
中国黑戈壁植物多样性分布格局及其影响因素
2016-08-09 05:52:13王健铭董芳宇巴海那斯拉李景文李俊清冯益明
生态学报 2016年12期
王健铭,董芳宇, 巴海那斯拉,3,李景文,*, 李俊清,冯益明, 卢 琦
1 北京林业大学林学院,北京 100083 2 中国林业科学研究院荒漠研究所,北京 100091 3 新疆重点林业工程质量管理总站, 乌鲁木齐 830000
中国黑戈壁植物多样性分布格局及其影响因素
王健铭1,董芳宇1, 巴海那斯拉1,3,李景文1,*, 李俊清1,冯益明2, 卢琦2
1 北京林业大学林学院,北京100083 2 中国林业科学研究院荒漠研究所,北京100091 3 新疆重点林业工程质量管理总站, 乌鲁木齐830000
摘要:我国西北地区内陆分布着近20万km2的黑戈壁,由于其环境的特殊性,使其具有独特的生态系统,境内分布着多样的植被,蕴藏着大量特有的自然资源。但由于自然环境苛刻与交通条件不便,目前我国关于黑戈壁区系统的植被与物种多样性的研究还很缺乏。针对黑戈壁区植物多样性组成与分布特点,基于遥感及实地调查,采用DCCA排序和半变异函数模型等分析方法,对黑戈壁区植物群落组成,植物多样性特点及影响因素进行分析。研究结果表明:研究区植物以藜科和蒺藜科灌木或半灌木为主,群落物种生活型具有逐渐趋于简单甚至单一的特性,重要值>0.1的植物主要有梭梭、红砂、白刺等13种;群落物种多样性呈现区域性的斑块化分布,结构性因子引起的物种多样性空间异质性占主导地位;作为极端干旱区,该区植物群落类型具有贫乏化及单一化的趋势,群落结构简单,植被覆盖度低,植物生长随环境的变化具有明显的可塑性,群落空间分异明显,群落空间演变具有明显的水分及土壤结构梯度;DCCA结果显示气候、土壤、地形是群落物种及类型变化的主要原因,海拔、坡位、土壤机械组成、降水、温度等环境因子对群落有着显著的影响,水土条件的空间异质性是戈壁植物多样性维持的关键因素。
关键词:黑戈壁;植物群落;物种多样性;分布;因素
植物多样性作为群落的重要特征,是生态系统结构、功能及其稳定性的基础[1- 2]。随着生态研究的不断深入,植物多样性格局形成机制研究成为了生态学热点问题[3-4],该方面的研究是理解区域生物多样性维持机制重要的内容;同时对生物多样性的保护管理等有着重要意义[5-8]。我国在生物多样性格局与维持机制方面开展了很多卓有成效的研究,但对于西北干旱区,特别是自然环境严苛,交通不便的极端干旱的戈壁区,这方面工作还很缺乏。目前关于戈壁植物多样性的研究仍然较少,国内相关研究主要是植被地理与植物分布[9-10]、植物区系特点[11-12],以及周边区域的相关研究[13- 14],近年来,德国、日本学者在我国西北和蒙古国,主要是在蒙古国一些戈壁地区开展了一些研究工作,包括植被及物种多样性分布与环境因子关系[15-18],气候变化对群落生产力影响以及植被生长与沙尘发生关系[18-21]等。而黑戈壁区至今还没有系统的植物多样性分布格局的研究。
黑戈壁区位于甘肃与新疆之间,为东起额济纳河,北抵中蒙界山,南临河西走廊西段,西依天山东段的一个大约20万km2的区域。该区年降水量在100mm以下,多数区域低于50mm,蒸发量极高,为我国极端干旱区。受严苛环境及交通不便的影响,目前黑戈壁这一戈壁荒漠中最干旱区域的植被与物种多样性格局的系统研究尚未开展。而随着西部开发及丝绸之路经济带发展,正在建设的京新高速公路及铁路直穿黑戈壁腹地,同时矿产资源开发范围扩大,人为干扰对该区脆弱的生态系统及其植被的影响日益严重。未来该区自然环境维护和植被及其物种多样性的保护也急需全面系统的本底调查。
针对我国黑戈壁植物多样性分布格局的研究缺乏问题,从2011年开始由中国林科院荒漠研究所牵头,组织了多个单位对黑戈壁地区进行了包括地貌特征、水文、土壤、气候与动植物资源等生态本底的综合考察[22]。本文在黑戈壁区近2年的植物多样性全面调查基础上,重点分析黑戈壁区植物多样性分布格局及其主要影响因素,阐明黑戈壁植物多样性分布特征及其特殊性;为黑戈壁区植物多样性保护、区域生态环境维持提供理论依据。
1研究方法
1.1调查区域与线路的确定
本研究依据中国黑戈壁综合科学考察线路,设计植被调查的地区和线路,调查区域尽量覆盖黑戈壁不同地理区域、地形地貌特征、植被类型等,于2011—2012年生长季沿着调查线路对甘肃敦煌、肃北,新疆哈密、伊吾、巴里坤,内蒙额济纳旗等典型黑戈壁区进行植物本底调查,调查线路总长度达到4900km。
1.2调查方法
依据规划的调查线路,在植被分布区内,根据植被组成、水土条件的变化梯度布设样点,在每个样点内根据植被分布面积大小和组成复杂程度设置3—5个10m×10m的临时样方,共设置样点56个,临时样方174个;在每个临时样方内调查植被类型、物种的组成。植物物种调查指标包括地理坐标、海拔、地形特征、戈壁类型[22]、砾石覆盖度与粒级,群落特征包括群落类型、群落总盖度、植物物种盖度、主要物种组成、个体数量。记录样方内所有植物的种类、株数、盖度、平均高度、生长状况、物候、分布情况等。
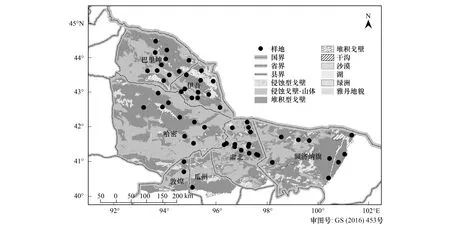
图1 调查样点分布图Fig.1 Distribution of survey samples
2数据处理
2.1群落特征数据及环境相关性分析
依据《中国植被》[23]的分类原则,依据生境特点、群落结构与外貌、物种优势度及物种组成划分植物群落[24]。根据临时样方内植物个体高度、盖度、密度以及频度,计算物种的重要值:重要值=(相对密度+相对频度+相对盖度+相对高度)/4[5- 7,25];相对密度=某一种个体数/所有种的个体数总和,相对盖度=某物种的盖度/全部种的盖度总和,相对高度=某一种植株平均高度/所有种植株平均高度总和,相对频度=某物种的频度/所有种的频度之和。依据物种多样性测度指数应用的广泛程度以及对群落物种多样性状况的反映能力,本文选取以下4种多样性指数来测度和分析群落物种多样性特征[5- 7,25]:
(1)Simpson指数(D)
(2)Shannon-Wiener 指数(H)
(3)Pielou 均匀度指数(J)
J=H/lnS
(4)Margalef丰富度指数(M)
M=(S-1)/lnN
式中,S为样方内物种数;N为样方内所有物种个体数总和;Pi为种i的相对重要值。
利用SPSS 22软件对数据进行均值分析,并对群落环境因子及植物群落特征与多样性指数等数据进行相关性系数及显著性分析。
2.2植物群落空间分布格局及环境相关性分析
因样方中分布频率较低物种的存在,会影响排序结果,使最终解释与实际有所偏差[25- 27],所以本文在排序分析时,利用CANOCO 4.5软件设置处理的方法剔除样方分布频率小于5%的植物[13,25- 27]。最后得到162(样方)×36(物种)维物种数据矩阵。环境数据包括年平均温度、最热月均温、年降水量、海拔、坡度、坡位、砾石盖度、砾石径级等8个环境因子,得到162(样方)×8(环境变量)维的环境数据矩阵;其中气候数据来取自于中国黑戈壁综合科学考察队气候组数据库[22]。实际分析中使用CANOCO 4.5软件包中的蒙特卡罗拟合(Monte Carlo permutation test)检验植物群落组成与环境变量之间的相关显著性,以及运用偏典范对应分析(pDCCA)分离环境变量对植物群落变异的相对贡献率[26- 28]。
2.3群落物种多样性空间格局及多样性预测分析
利用GS+9.0软件对群落物种多样性空间分布进行半变异函数与方差模型分析,并在模型和方差分析的基础上,在ArcGIS 10.2上,利用普通克里金插值法,绘制黑戈壁物种多样性预测图。
3结果与分析
3.1黑戈壁植物种类及群落数量特征分析
调查中记录黑戈壁区植物154种,分属28科,85属,重要值与分布频度较高的植物主要有梭梭(Haloxylonammodendron)、红砂(Reaumuriasongarica)、芨芨草(Achnatherumsplendens)、白刺(Nitrariatangutorum)、沙拐枣(Calligonummongolicum)、膜果麻黄(Ephedraprzewalskii)、霸王(Sarcozygiumxanthoxylon)、合头草(Sympegmaregelii)、泡泡刺(Nitrariasphaerocarpa)、戈壁藜(Iljiniaregelii)、盐生草(Halogetonglomeratus)、芦苇(Phragmitesaustralis)、柽柳(Tamarixhohenackeri)等13种植物,为黑戈壁主要优势植物(表1)。总体上,黑戈壁优势植物以藜科和蒺藜科灌木或半灌木为主,草本所占的比例很小,反映了黑戈壁环境严苛和异质性程度低的条件下,植被组成物种生活型逐渐趋于简单甚至单一的特性。

表1 优势植物种类及其重要值
红砂在所有样方中出现的频率最高,是分布范围最广的物种,梭梭、膜果麻黄、沙拐枣、白刺等植物也是黑戈壁主要的优势物种,在样方内出现频率也较高;但在不同生境中植物的优势度及生长指标差异很大,反映了植物对不同环境梯度具有生长适应性。柽柳、芦苇等群落,主要分布在季节性积水河道、湖盆或有地质断裂带的泉水生境中,属于该区隐域性植被,分布范围小,生境较为特殊。而戈壁针茅(StipatianschanicaRoshev vargobica)等主要分布在黑戈壁海拔相对较高的边缘地区及荒漠草原的过渡区内。

图2 主要群落数量特征Fig.2 Quantitative characteristics of the major plant communities1—13分别代表:戈壁藜、梭梭、柽柳、芦苇、红砂、芨芨草、膜果麻黄、合头草、霸王、白刺、泡泡刺、沙拐枣、盐生草群落
如图2所示,不同优势群落间植被盖度及高度、群落植株密度存在极显著的差异(P<0.01),物种丰富度也存在显著差异(P<0.05),戈壁藜、白刺、泡泡刺、盐生草等群落主要分布在地表粗糙度高的砾石堆积戈壁,植被稀疏,群落覆盖度低,群落植物个体密度变异明显;霸王、合头草等群落多分布在海拔较高、降水相对丰富的地区,其植被生长较为良好,物种丰富度也较高;而柽柳、芦苇等隐域性植被,多分布在砾石覆盖少的地势较低区域,其植被高度和盖度情况最好。
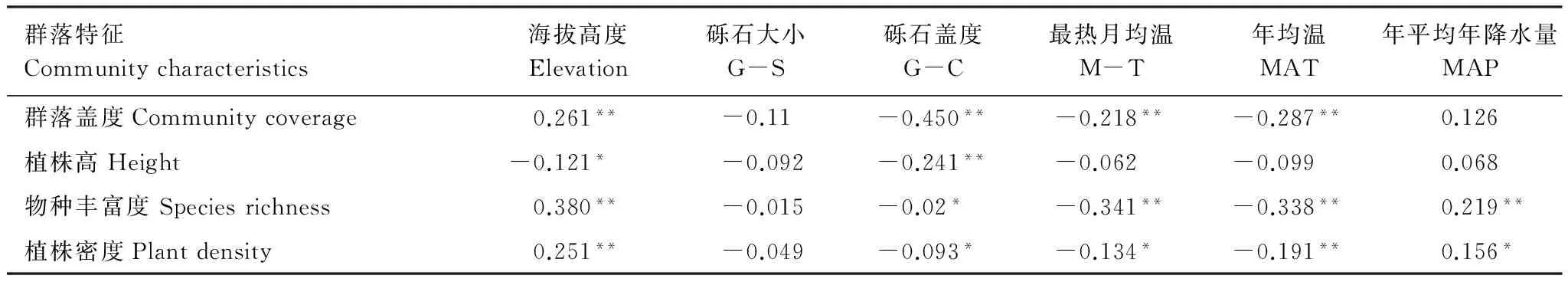
表2 群落数量特征与环境因子Pearson相关性分析
**P<0.01, *P<0.05
由表2可知,群落物种丰富度与温度、砾石盖度呈显著负相关,与年平均降水量及海拔呈正相关;群落植被盖度与海拔呈显著正相关,与温度及砾石盖度呈负相关;其植被高度仅与海拔高度及砾石盖度呈负相关,与其它环境因子则没有显著关系;环境因子与群落植株密度的关系和其与植被盖度的关系相似,不同之处在于群落植株密度还与降水呈正相关。这表明群落数量特征与海拔、地表覆盖物、水热条件有着密切的关系,地形及大气路径导致的水热条件在空间上的异质性[22]对植物群结构和组成存在显著影响,植物地上部分随着环境梯度的变化具有生长(盖度及高度)的可塑性。
3.2黑戈壁植物群落DCCA排序分析
通过对数据进行DCCA排序分析,并在CANOCO 4.5软件包中对排序轴进行蒙特卡罗拟合检验,结果显示物种变量与环境变量之间呈现极显著的相关性(P=0.0001),表明环境因子对群落的物种组成具有显著的影响。与此同时,对8个环境变量进行Monte Carlo检验,检验变量是否存在冗余现象,结果显示8个环境因子均与植物群落存在显著的相关性,不存在冗余现象(P<0.001)。

图3 DCCA植物物种排序图Fig.3 Plant species ordination diagram of DCCA图中列出了物种加权值大于10%的优势物种:Ⅰ,泡泡刺Nitraria sphaerocarpa;Ⅱ,白刺Nitraria tangutorum;III,灌木亚菊Ajania fruticulosa;IV,短叶假木贼Anabasis brevifolia;Ⅴ,五柱红纱Reaumuria kaschgarica ;Ⅵ,红砂Reaumuria songarica;Ⅶ,松叶猪毛菜Salsola laricifolia ;Ⅷ,柠条锦鸡儿Caragana Korshinskii;IX,白沙蒿Artemisia blepharolepis ;Ⅹ,膜果麻黄Ephedra przewal;Ⅺ,合头草Sympegma regelii;Ⅻ,芨芨草Achnatherum splendens;,木本猪毛菜Salsola arbuscula;,芦苇Phragmites australis;XV,胀果甘草Glycyrrhiza inflata ;XVI,柽柳Tamarix hohenackeri;XVII,黑果枸杞Lycium ruthenicum;XVIII,戈壁针茅Stipa tianschanica Roshev var gobica;XIX,密头蒿Tanacetum crassipes;XX,沙拐枣Calligonum mongolicum;XXI,骆驼刺Alhagi sparsifolia ;XXII,花花柴Karelinia caspia;XXIII,盐生草Halogeton glomeratus;XXIV,戈壁藜Iljinia regelii;XXV,梭梭Haloxylon ammodendron;;XXVI,霸王Sarcozygium xanthoxylon;XXVII,甘青铁线莲Clematis tangutica;XXVIII,驼绒藜Ceratoides laten;XXIX,白皮锦鸡儿Caragana leucophloea;XXX,木旋花Convolvulus fruticosus;A-T,年均温;M-T:最热月均温;A-P:年平均降雨量;Elevation:海拔;G-S:砾石大小;Slope:坡度:G-T:坡位;G-C:砾石盖度
采用8个环境变量,对研究区植物群落格局与环境的关系进行排序分析,结果显示第一和第二轴的特征值分别为0.655、0.396,贡献率分别为0.492、0.297,前两轴特征值之和占全部特征值总和的78.9%,包含了绝大多数的排序信息,因此采用前两轴的数据来分析环境因子与植物群落的关系。
从图3可以看出,DCCA排序图第一轴主要反映海拔、气候、坡位等环境因子对植物分布的影响;其中海拔对植物群落分布影响最显著, 地形是植物所处环境气候、地表覆盖度产生差异的重要原因,芦苇、柽柳、花花柴(Kareliniacaspia)等植被主要分布在地势较低、温度较高的季节性河流、湖盆生境,红砂、梭梭等典型的戈壁植物则分布在广阔的山前戈壁生境上,而戈壁针茅、密头蒿(Tanacetumcrassipes)、灌木亚菊(Ajaniafruticulosa)、五柱红砂(Reaumuriakaschgarica)等植物主要分布在海拔较高、降水较好的生境,形成戈壁向草原过渡的植被。第二轴主要反映了地形起伏、地表覆盖、土壤质地对植物群落空间分布的影响;戈壁针茅、芨芨草、柠条锦鸡儿(Caraganakorshinskii) 等植物主要分布在地形起伏小、砾石较少的地区,白皮锦鸡儿(Caraganaleucophloea)、戈壁藜、短叶假木贼(Anabasisbrevifolia)、木旋花(Convolvulusfruticosus)等植物等分布在地形起伏较大、地表覆盖粗糙的生境中。

表3 环境因子对植物群落影响的Monte Carlo检验及相对贡献率分析
***P<0.001,**P<0.01,*P<0.05
运用偏典范对应分析(pDCCA)分离环境变量对植物群落变异的解释能力,并将8个环境因子分为地形、气候、土壤3部分,来分析各环境因子对植物群落格局的影响[29]。由研究结果可知,地形(39.79%)是影响黑戈壁群落格局的主控因子,此外土壤(23.53%)、气候(15.08%)对群落格局也有着显著的影响;单个环境因子中,海拔(18.52%)、坡位(15.46%)、砾石盖度(15.1%)对群落的影响最明显,年平均年降水量(8.07%)、砾石大小(8.43%)、温度(7.01%)也对群落格局存在重要作用,此外未解释部分为48.1%,可能包括土壤化学性质、生物相互作用、干扰因素和随机因素等。
3.3植物群落物种多样性分布格局分析3.3.1黑戈壁植物物种多样性水平变化分析
从图4可以看出,多数样方内物种多样性指数都比较低,不同生境下样方多样性水平存在明显的差异。经向变化规律表现为多样性指数整体连续波动,变化剧烈,在92.6°E、93.3°E、93.9°E、96.1°E、97.5°E、100.5°E形成6个物种多样性高值区,在93.9E附近达到了最高值,而最低值则出现在100.9°E附近。从纬向变化上分析,多样性指数同样持续波动变化,在40.9°N、41.8°N、42°N、43.4°N、43.9°N、44.2°N这6个区域达到峰值,并在43.9°N达到最高值,在41.92°N、42.6°N附近出现了最低值区。

图4 物种多样性指数经度和纬度梯度变化Fig.4 Variation of species diversity index along latitude and longitude
整体而言,物种多样性随着经度与纬度的变化呈现为持续剧烈波动的复杂变化特征,在经向及纬向均没有明显的变化趋势。结合样方环境因子的数据分析,发现几个峰值出现的区域主要为西北部降水较多的天山南北的山前戈壁、中部海拔较高的肃北地区以及额济纳绿洲黑河流经的地区,水热条件的分配对物种多样性有着显著的影响。
3.3.2黑戈壁植物物种多样性垂直变化分析
黑戈壁地区海拔变化较大,调查的样方海拔在300—2400m之间。由于海拔导致的温度和降水的差异,使植被随着海拔的变化,形成不同的群落类型。由于地表粗糙度过大时,容易导致单优群落的出现,因此随着海拔的升高,物种多样性极高值和极低值同时存在,并在几个海拔区间上出现峰值;物种多样性在海拔300—1200m之间比较稳定,变化趋势趋于平坦,但当海拔增加到1400m以上时,物种多样性随着海拔的升高有着明显的增加趋势,并且与水平方向相比,垂直方向上物种多样性的变化趋势要更加明显。
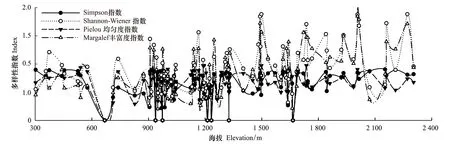
图5 物种多样性垂直梯度变化Fig.5 Variation of species diversity index along vertical gradient
3.3.3植物群落物种多样性空间异质性分析
从表4物种多样性半变异函数模型及方差分析结果可知,黑戈壁4种多样性指数除了均匀度指数为球面模型外,其他都更加符合指数模型,半变异函数模型能够很好地反映物种多样性指数的空间结构特性。在分析模型中块金值表征由于取样误差和小于取样尺度上随机部分引起的空间异质性,基台值越大,总的空间异质性越高;块金与基台的比值表示由系统变量引起的空间相关性,比值小于0.25表示空间相关性强烈,0.25—0.75表示中等的空间相关性,大于0.75表示空间相关性较弱[30];4种多样性指数比值都在0.25—0.75之间,因此研究区的物种多样性为中等的空间相关性,物种多样性呈斑块化分布。
同时也说明在大于研究尺度,小于变程的范围内结构性因子所占比例为0.673—0.742,随机性因子所占比例为0.258—0.327之间,说明由气候、海拔、地形、土壤理化条件等结构性因子引起的空间异质性占主导地位,而随机因子,如人为活动引起的空间异质性较弱。
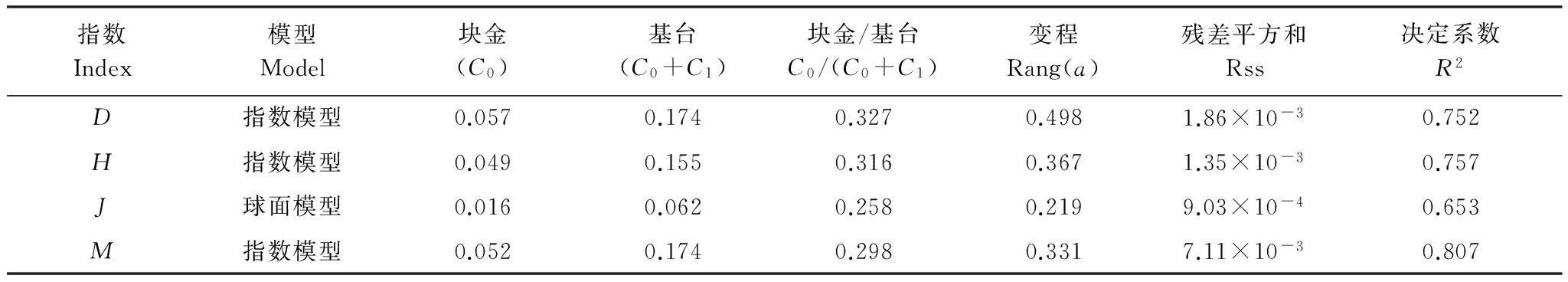
表4 物种多样性半变异函数模型及其相关参数
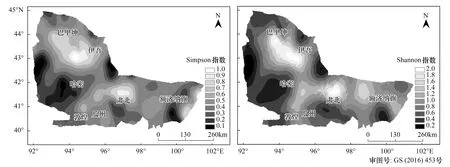
图6 黑戈壁物种多样性预测图Fig.6 Predicting maps of species diversity in Black Gobi
在半方差结构分析和球面模型的基础上,利用ArcGIS 10.2普通克里金插值法得到黑戈壁地区物种多样性预测图6,从中可以明显的看出黑戈壁地区的植物物种多样性的空间分布状况。物种多样性没有明显的经向和纬向变化规律,受大气活动路径以及地形对气候的影响,更多表现为自西北天山北麓向东和向南延伸的过程中不断减少的趋势,东天山北麓至北山及马鬃山迎风坡区域的物种多样最丰富,敦煌、哈密南部等降水极少地区,生物多样性水平普遍较低,表现出明显的斑块化分布特点。
4结论与讨论
4.1黑戈壁植物群落特征及其与环境因子的关系
黑戈壁地区地处我国的温带荒漠区,环境恶劣,土壤贫瘠,群落类型贫乏,物种生活型单一,主要以温带荒漠灌木和半灌木群落为主,并在部分区域形成温带灌丛和草甸。与其他荒漠区比较[11-12],在群落物种组成上,灌木、半灌木、小灌木所占比例更高,境内水土条件更加严苛,群落组成更为简单,优势群落优势度更高,群落组成物种通常为2—5个;建群种多为灌木、半灌木植物;与沙漠植被组成相比,则缺乏短命植物。
植被分布格局是不同尺度上包括气候、土壤、地形在在内多种环境因子共同影响的结果。区域尺度上,地带性气候(温度与降水)是决定植被类型分布及物种生活型的决定性因子,是地带性植被形成的基础[31- 32];而局部尺度上,包括微生境及土壤等非地带性环境因子决定着植被格局[31- 33],地形通过调节局部气候条件,进而控制地表的水热等因子,最终对群落的分布格局产生重要影响[34]。本研究结果证实了气候、土壤、地形是导致群落类型与组成产生差异的主要因素,研究区极端干旱的气候条件是戈壁荒漠植被形成的基础,而受到海拔等地形因子的影响,水土资源在局部尺度上的空间异质性是群落空间分异的主因,这与前人[34- 35]的研究结果一致;同时有48.1%的环境变量没有得到解释,表明群落分布还受到包括土壤化学性质、生物相互作用因素、干扰因素和随机因素等的影响[36]。
4.2黑戈壁植物群落物种多样性空间变化
黑戈壁地区物种多样性分布主要受到水热条件的控制;受天山、马鬃山等山脉的影响使得水热条件发生再分配,整体上出现了西北—东南走向的水分和热量的变化梯度[22],物种多样性没有明显的经向和纬向变化规律,更多表现为自西北天山北麓向东和向南延伸的过程中不断减少的趋势。我国内陆干旱区物种多样性多表现为中部隆起型的垂直梯度格局[30],黑戈壁地区物种多样性格局与其非常相似,在西部地区受到天山影响,海拔落差在2400m以上,而中部地区马鬃山等海拔也在2000m以上,东部的额济纳地区绿洲为发源于祁连山的黑河下游地区;研究区水热条件垂直分异明显,海拔对物种多样性有着重要的影响,海拔落差越大,这种影响越显著。
整体上,黑戈壁区不同群落间空间异质性强烈,为中等空间关联性,地理因素和海拔导致的水热条件和土壤条件等结构性因子异质性是群落产生空间异质性的主导因素;利用克里金插值得到的物种多样性预测图也说明了地理因素对植物物种多样有着重要影响。黑戈壁物种多样性的空间变异模式表明植物群落空间异质性强,呈现为斑块性分布,但总体上受到水热条件的控制。
4.3黑戈壁植物群落及其物种多样性维持的一般机理
关于干旱地区植被维持的问题,不同学者观点也不一致。目前,主要的观点包括空间异质性影响、空间自组织等,或者兼而有之[31-35,37-40]。Wesche通过对戈壁地区植被分布研究提出[16],水资源的可利用性是决定植被存在与分布的最关键因素。地形可能对于水热条件再分配起到重要的作用,在植被格局形成过程中,植物对有限资源的竞争和利用被认为是最重要的驱动力[41]。而物种在竞争过程中所形成的生长、繁殖以及扩散能力的分化对于物种的共存是非常重要的[40-42]。在黑戈壁地区,植物群落组成主要以灌木和半灌木植物为主,并且密集型的克隆生长比较普遍,植物有性繁殖方式也很特殊甚至特化[12,22]。目前的研究也表明:植物克隆生长和不同繁殖特性对于个体生存以及群落动态有着重要的作用[43]。戈壁植物群落格局及其物种多样性与水文过程以及戈壁植物特殊的生长与繁殖适应性等因素有关,但戈壁植物群落格局及物种多样性维持机理与过程具体是怎样的,这些问题还需要深入研究来解决。
参考文献(References):
[1]Levine J M, HilleRisLambers J. The importance of niches for the maintenance of species diversity. Nature, 2009, 461(7261): 254- 257.
[2]Thibaut L M, Connolly S R. Understanding diversity-stability relationships: towards a unified model of portfolio effects. Ecology Letters, 2013, 16(2): 140- 150.
[3]Burke A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. Journal of Vegetation Science, 2001, 12(1): 53- 60.
[4]张雪妮, 吕光辉, 王庭权, 马玉, 阿布里孜·阿不都热合, 赵晓英, 郭振洁, 朱修逸. 荒漠区垂直河岸带植物多样性格局及其成因. 生态学报, 2015, 35(18): 5966- 5974.
[5]张锦春, 王继和, 赵明, 刘虎俊, 廖空太, 徐先英. 库姆塔格沙漠南缘荒漠植物群落多样性分析. 植物生态学报, 2006, 30(3): 375- 382.
[6]袁蕾, 周华荣, 宗召磊, 叶琴. 乌鲁木齐地区典型灌木群落结构特征及其多样性研究. 西北植物学报, 2014, 34(3): 595- 603.
[7]孟莹莹, 周莉, 周旺明, 于大炮, 包也, 王晓雨, 郭焱, 代力民. 长白山风倒区植被恢复26年后物种多样性变化特征. 生态学报, 2015, 35(1): 142- 149.
[8]Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220- 227.
[9]陈昌笃, 张立运. 中国的极旱荒漠. 干旱区资源与环境, 1987, 1(3/4): 1- 12.
[10]吴正. 中国沙漠及其治理. 北京: 科学出版社, 2009.
[11]陈鹏, 潘晓玲. 河西走廊地区植物的区系特征. 植物研究, 2001, 21(1): 24- 30.
[12]夏延国, 宁宇, 李景文, 李俊清, 冯益民, 吴波, 卢琦. 中国黑戈壁地区植物区系及其物种多样性研究. 西北植物学报, 2013, 33(9): 1906- 1915.
[13]朱军涛, 于静洁, 王平, 王志勇. 额济纳荒漠绿洲植物群落的数量分类及其与地下水环境的关系分析. 植物生态学报, 2011, 35(5): 480- 489.
[14]鱼腾飞, 冯起, 司建华, 席海洋, 陈丽娟. 黑河下游额济纳绿洲植物群落物种多样性的空间异质性. 应用生态学报, 2011, 22(8): 1961- 1966.
[15]Pfeiffer M, Chimedregzen L, Ulykpan K. Community organization and species richness of ants (Hymenoptera/Formicidae) in Mongolia along an ecological gradient from steppe to Gobi desert. Journal of Biogeography, 2003, 30(12): 1921- 1935.
[16]Wesche K, Miehe S, Miehe G. Plant communities of the Gobi Gurvan Sayhan National park (South Gobi Aymak, Mongolia). Candollea, 2005, 60(1): 149- 205.
[17]Von Wehrden H, Wesche K. Relationships between climate, productivity and vegetation in southern Mongolian drylands. Basic and Applied Dryland Research, 2007, 1(2): 100- 120.
[18]Wesche K, von Wehrden H. Surveying Southern Mongolia: application of multivariate classification methods in drylands with low diversity and long floristic gradients. Applied Vegetation Science, 2011, 14(4): 561- 570.
[19]Sasaki T, Okayasu T, Takeuchi K, Jamsran U, Jadambaa S. Patterns of floristic composition under different grazing intensities in Bulgan, South Gobi, Mongolia. Grassland Science, 2005, 51(3): 235- 242.
[20]Okayasu T, Muto M, Jamsran U, Takeuchi K. Spatially heterogeneous impacts on rangeland after social system change in Mongolia. Land Degradation & Development, 2007, 18(5): 555- 566.
[21]Sugimoto N, Hara Y, Yumimoto K, Uno I, Nishikawa M, Dulam J. Dust emission estimated with an assimilated dust transport model using lidar network data and vegetation growth in the Gobi desert in Mongolia. SOLA, 2010, 6: 125- 128.
[22]中国黑戈壁地区生态本底科学考察队. 中国黑戈壁研究. 北京: 科学出版社, 2014.
[23]吴征镒. 中国植被. 北京: 科学出版社, 1980.
[24]Jennings M D, Faber-Langendoen D, Loucks O L, Peet R K, Roberts D. Standards for associations and alliances of the U.S. National Vegetation Classification. Ecological Monographs, 2009, 79(2): 173- 199.
[25]张金屯. 数量生态学. 北京: 科学出版社, 2004.
[27]曹静, 苗艳明, 冯飞, 许强, 张钦弟, 毕润成. 稀有种不同处理对典范对应分析排序结果影响的比较. 植物生态学报, 2015, 39(2): 167- 175.
[28]ter Braak C J F,milauer P. CANOCO Reference Manual and CanoDraw for Windows User′s Guide: Software for Canonical Community Ordination (Ver. 4.5). New York: Microcomputer Power, Ithaca, 2002.
[29]Borcard D, Legendre P, Drapeau P. Partialling out the spatial component of ecological variation. Ecology, 1992, 73(3): 1045- 1055.
[30]马斌, 周志宇, 张莉丽, 高文星, 陈善科, 张宝林. 阿拉善左旗植物物种多样性空间分布特征. 生态学报, 2008, 28(12): 6099- 6106.
[31]Van Couwenberghe R, Collet C, Lacombe E, Pierrat J C, Gégout J C. Gap partitioning among temperate tree species across a regional soil gradient in windstorm-disturbed forests. Forest Ecology and Management, 2010, 260(1): 146- 154.
[32]Siefert A, Ravenscroft C, Althoff D, Alvarez-Yépiz J C, Carter B E, Glennon K L, Heberling J M, Jo I S, Pontes A, Sauer A, Willis A, Fridley J D. Scale dependence of vegetation-environment relationships: a meta‐analysis of multivariate data. Journal of Vegetation Science, 2012, 23(5): 942- 951.
[33]李国庆, 王孝安, 郭华, 朱志红. 陕西子午岭生态因素对植物群落的影响. 生态学报, 2008, 28(6): 2463- 2471.
[34]余敏, 周志勇, 康峰峰, 欧阳帅, 米湘成, 孙建新. 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释. 植物生态学报, 2013, 37(5): 373- 383.
[35]沈泽昊, 张新时, 金义兴. 地形对亚热带山地景观尺度植被格局影响的梯度分析. 植物生态学报, 2000, 24(4): 430- 435.
[36]赵从举, 康慕谊, 雷加强. 准噶尔盆地典型地段植物群落及其与环境因子的关系. 生态学报, 2011, 31(10): 2669- 2677.
[37]Couteron P, Lejeune O. Periodic spotted patterns in semi-arid vegetation explained by a propagation inhibition model. Journal of Ecology, 2001, 89(4): 618- 628.
[38]Van de Koppel J, Rietkerk M, Van Langevelde F, Kumar L, Klausmeier C A, Fryxell J M, Hearne J W, van Andel J, de Ridder N, Skidmore A, Stroosnijder L, Prins H T P. Spatial heterogeneity and irreversible vegetation change in semi-arid Grazing systems. The American Naturalist, 2002, 159(2): 209- 218.
[39]Rietkerk M, Ouedraogo T, Kumar L, Sanou S, van Langevelde F, Kiema A, van de Koppel J, van Andel J, Hearne J, Skidmore A, de Ridder N, Stroosnijder L, Prins H H T. Fine-scale spatial distribution of plants and resources on a sandy soil in the Sahel. Plant and Soil, 2002, 239(1): 69- 77.
[40]HilleRisLambers R, Rietkerk M, Van Den Bosch F, Prins H H T, de Kroon H. Vegetation pattern formation in semi-arid grazing systems. Ecology, 2001, 82(1): 50- 61.
[41]Ryabov A B, Blasius B. A graphical theory of competition on spatial resource gradients. Ecology Letters, 2011, 14(3): 220- 228.
[42]RietKerk M, Boerlijst M C, van Langevelde F, HilleRisLambers R, Van De Koppel J, Kumar L, Prins H H T, De Roos A M. Self-Organization of vegetation in arid ecosystems. The American Naturalist, 2002, 160(4): 524- 530.
[43]Charpentier A. Consequences of clonal growth for plant mating. Evolutionary Ecology, 2002, 15(4/6): 521- 530.
基金项目:林业行业公益项目(201404304);中国林业科学研究院重大项目 (CAFYBB2011002);国家自然科学基金资助项目(31570610)
收稿日期:2015- 06- 10;
修订日期:2016- 01- 25
*通讯作者
Corresponding author.E-mail: lijingwen@bjfu.edu.cn
DOI:10.5846/stxb201506101176
Plant distribution patterns and the factors influencing plant diversity in the Black Gobi Desert of China
WANG Jianming1, DONG Fangyu1, BahaiNasina1,3, LI Jingwen1,*,LI Junqing1,FENG Yiming2, LU Qi2
1TheCollegeofBeijingForestryUniversity,Beijing100083,China2InstituteofDesertificationStudies,ChineseAcademyofForestry,Beijing100091,China3XinjiangKeyForestryEngineeringQualityManagementStation,Urumchi830000,China
Abstract:The Black Gobi Desert covers a large area in the northwest of China, with a total area of about 200,000 km2. The general climate of the Black Gobi can be characterized as having a high evaporative demand, with variable but low precipitation, making it the most arid region of the country′s dryland areas. Nevertheless, despite being considered the most dry and bare ecosystem in the world, it is affluent in natural resources. Because of its harsh environment and poor accessibility, systematic studies of plant diversity in the Black Gobi Desert are lacking. Understanding the underlying causes of geographic plant diversity gradients in arid and semiarid regions is important for the conservation and sustainable use of biodiversity there. However, the underlying mechanisms behind the Black Gobi Desert′s plant diversity patterns are still elusive. Therefore, the aim of our study was to analyze the plant community composition, as well as the species diversity of the Black Gobi ecosystem. We sampled 174 plots across the Black Gobi to demonstrate the vegetation pattern and its relationship with the environmental factors. We observed a trend of the plant communities becoming poorer and simpler in composition while vegetation cover decreases. Moreover, the overall structure and composition of the plant communities were simple, the majority of plants having an importance value lower than 0.1. The plant communities consisted mainly of Haloxylon ammodendron, Reaumuria songarica, Nitraria tangutorum, and 10 other species. Species diversity showed non-significant latitudinal and longitudinal trends, but was significantly influenced by climate, elevation, and topography. In addition, structural factors were most important in determining the spatial variation of species diversity. The result of a detrended cross-correlations analysis (DCCA) indicated that climate, elevation, topography, soil texture, and surface cover had a significant effect on community composition and plant distribution. Overall, the vegetation spanned a gradient in elevation,soil texture and water availability. Plant species are limited by water availability, and it should be noted that plant community composition was strongly related to changes in elevation, which influence soil texture and water availability. In sum, the spatial heterogeneity of the soil texture and water availability were the main factors to determine plant distribution and species diversity.
Key Words:Black Gobi; plant communities; species diversity; plant distribution; spatial heterogeneity
Wang J M, Dong F Y, BahaiNasina, Li J W,Li J Q,Feng Y M, Lu Q.Plant distribution patterns and the factors influencing plant diversity in the Black Gobi Desert of China.Acta Ecologica Sinica,2016,36(12):3488- 3498.
猜你喜欢
中国医药导报(2016年28期)2017-01-06 19:25:52
中国当代医药(2016年29期)2017-01-03 15:20:06
绿色科技(2016年21期)2016-12-27 17:06:10
绿色科技(2016年21期)2016-12-27 10:53:26
农业研究与应用(2016年1期)2016-12-26 12:35:42
现代农业科技(2016年20期)2016-12-20 09:17:04
科技创新导报(2016年21期)2016-12-17 12:53:50
农业与技术(2016年15期)2016-11-09 16:58:30
现代经济信息(2016年13期)2016-06-17 21:16:45
中国科技博览(2016年9期)2016-04-25 10:49:09
