维数(D0)、信息维数(D1)和关联维数(D2)不仅可作为判定植物种群空间多重分形分析能否进行的依据,也可反映植物种群生态位大小、格局复杂程度以及种内个体竞争强度;第三,多重分形可揭示植物种群空间分布的均匀程度及其高低分布区的作用大小。这与张金屯(2011)在数量生态学中提到的观点存在偏差,其认为种群空间分布研究采用点格局会更好。本研究认为植物种群空间分布是其与环境及放牧利用长期适应和选择的结果,具有复杂的生态学过程和特定的群落内部调节机制;任何可从某一特定角度揭示种群空间分布特征的研究方法均值得推敲和应用,将多重分形理论引入草地植物种群空间分布研究,可丰富植物种群空间分布研究方法,也可为进一步揭示植物种群空间分布特征和变化规律提供理论研究依据。
4 结论
建群种短花针茅、优势种碱韭和无芒隐子草空间分布对春季休牧后放牧强度变化的响应具有特异性,且均明显存在多重分形特征。主要种群多重分形特征与种群自身生态位宽度、个体间的竞争强弱直接相关,反映了植物种群空间分布的复杂性。多重分形奇异谱显示,春季休牧后,春季休牧+夏季重牧+秋季适牧处理区建群种空间分布的不均匀程度大于春季休牧+夏季适牧+秋季重牧处理区,但主导这种不均匀强度的密度分布区存在差异。
参考文献:
CHENG Q M. 1999. Multifractality and spatial statistics [J]. Computers & Geosciences, 25(9): 949-961.
PERFECT E, GENTRY R W, SUKOP M C, et al. 2006. Multifractal Sierpinski carpets: Theory and application to upscaling effective saturated hydraulic conductivity [J]. Geoderma, 134(3-4): 240-252.
POSADAS A N D, GIMENEZ D, QUIROZ R, et al. 2003. Multifractal characterization of soil pore systems [J]. Soil Science Society of America Journal, 67(5): 1361-1369.
SCHEURING I, RIEDI R H. 1994. Application of multifractal to the analysis of vegetation pattern [J]. Journal of Vegetable Science, 5(4): 489-496.
卞正富. 2004. 矿区开采沉陷农用土地质量空间变化研究[J]. 中国矿业大学学报, 33(2): 213-218.
陈志军. 2007. 多重分形局部奇异性分析方法及其在矿产资源信息提取中的应用[D]. 武汉: 中国地质大学.
褚文彬, 卫智军, 运向军, 等. 2008. 短花针茅荒漠草原土壤含水量和地上现存量对禁牧休牧的响应[J]. 中国草地学报, 30(3): 106-109.
高娃, 段新桥, 金彩霞, 等. 2010. 锡林郭勒草原休牧效果动态监测[J].畜牧与饲料科学, 31(Z1): 327-330.
管孝艳, 王少丽, 吕烨, 等. 2013. 土壤水盐空间变异性及其尺度效应的多重分形分析[J]. 水利学报, 44(Z1): 8-14.
和平, 褚文彬, 运向军, 等. 2008. 短花针茅荒漠草原群落现存量和营养动态对禁牧休牧的影响[J]. 当代畜禽养殖业, (6): 41-44.
黄琛, 张宇, 王静, 等. 2014. 不同放牧强度下短花针茅荒漠草原植被的空间异质性[J]. 植物生态学报, 38(11): 1184-1193.
李鹏远, 程积民, 万惠娥, 等. 2008. 云雾山自然保护区优势植物种群分布格局的分形特征[J]. 生态学报, 28(10):5161-5165.
李青丰, 赵钢, 郑蒙安, 等. 2005. 春季休牧对草原和家畜生产力的影响[J]. 草地学报, 13(S1): 53-56, 66.
刘红梅, 王娜, 吕世杰, 等. 2013. 蒙古栎幼树生长状况调查研究[J]. 内蒙古林业科技, 39(1): 15-17, 26.
刘红梅, 卫智军, 杨静, 等. 2011. 不同放牧制度对荒漠草原短花针茅空间异质性的影响[J]. 干旱区资源与环境, 25(8): 138-143.
刘继龙, 马孝义, 张振华. 2010. 不同土层土壤水分特征曲线的空间变异及其影响因素[J]. 农业机械学报, 41(1): 46-52.
吕春娟,白中科,陈卫国. 2011. 黄土区采煤排土场生态复垦工程实施成效分析[J]. 水土保持通报, 31(6): 232-236.
吕世杰, 刘红梅, 吴艳玲, 等. 2014a. 放牧对荒漠草原建群种和优势种生态位的影响[J]. 生态环境学报, 23(10): 1600-1609.
吕世杰, 刘红梅, 吴艳玲, 等. 2014b. 放牧对短花针茅荒漠草原建群种与优势种空间分布关系的影响[J]. 应用生态学报, 25(12): 3469-3474.
倪红伟, 郭亚胜, 吴海一. 2000b. 小叶章种群分布格局的分形特征Ⅱ信息维数与关联维数[J]. 植物研究, 20(3): 351-354.
倪红伟, 马克明, 赵伏臣. 2000a. 小叶章种群分布格局的分形特征Ⅰ计盒维数[J]. 植物研究, 20(2): 229-234.
茹豪, 张建军, 李玉婷, 等. 2015. 黄土高原土壤粒径分形特征及其对土壤侵蚀的影响[J]. 农业机械学报, 46(4): 176-182.
孙世贤. 2014. 短花针茅荒漠草原群落特征和土壤对放牧强度季节调控的响应[D]. 呼和浩特: 内蒙古农业大学.
王金满, 张萌, 白中科, 等. 2014. 黄土区露天煤矿排土场重构土壤颗粒组成的多重分形特征[J]. 农业工程学报, 30(4): 230-238.
王敏, 卫智军, 孙世贤, 等. 2014. 短花针茅草原群落特征对全年放牧和春季休牧的响应[J]. 草原与草业, 26(3): 51-56.
卫智军, 韩国栋, 赵钢, 等. 2013. 中国荒漠草原生态系统研究[M]. 北京: 科学出版社.
魏德平, 达布希拉图, 张春信, 等. 2005. 锡林郭勒盟 2005年春季休牧监测研究[J]. 内蒙古草业, 17(3):7-9.
吴艳玲. 2012. 短花针茅草原群落特征与空间异质性对放牧强度季节调控的响应[D]. 呼和浩特: 内蒙古农业大学.
希吉日塔娜, 吕世杰, 卫智军, 等. 2013. 不同放牧制度下短花针茅草原主要植物种群的空间变异[J]. 中国草地学报, 35(2): 76-82.
运向军, 卫智军, 杨静, 等. 2010. 禁牧休牧短花针茅草原地上现存量与土壤含水量的关系[J]. 中国草地学报, 32(2): 75-79.
张金屯. 2011. 数量生态学(第二版) [M].北京: 科学出版社.
赵钢, 李青丰, 张恩厚. 2006. 春季休牧对绵羊和草地生产性能的影响[J]. 仲恺农业技术学院学报, 19(1):1-4.
朱立博, 曾昭海, 赵宝平, 等. 2008. 春季休牧对草地植被的影响[J]. 草地学报, 16(3): 278-282.
DOI:10.16258/j.cnki.1674-5906.2016.05.001
中图分类号:X17; Q948
文献标志码:A
文章编号:1674-5906(2016)05-0729-08
基金项目:国家自然科学基金项目(31460126);内蒙古自然科学基金项目(2015MS0349);国家重点基础研究计划项目(2014CB138800);内蒙古自治区高等学校科学研究项目(NJZC16073);内蒙古农业大学基础研究项目(JC2015002)
作者简介:卫智军(1957年出生),男,教授,博士生导师,主要从事草地放牧生态与管理研究。E-mail: nmndwzj@163.com
收稿日期:2015-10-30
Effect of Grazing Intensity after Spring Banning Grazing on Spatial Distribution of Dominant Populations in Stipa breviflora Desert Steppe
WEI Zhijun1, ZHANG Shuang1, LV Shijie2, XIE Yun2, LIU Wenting1, LIU Hongmei3, WU Yanling4
1. College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Hohhot 010019, China;2. College of Science, Inner Mongolia Agricultural University, Hohhot 010018, China; 3. Inner Mongolia Academy of Forestry Science, Hohhot 010010, China;4. College of Life Science and Technology, Inner Mongolia Normal University, Hohhot 010022, China
Abstract:In this paper, we introduced the multi-fractal theory in order to investigate the grazing intensity after banning grazing in spring how to influence the spatial distribution pattern, niche changes and the effects of the individual relations of plant population. The results shown that there was unique multi-fractal characteristics of spatial distribution on Stipa breviflora, Allium polyrhizum and Cleistogenes songorica in each grazing treatment (box-counting dimension D0> information dimension D1> correlation dimension D2). Population Stipa breviflora is mainly affected by the low density distribution (multifractal spectrum Δf < 0), while that of Allium polyrhizum is mainly influenced by the high density area (Δf > 0), and the dominant factor in the spatial distribution of Cleistogenes songorica turned low-density area (Δf < 0) to a high-density area (Δf > 0). Banning grazing in spring + heavy grazing in summer + moderate grazing in autumn (SA2) led to a more complex spatial distribution relatively to banning grazing in spring + moderate grazing in summer + heavy grazing in autumn (SA1) (a wide multifractal spectrum Δα showed SA2 > SA1). Banning grazing in spring can made niche breadth and individuals competition intensity of population Stipa breviflora increased (fractal dimension increased than CK area), and increased the degree of SA2 > SA1. Moreover, increasing of SA1 is benefit to increase the niche breadth and individuals' competition intensity of population Allium polyrhizum and Cleistogenes songorica (fractal dimension D0, D2more closed to 2).
Key words:desert steppe; banning grazing in spring; plant population; spatial distribution; multi-fractal theory
猜你喜欢
南水北调与水利科技(2017年1期)2017-02-27
中国人口·资源与环境(2016年11期)2017-02-17
安徽农学通报(2016年24期)2017-01-12
绿色科技(2016年20期)2016-12-27
科技视界(2016年18期)2016-11-03
科技视界(2016年13期)2016-06-13
湖北农业科学(2015年16期)2015-10-28
湖北农业科学(2015年17期)2015-10-09
地球科学与环境学报(2015年4期)2015-07-31
