
福州市绿地生态系统光合特性及土壤呼吸研究
2016-07-25 02:13郁珊珊王浩王亚军
江苏农业科学 2016年6期
关键词:福州市
郁珊珊+王浩+王亚军



摘要:为了探寻福州市不同绿地生态系统(草地、林地、耕地和园地)光合特征和土壤呼吸变化规律及其影响因素,利用指数模型和相关系数分析土壤呼吸与土壤温度、湿度和光合特征的关系。结果表明:(1)4种绿地生态系统植物LSP、LCP、AQY、CE、Ls和WUE基本表现出一致的变化规律,综合表现为林地和耕地显著高于草地和园地;(2)1 d中各时期净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为林地和耕地显著高于草地和园地,4种绿地生态系统Pn和EVAP日变化均呈双峰曲线,气孔限制导致了光合“午休”现象,Ci和Gs日变化随时间呈“W”字形变化规律,通过SPSS最佳曲线拟合4种绿地生态系统植物的Pn与PAR、CO2浓度和温度变化呈一致的二次曲线关系;(3)在日的时间尺度上,4种绿地生态系统土壤呼吸均表现为单峰型,且峰值出现的时间基本一致,约在14:00达到最大,最低值出现在06:00,相同时间基本表现为林地和耕地显著高于草地和园地;(4)4种绿地生态系统土壤呼吸与土壤温度和湿度之间关系以指数方程拟合最好,通过偏相关分析可知,草地、林地、耕地和园地土壤呼吸均与Pn、EVAP和WUE呈显著或者极显著的相关关系,而与其他光合生理指标基本没有显著相关关系(P>005),由此表明土壤温度、土壤湿度、Pn、EVAP和WUE是影响福州市4种绿地生态系统土壤呼吸的主要环境因子。
关键词:福州市;绿地生态系统;光和特性;土壤呼吸
中图分类号: S181.6文献标志码: A文章编号:1002-1302(2016)06-0483-05
收稿日期:2016-01-25
项目基金:国家自然科学基金(编号:2014BC210);福建省厦门市科技专项课题(编号:14020041)。
作者简介:郁珊珊(1981—),女,福建厦门人,博士,副教授,研究方向为风景园林规划与设计、城市绿地生态系统规划。E-mail:shanshan_yu81@163.com
通信作者:王浩,博士,教授,博士生导师,研究方向为风景园林规划与设计。城市绿地生态系统是由各类城市绿地(草地、林地、耕地、公园绿地等)相互联系组合的城市绿色环境体系[1-2]。具有自净能力、自动调节能力和生命力,在改善城市生态环境、美化市容、调节城市生态平衡中发挥着主导作用,已成为衡量城市生态文明和城市现代化的重要标志[3]。城市绿地可以通过植物的蒸腾、蒸散、吸收、吸附、反射等功能,降低温度,增加湿度,固碳释氧,抗污染(吸收粉尘、Cl2、SO2、CO等),降低噪音,保护生物多样性等[4]。随着生态城市概念的提出、建设和发展,人们日益注意到城市绿地的生态意义和环境价值[5],而快速工业化及城市化给城市绿地的生长环境造成了巨大的胁迫效应,加速了城市的“生态环境危机”[6-7]。
光照是城市绿地植物生命活动中起重大作用的生存因子,也是对城市绿地植物光合结构重要的环境因素,城市绿地植物需要在一定的光照条件下完成生长发育过程,但不同城市绿地植物对光照强度的适应范围亦有所差别[8-9]。土壤作为大气CO2重要源或汇,土壤CO2细微的改变都会显著改变大气中CO2的浓度和碳的累积速率[10]。在陆地生态系统中,土壤碳库是大气碳贮量的2~3倍,通过土壤呼吸作用向大气释放的CO2约占全球CO2交换量的25%[11]。同时,土壤呼吸受多种因素的共同交互影响,包括土壤温度、湿度、土壤养分、净生态系统生产力(NEP)和人类干扰等,通过测定城市绿地生态系统土壤呼吸速率特征并阐明影响土壤呼吸、植物光合相互作用关系及其环境因子,不仅有利于改变人们对城市绿化植被的认识,更有利于科学地选取和配置绿地绿化植物,使其最大化发挥其生态效益,从而促进城市绿地系统规划、优化植物群落结构、改善环境质量、促进城市生态建设[12-13]。因此,本研究采用美国Li-6400便携式气体分析系统和 Li-6400 土壤呼吸室测量福建省福州市绿地生态系统植物光合特性、土壤呼吸速率及影响因子,以期深入了解植物光合特性和土壤呼吸作用的变化过程及变化机理,从而为城市绿地系统建设中植物的选择和应用提供科学依据。
1材料与方法
1.1研究区概况
福州市位于福建省东部沿海闽江下游(25°15′~26°39′N,118°08′~120°31′E),气候资源丰富、温暖湿润、四季常青、雨量充沛、霜少无雪、多低山丘陵,农业活动对土壤的影响深刻,水文条件各不相同,地貌类型以丘陵为主,地形复杂多样,地势起伏不平,土壤类型较多,主要土类有黄棕壤、红壤、石灰岩土、紫色土、潮土和沼泽土,平均海拔9.2 m,无霜期达 326 d,年均日照数为1 700~1 980 h,年均降水量为900~2 100 mm,年均气温为16~20 ℃,年相对湿度约77%。福州市土地总面积为12 251 km2,其中林地面积7 032 km2,占土地总面积的57.4%,绿地面积81.2 km2,绿地率达36.9%,绿化覆盖面积88.7 km2,绿化覆盖率高达40.3%,人均公园绿地高达11.2 m2。曾荣获“全国文明城市”“全国绿化模范城市”“全国城乡绿化先进集体”“国家卫生城市”和“中国优秀旅游城市”等光荣称号[14]。
1.2测定方法
1.2.1土壤呼吸的测定选择保护区内落叶季雨林和常绿季雨林,每个林分布设2个监测点,相距80 m,在第1次测定土壤呼吸之前,提前1 d将测定基座(soil collar)嵌入土壤中,在每个基座内存留的地表植被自土壤表层彻底剪除,聚氯乙烯圆柱体经过24 h的平衡后,土壤呼吸速率会恢复到基座放置前的水平,从而避免了因安置气室对土壤扰动而造成的短期内呼吸速率波动。采用动态密闭气室红外CO2分析仪(IRGA)法,测定仪器型号为美国Li-6400便携式气体分析系统和Li-6 400土壤呼吸室,土壤呼吸日变化测定为白天每隔2 h测定1次,每个监测点1次测定2个重复,取平均值。季节变化测定为2015年7月中旬,在08:00—10:00之间测量,有报道指出该时间段测定值最为接近24 h平均值,具有较好的代表性。在观测土壤呼吸的同时,使用手持长杆电子温度探针(SK-250 WP,Sato Keiryoki,Kanda,Japan)测定0~10 cm深处的土壤温度,使用时域反射仪(Model TDR 300,Spectrum,Aurora,USA)测定每个监测点附近0~10 cm范围内的土壤湿度,每个样地温湿度安装数据采集器(HOBO ProRH/TEMP Data Logger),以获取距地面50 cm高度的大气温度和相对湿度数据,用于分析土壤呼吸速率和水热因子的关系。
1.2.2光合特性测定(1)美国LI-COR公司生产的LI-6400系列便携式光合作用测量系统进行气体交换光合生理特征测定,2015年6月下旬07:00—19:00(晴天无风),每隔 2 h 测定1次,连续重复测定3 d,每个处理选择3株健康油茶重复测定,每株选取当年生的成熟叶片(从植株顶部数第3~6片完全展开的成熟叶),轮流测定以消除测定时间上的误差。红蓝光源光照设定为1 000 μmol/(m2·s),温度30 ℃,CO2流量为400 μmol/s,不同测定日期温度变化区间为22.1~35.4 ℃,辐射强度(PAR)为112~1 371 μmol/(m2·s),空气相对湿度在24.6%~45.7%之间,选取充分受光、叶位一致的叶片(上、中、下当年生成熟叶)进行测定,轮流测定以消除测定时间上的误差。仪器记录净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)、胞间CO2浓度(Ci)、环境CO2浓度(Ca)和光照强度(PAR),其中PAR和CO2浓度以外界条件为准。
(2)光合-光强响应特性的测定,测定时CO2摩尔分数控制为350 μmol/mol,温度为25 ℃,湿度约为大气湿度的99%。设定光照度梯度为1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、0 μmol/(m2·s),每个光照度下适应 5 min 后测定Pn值。根据Pn-PAR回归方程求出各品种的光补偿点(LCP)、光饱合点(LSP);表观量子效率(AQY)根据低于200 μmol/(m2·s)光照度下测定的叶片PFD-Pn曲线计算求得;CO2羧化效率(CE)根据CO2 250 μmol/(m2·s)下测定的叶片CO2-Pn曲线计算求得[15]。
(3)气孔限制值(Ls)和叶片瞬时水分利用效率(WUE)由公式计算,即Ls=1-Ci/Ca,WUE=Pn/EVAP[16]。
1.3数据处理与分析
采用Excel 2007和SPSS 18.0进行数据分析,单因素方差分析(One-way ANOVA)和最小显著法(LSD)检验其差异显著性,Pearson相关性系数检验各指标的相关性,采用指数方程拟合土壤呼吸与土壤温度和湿度的响应关系,由原始数据拟合得到的多元回归关系经统计学检验得到拟合度参数R2和Radj2,并在P<0.01水平检验相关系数的显著性。
2结果与分析
2.1绿地生态系统植物光合特性
2.1.1绿地生态系统植物叶片生理特性由图1可知,福州市绿地生态系统植物光饱和点(LSP)变化范围为658.9~1 235.8 μmol/(m2·s),依次表现为林地>耕地>草地>园地,4种绿地LSP差异均显著(P<0.05);光补偿点(LCP)变化范围为74.3~125.3 μmol/(m2·s),依次表现为林地>耕地>草地>园地,其中草地和园地差异不显著(P>0.05),二者显著低于林地和耕地(P<0.05);表观量子效率(AQY)变化范围为18.9×10-3~33.5×10-3,依次表现为林地>耕地>园地>草地,4种绿地AQY差异均不显著(P>0.05);羧化效率(CE)变化范围为32.7×10-3~69.7×10-3,依次表现为耕地>林地>草地>园地,其中园地显著低于其他绿地
(P<0.05);气孔限制值(Ls)变化范围为0.51~0.93 μmol/(m2·s),依次表现为耕地>林地>草地>园地,其中耕地与林地之间差异并不显著(P>0.05),二者显著高于草地和园地(P<0.05);水分利用效率(WUE)变化范围为2.59~6.58 μmol/(m2·s),依次表现为林地>耕地>草地>园地,4种绿地WUE差异均显著(P<0.05)。
2.1.2绿地生态系统植物叶片光合特性福州市绿地生态系统植物叶片光合特性见图2,4种绿地生态系统植物叶片光合指标在1 d之中的变化趋势基本保持一致,1 d中各时期净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为林地和耕地>草地和园地,局部有所波动;绿地生态系统植物叶片光合速率(Pn)日变化均表现为典型的双峰型曲线,第1峰出现在约10:00,12:00—13:00光合速率出现峰谷,第二高峰出现在约16:00,但明显小于第1峰值,16:00以后光合速率明显下降,表现出明显的光合“午休”现象;胞间CO2浓度(Ci)日变化随时间呈“W”字形变化规律,在约10:00出现第1个波谷,12:00达到最大(峰值),12:00 以后急剧下降,约16:00出现第2峰谷,第2波谷值要高于第1波谷值,而后随时间的变化出现缓慢上升趋势;气孔导度(Gs)日变化随时间呈“W”字形变化规律,其第1波峰出现在08:00,在10:00出现第1个波谷,第2波峰出现在13:00,但第2波峰明显低于第1波峰,16:00出现第1次峰谷,而后随时间的变化缓慢上升;蒸腾速率(EVAP)日变化规律与光合速率(Pn)保持一致,均呈双峰曲线,局部峰值推迟或者提前。
2.1.3绿地生态系统植物叶片光合特性与生理响应由表1可知,通过SPSS最佳曲线拟合(分别拟合光合速率Pn与PAR、CO2浓度和温度之间的曲线关系),筛选出R2最大(最佳拟合度及校准以后的最大R2)和P值(显著性)最小的曲线组合,并在此基础上检验其F值和差异显著性(P<0001)。福州市绿地生态系统植物的Pn-PAR呈一致的二次曲线关系,曲线拟合方程见表3。
2.2绿地生态系统土壤呼吸特性
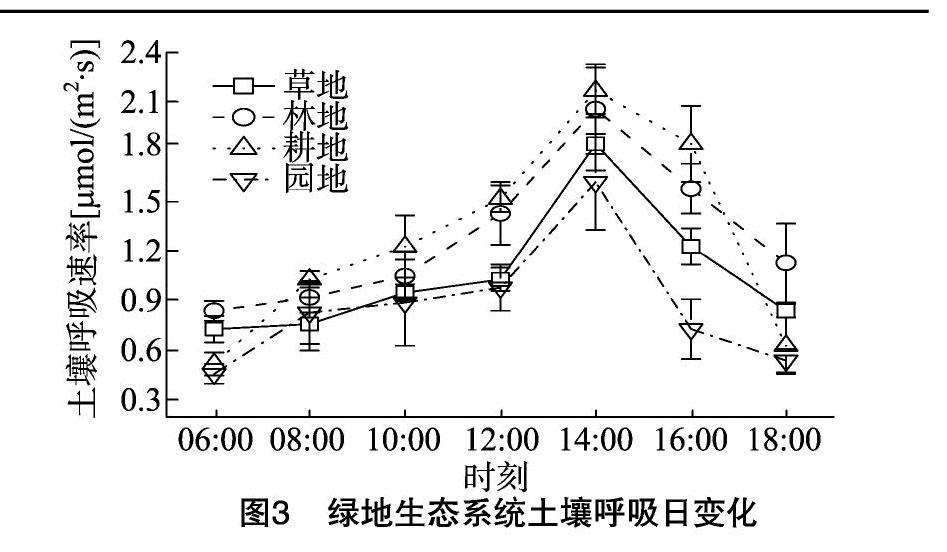
2.2.1绿地生态系统土壤呼吸日变化由图3可知,4种绿地生态系统土壤呼吸存在明显的日变化规律, 均表现为单峰型,且峰值出现的时间基本一致,土壤呼吸速率都在约14:00达到最大,最低值出现在06:00,在06:00—10:00,4种绿地生态系统土壤呼吸升高缓慢,12:00以后急剧上升,达到最大值以后急剧降低。草地土壤呼吸日变化范围为0.73~1.85 μmol/(m2·s),林地土壤呼吸日变化范围为0.84~2.06 μmol/(m2·s),耕地土壤呼吸日变化范围为0.52~217 μmol/(m2·s),园地土壤呼吸日变化范围为0.46~162 μmol/(m2·s)。相同时间土壤呼吸速率基本表现为耕地>林地>草地>园地。
2.2.2土壤温度和湿度对土壤呼吸的影响为进一步探讨绿地生态系统土壤呼吸与其显著影响因子土壤温度和湿度的关系,国内外学者一般采用线性模型、二次方程、指数模型等多种方法进行拟合,本试验结果中将土壤呼吸强度与土壤 10 cm 温度进行曲线拟合,筛选出最佳拟合度的方程(R2最大,P值最小),得出的不同拟合方程;由表4可知,土壤呼吸与土壤温度之间关系以指数方程拟合最好。4种绿地生态系统土壤呼吸与土壤温度的指数关系达到了极显著水平(P<0.01),且指数模型的决定系数最大,故指数模型的拟合效果最好。4种绿地生态系统土壤呼吸与土壤湿度之间关系也以指数方程拟合最好。
2.3土壤呼吸与光合的相关关系
由绿地生态系统土壤呼吸与植物叶片光合特征参数标准化经SPSS双变量Pearson相关系数法分析的结果(表5)可知,土壤呼吸主要是植物的根以及土壤微生物的呼吸,植物的生长、微生物的氧化分解等活动都要受周围环境如土壤的温度、土壤的水分、气温、空气湿度等的影响,因此,这些环境因子也是影响土壤呼吸强度的重要因素。由表5可知,4种绿地生态系统土壤呼吸强度与各环境因子的Pearson相关关系均显著,但有时由于某一共同变量的干扰会导致2个变量间的假相关,为排除其他因素的干扰,对各因子分别作偏相关分析,控制其他几个变量以分析土壤呼吸与另一个变量的净相关关系。结果表明,草地、林地、耕地和园地土壤呼吸均与Pn、EVAP和WUE呈显著(P<0.05)或极显著(P<0.01)的相关关系,而与其他光合生理指标基本没有显著相关关系(P>0.05)。
3讨论
3.1绿地生态系统光合特性特征分析
光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)、羧化效率(CE)、气孔限制值(Ls)和水分利用效率(WUE)均能衡量植物对强光或弱光的利用能力。本研究中4种绿地生态系统植物LSP、LCP、AQY、CE、Ls和WUE基本表现出一致的变化规律,综合比较可知林地和耕地显著高于草地和园地,主要是受植被类型及植物本身特性的影响,林地和耕地植物叶片对低CO2浓度和低光强的利用率最强,具有较强的再生速率。此外,本研究中光合生理指标属于瞬间测定的结果,受环境影响较大,所测定的结果有一定的偏差,后续还需要进行反复大量的野外监测来验证本试验的研究结果。
植物的光合特性反映了对外界环境的响应和适应,受到众多环境因素的影响,其中影响光合速率的因素主要是气孔限制和非气孔限制因素,即气孔的开张和叶肉细胞的光合活性[17]。4种绿地生态系统Pn日变化表现为双峰曲线,有明显的“午休”现象,气孔导度(Gs)和胞间CO2浓度(Ci)变化方向与净光合速率(Pn)变化方向相反,1 d之中相同时间段净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和空气中CO2浓度基本表现为林地和耕地显著高于草地和园地,主要是由于林地和耕地(灌木乔木为主)比草地和园地(草本为主)能够利用更多的时间合成有机物而增产,中午持续的强光使气温、蒸腾速率急剧增加,叶片水分代谢失调,叶面温度过高而抑制了参与光合过程酶的活性,气孔导度(Gs)、胞间CO2浓度(Ci)和空气中CO2浓度导致Pn下降,从而造成了光合“午休”现象[18-19]。气孔是水分与叶片进行气体交换的通道,也是CO2进入细胞的门户,控制着叶片和大气之间的CO2和水分交换,气孔的闭合程度对光合与蒸腾作用会产生直接的影响,本研究中4种绿地生态系统气孔导度(Gs)日变化与蒸腾速率(EVAP)曲线相似,说明气孔的开闭程度直接影响蒸腾作用,主要是由水蒸气浓度差及水蒸气扩散阻力所决定[20];EVAP与Gs变化趋势一致,主要是由于叶内外水蒸气压差的增大和水蒸气浓度减少,加剧了植物体蒸腾扩散阻力,从生理生态的角度看,光合和蒸腾分别是CO2和H2O通过叶片气孔的内外物质交换过程,气孔行为对光合与蒸腾将产生直接影响,叶内外水蒸气压差的增大和水蒸气浓度减少,加剧了植物体蒸腾扩散阻力[21-22];本研究中在绝大多数时段Ci和Gs变化方向与其Pn变化方向相反,气孔主导其光合速率的变化,光合速率绝大多数取决于气孔限制因素,即叶肉细胞光合能力的变化主导其光合速率的变化。同时,绿地生态系统植物也能采取不同的适应策略应对环境的变化,叶片水分饱和亏缺逐渐增加的同时,净光合速率和蒸腾速率逐渐减少,光合限制因素由气孔限制为主逐渐转变为非气孔限制为主[23]。此外,水分状况也是影响小麦光合作用最重要的因子之一,4种绿地生态系统具有较高的EVAP,说明其需水量较多,因此要加强对水分的管理。
通过SPSS最佳曲线拟合(分别拟合光合速率Pn与PAR、CO2浓度和温度之间的曲线关系),福州市4种绿地生态系统植物的Pn与PAR、CO2浓度和温度变化呈一致的二次曲线关系,由曲线计算可知绿地生态系统植物光合作用最适温度为23~25 ℃,这与前人的研究结论[20-21]一致。研究还发现,虽然绿地生态系统植物光合适温相差不大,但在最适温度时的光合速率却相差较大,说明温度的变化对4种绿地生态系统植物光合作用有较大的影响。不同植物的光合作用对温度的响应不同,可能与物种的温度适应性(原产地的气候、环境)有关。
3.2绿地生态系统土壤呼吸特征分析
福州市4种绿地生态系统大气温度存在显著的日变化,气温的日较差在早、晚可达10 ℃以上,白天较高的温度有利于植物进行光合作用,下午较低的温度下植物呼吸作用渐弱,有利于植物进行有机物质的积累,大气温度变化幅度大,14:00 温度达到最高,土壤呼吸速率对地表温度响应较快,因此土壤呼吸速率在此时段达到峰值。从4种绿地生态系统土壤呼吸的日变化分布特征可知,在白天林冠的调节作用趋向于在昼间热量充足时从外界获取能量,夜间则趋向于保持环境稳定,将林内外气温保持在一个可控的范围内。大量研究结果显示,森林、草原以及湿地等的土壤呼吸速率呈现单峰型特点,本研究中绿地生态系统土壤呼吸日和季节变化同样呈现明显的单峰曲线,日变化峰值出现在14:00,最小值约出现在06:00,此时段地表温度较低、湿度较大,此外,根系活动和呼吸作用较弱,因此土壤呼吸速率低,这与前人的研究结果[24-25]相一致。
土壤呼吸速率主要由温度、湿度、环境因子等共同作用所驱动,控制因子随生态系统类型和气候类型变化而变化。对于城市绿地生态系统,水热环境因子是影响其土壤呼吸最为主要的两大因素,土壤湿度在一定程度上降低土壤呼吸速率对土壤温度的响应,土壤湿度的限制作用可能是导致土壤呼吸速率的敏感性降低[24,26]。因此,土壤温度、湿度和养分含量相互作用影响着绿地生态系统土壤呼吸,同时,在本研究中坡度和坡向的变化也影响着绿地生态系统土壤呼吸。本研究中4种绿地生态系统土壤呼吸作用受土壤湿度和温度的影响较为明显,土壤呼吸速率与土壤(0~10 cm)湿度呈负指数相关关系,与土壤(0~10 cm)温度呈正指数相关关系,土壤温度和土壤湿度是影响福州市4种绿地生态系统土壤呼吸的重要环境因子。相关性分析可知,草地、林地、耕地和园地土壤呼吸均与Pn、EVAP和WUE呈显著(P<0.05)或者极显著(P<0.01)的相关关系,而与其他光合生理指标基本没有显著相关关系(P>0.05),由此表明土壤温度、土壤湿度、Pn、EVAP和WUE是影响福州市4种绿地生态系统土壤呼吸的主要环境因子。
参考文献:
[1]李锋,王如松. 城市绿地系统的生态服务功能评价、规划与预测研究——以扬州市为例[J]. 生态学报,2003,23(9):1929-1936.
[2]张浩,王祥荣. 城市绿地的三维生态特征及其生态功能[J]. 中国环境科学,2001,21(2):101-104.
[3]周廷刚,郭达志. 基于GIS的城市绿地景观空间结构研究——以宁波市为例[J]. 生态学报,2003,23(5):901-907.
[4]Harawa N T,Greenland S,Bingham T A,et al. Associations of race/ethnicity with HIV prevalence and HIV-related behaviors among young men who have sex with men in 7 urban centers in the United States[J]. Journal of Acquired Immune Deficiency Syndromes,2004,35(5):526-536.
[5]Tajima K. New estimates of the demand for urban green space:implications for valuing the environmental benefits of Bostons big dig project[J]. Journal of Urban Affairs,2003,25(5):641-655.
[6]何兴元,陈玮,徐文铎,等. 沈阳城区绿地生态系统景观结构与异质性分析[J]. 应用生态学报,2003,14(12):2085-2089.
[7]蔺银鼎,韩学孟,武小刚,等. 城市绿地空间结构对绿地生态场的影响[J]. 生态学报,2006,26(10):3339-3346.
[8]Erickson R A,Jimenez R. Microfluidic cytometer for high-throughput measurement of photosynthetic characteristics and lipid accumulation in individual algal cells[J]. Lab on a Chip,2013,13(15):2893-2901.
[9]Yuan L,Shu S,Sun J,et al. Effects of 24-epibrassinolide on the photosynthetic characteristics,antioxidant system,and chloroplast ultrastructure in Cucumis sativus L. under Ca(NO3)2 stress[J]. Photosynthesis Research,2012,112(3):205-214.
[10]Gomez-Casanovas N,Matamala R,Cook D R. Net ecosystem exchange modifies the relationship between the autotrophic and heterotrophic components of soil respiration with abiotic factors in prairie grasslands[J]. Global Change Biology,2012,18(8):2532-2545.
[11]Reid J P,Adair E C,Hobbie S E,et al. Biodiversity,nitrogen deposition,and CO2 affect grassland soil carbon cycling but not storage[J]. Ecosystems,2012,15(4):580-590.
[12]Lorenz K,Lal R. Biogeochemical C and N cycles in urban soils[J]. Environment International,2009,35(1):1-8.
[13]Kaye J P,Mcculley R L,Burke I C. Carbon fluxes,nitrogen cycling,and soil microbial communities in adjacent urban,native and agricultural ecosystems[J]. Global Change Biology,2005,11(4):575-587.
[14]李房英,吴少华,雷波.福州市园林绿地植物应用调查研究[J]. 城市环境与城市生态,2002,15(1):50-52.
[15]Cramer M,Myers J. Growth and photosynthetic characteristics of euglena gracilis[J]. Archives of Microbiology,1952,17(1/2/3/4):384-402.
[16]Talling J F. Photosynthetic characteristics of some freshwater plankton diatoms in relation to underwater radiation[J]. New Phytologist,1957:29-50.
[17]Van T K,Haller W T,Bowes G. Comparison of the photosynthetic characteristics of three submersed aquatic plants[J]. Plant Physiology,1976,58(6):761-768.
[18]Koehl M R,Alberte R S. Flow,flapping,and photosynthesis of Nereocystis leutkeana:a functional comparison of undulate and flat blade morphologies[J]. Marine Biology,1988,99(3):435-444.
[19]Karlsson P S. Photosynthetic characteristics and leaf Carbon economy of a deciduous and an evergreen dwarf shrub:Vaccinium uliginosum L.and V.vitisidaea L.[J]. Ecography,1985,8(1):9-17.
[20]张璐,张纪林,教忠意,等. 不同光照条件下3种冬青属植物的光合特征日变化研究[J]. 西北植物学报,2006,26(3):490-495.
[21]郑淑霞,上官周平. 不同功能型植物光合特性及其与叶氮含量、比叶重的关系[J]. 生态学报,2007,27(1):171-181.
[22]龚吉蕊,赵爱,苏培玺,等. 黑河流域几个主要植物种光合特征的比较研究[J]. 中国沙漠,2005,25(4):587-592.
[23]常宗强,冯起,苏永红,等. 额济纳绿洲胡杨的光合特征及其对光强和CO2浓度的响应[J]. 干旱区地理,2006,29(4):496-502.
[24]刘绍辉,方精云. 土壤呼吸的影响因素及全球尺度下温度的影响[J]. 生态学报,1997,17(5):19-26.
[25]李玉宁,王关玉,李伟. 土壤呼吸作用和全球碳循环[J]. 地学前缘,2002,9(2):351-357.
[26]王淼,姬兰柱,李秋荣,等. 土壤温度和水分对长白山不同森林类型土壤呼吸的影响[J]. 应用生态学报,2003,14(8):1234-1238.
郁珊珊 王浩 王亚军


摘要:为了探寻福州市不同绿地生态系统(草地、林地、耕地和园地)光合特征和土壤呼吸变化规律及其影响因素,利用指数模型和相关系数分析土壤呼吸与土壤温度、湿度和光合特征的关系。结果表明:(1)4种绿地生态系统植物LSP、LCP、AQY、CE、Ls和WUE基本表现出一致的变化规律,综合表现为林地和耕地显著高于草地和园地;(2)1 d中各时期净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为林地和耕地显著高于草地和园地,4种绿地生态系统Pn和EVAP日变化均呈双峰曲线,气孔限制导致了光合“午休”现象,Ci和Gs日变化随时间呈“W”字形变化规律,通过SPSS最佳曲线拟合4种绿地生态系统植物的Pn与PAR、CO2浓度和温度变化呈一致的二次曲线关系;(3)在日的时间尺度上,4种绿地生态系统土壤呼吸均表现为单峰型,且峰值出现的时间基本一致,约在14:00达到最大,最低值出现在06:00,相同时间基本表现为林地和耕地显著高于草地和园地;(4)4种绿地生态系统土壤呼吸与土壤温度和湿度之间关系以指数方程拟合最好,通过偏相关分析可知,草地、林地、耕地和园地土壤呼吸均与Pn、EVAP和WUE呈显著或者极显著的相关关系,而与其他光合生理指标基本没有显著相关关系(P>005),由此表明土壤温度、土壤湿度、Pn、EVAP和WUE是影响福州市4种绿地生态系统土壤呼吸的主要环境因子。
关键词:福州市;绿地生态系统;光和特性;土壤呼吸
中图分类号: S181.6文献标志码: A文章编号:1002-1302(2016)06-0483-05
收稿日期:2016-01-25
项目基金:国家自然科学基金(编号:2014BC210);福建省厦门市科技专项课题(编号:14020041)。
作者简介:郁珊珊(1981—),女,福建厦门人,博士,副教授,研究方向为风景园林规划与设计、城市绿地生态系统规划。E-mail:shanshan_yu81@163.com
通信作者:王浩,博士,教授,博士生导师,研究方向为风景园林规划与设计。城市绿地生态系统是由各类城市绿地(草地、林地、耕地、公园绿地等)相互联系组合的城市绿色环境体系[1-2]。具有自净能力、自动调节能力和生命力,在改善城市生态环境、美化市容、调节城市生态平衡中发挥着主导作用,已成为衡量城市生态文明和城市现代化的重要标志[3]。城市绿地可以通过植物的蒸腾、蒸散、吸收、吸附、反射等功能,降低温度,增加湿度,固碳释氧,抗污染(吸收粉尘、Cl2、SO2、CO等),降低噪音,保护生物多样性等[4]。随着生态城市概念的提出、建设和发展,人们日益注意到城市绿地的生态意义和环境价值[5],而快速工业化及城市化给城市绿地的生长环境造成了巨大的胁迫效应,加速了城市的“生态环境危机”[6-7]。
光照是城市绿地植物生命活动中起重大作用的生存因子,也是对城市绿地植物光合结构重要的环境因素,城市绿地植物需要在一定的光照条件下完成生长发育过程,但不同城市绿地植物对光照强度的适应范围亦有所差别[8-9]。土壤作为大气CO2重要源或汇,土壤CO2细微的改变都会显著改变大气中CO2的浓度和碳的累积速率[10]。在陆地生态系统中,土壤碳库是大气碳贮量的2~3倍,通过土壤呼吸作用向大气释放的CO2约占全球CO2交换量的25%[11]。同时,土壤呼吸受多种因素的共同交互影响,包括土壤温度、湿度、土壤养分、净生态系统生产力(NEP)和人类干扰等,通过测定城市绿地生态系统土壤呼吸速率特征并阐明影响土壤呼吸、植物光合相互作用关系及其环境因子,不仅有利于改变人们对城市绿化植被的认识,更有利于科学地选取和配置绿地绿化植物,使其最大化发挥其生态效益,从而促进城市绿地系统规划、优化植物群落结构、改善环境质量、促进城市生态建设[12-13]。因此,本研究采用美国Li-6400便携式气体分析系统和 Li-6400 土壤呼吸室测量福建省福州市绿地生态系统植物光合特性、土壤呼吸速率及影响因子,以期深入了解植物光合特性和土壤呼吸作用的变化过程及变化机理,从而为城市绿地系统建设中植物的选择和应用提供科学依据。
1材料与方法
1.1研究区概况
福州市位于福建省东部沿海闽江下游(25°15′~26°39′N,118°08′~120°31′E),气候资源丰富、温暖湿润、四季常青、雨量充沛、霜少无雪、多低山丘陵,农业活动对土壤的影响深刻,水文条件各不相同,地貌类型以丘陵为主,地形复杂多样,地势起伏不平,土壤类型较多,主要土类有黄棕壤、红壤、石灰岩土、紫色土、潮土和沼泽土,平均海拔9.2 m,无霜期达 326 d,年均日照数为1 700~1 980 h,年均降水量为900~2 100 mm,年均气温为16~20 ℃,年相对湿度约77%。福州市土地总面积为12 251 km2,其中林地面积7 032 km2,占土地总面积的57.4%,绿地面积81.2 km2,绿地率达36.9%,绿化覆盖面积88.7 km2,绿化覆盖率高达40.3%,人均公园绿地高达11.2 m2。曾荣获“全国文明城市”“全国绿化模范城市”“全国城乡绿化先进集体”“国家卫生城市”和“中国优秀旅游城市”等光荣称号[14]。
1.2测定方法
1.2.1土壤呼吸的测定选择保护区内落叶季雨林和常绿季雨林,每个林分布设2个监测点,相距80 m,在第1次测定土壤呼吸之前,提前1 d将测定基座(soil collar)嵌入土壤中,在每个基座内存留的地表植被自土壤表层彻底剪除,聚氯乙烯圆柱体经过24 h的平衡后,土壤呼吸速率会恢复到基座放置前的水平,从而避免了因安置气室对土壤扰动而造成的短期内呼吸速率波动。采用动态密闭气室红外CO2分析仪(IRGA)法,测定仪器型号为美国Li-6400便携式气体分析系统和Li-6 400土壤呼吸室,土壤呼吸日变化测定为白天每隔2 h测定1次,每个监测点1次测定2个重复,取平均值。季节变化测定为2015年7月中旬,在08:00—10:00之间测量,有报道指出该时间段测定值最为接近24 h平均值,具有较好的代表性。在观测土壤呼吸的同时,使用手持长杆电子温度探针(SK-250 WP,Sato Keiryoki,Kanda,Japan)测定0~10 cm深处的土壤温度,使用时域反射仪(Model TDR 300,Spectrum,Aurora,USA)测定每个监测点附近0~10 cm范围内的土壤湿度,每个样地温湿度安装数据采集器(HOBO ProRH/TEMP Data Logger),以获取距地面50 cm高度的大气温度和相对湿度数据,用于分析土壤呼吸速率和水热因子的关系。
1.2.2光合特性测定(1)美国LI-COR公司生产的LI-6400系列便携式光合作用测量系统进行气体交换光合生理特征测定,2015年6月下旬07:00—19:00(晴天无风),每隔 2 h 测定1次,连续重复测定3 d,每个处理选择3株健康油茶重复测定,每株选取当年生的成熟叶片(从植株顶部数第3~6片完全展开的成熟叶),轮流测定以消除测定时间上的误差。红蓝光源光照设定为1 000 μmol/(m2·s),温度30 ℃,CO2流量为400 μmol/s,不同测定日期温度变化区间为22.1~35.4 ℃,辐射强度(PAR)为112~1 371 μmol/(m2·s),空气相对湿度在24.6%~45.7%之间,选取充分受光、叶位一致的叶片(上、中、下当年生成熟叶)进行测定,轮流测定以消除测定时间上的误差。仪器记录净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)、胞间CO2浓度(Ci)、环境CO2浓度(Ca)和光照强度(PAR),其中PAR和CO2浓度以外界条件为准。
(2)光合-光强响应特性的测定,测定时CO2摩尔分数控制为350 μmol/mol,温度为25 ℃,湿度约为大气湿度的99%。设定光照度梯度为1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、0 μmol/(m2·s),每个光照度下适应 5 min 后测定Pn值。根据Pn-PAR回归方程求出各品种的光补偿点(LCP)、光饱合点(LSP);表观量子效率(AQY)根据低于200 μmol/(m2·s)光照度下测定的叶片PFD-Pn曲线计算求得;CO2羧化效率(CE)根据CO2 250 μmol/(m2·s)下测定的叶片CO2-Pn曲线计算求得[15]。
(3)气孔限制值(Ls)和叶片瞬时水分利用效率(WUE)由公式计算,即Ls=1-Ci/Ca,WUE=Pn/EVAP[16]。
1.3数据处理与分析
采用Excel 2007和SPSS 18.0进行数据分析,单因素方差分析(One-way ANOVA)和最小显著法(LSD)检验其差异显著性,Pearson相关性系数检验各指标的相关性,采用指数方程拟合土壤呼吸与土壤温度和湿度的响应关系,由原始数据拟合得到的多元回归关系经统计学检验得到拟合度参数R2和Radj2,并在P<0.01水平检验相关系数的显著性。
2结果与分析
2.1绿地生态系统植物光合特性
2.1.1绿地生态系统植物叶片生理特性由图1可知,福州市绿地生态系统植物光饱和点(LSP)变化范围为658.9~1 235.8 μmol/(m2·s),依次表现为林地>耕地>草地>园地,4种绿地LSP差异均显著(P<0.05);光补偿点(LCP)变化范围为74.3~125.3 μmol/(m2·s),依次表现为林地>耕地>草地>园地,其中草地和园地差异不显著(P>0.05),二者显著低于林地和耕地(P<0.05);表观量子效率(AQY)变化范围为18.9×10-3~33.5×10-3,依次表现为林地>耕地>园地>草地,4种绿地AQY差异均不显著(P>0.05);羧化效率(CE)变化范围为32.7×10-3~69.7×10-3,依次表现为耕地>林地>草地>园地,其中园地显著低于其他绿地
(P<0.05);气孔限制值(Ls)变化范围为0.51~0.93 μmol/(m2·s),依次表现为耕地>林地>草地>园地,其中耕地与林地之间差异并不显著(P>0.05),二者显著高于草地和园地(P<0.05);水分利用效率(WUE)变化范围为2.59~6.58 μmol/(m2·s),依次表现为林地>耕地>草地>园地,4种绿地WUE差异均显著(P<0.05)。
2.1.2绿地生态系统植物叶片光合特性福州市绿地生态系统植物叶片光合特性见图2,4种绿地生态系统植物叶片光合指标在1 d之中的变化趋势基本保持一致,1 d中各时期净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为林地和耕地>草地和园地,局部有所波动;绿地生态系统植物叶片光合速率(Pn)日变化均表现为典型的双峰型曲线,第1峰出现在约10:00,12:00—13:00光合速率出现峰谷,第二高峰出现在约16:00,但明显小于第1峰值,16:00以后光合速率明显下降,表现出明显的光合“午休”现象;胞间CO2浓度(Ci)日变化随时间呈“W”字形变化规律,在约10:00出现第1个波谷,12:00达到最大(峰值),12:00 以后急剧下降,约16:00出现第2峰谷,第2波谷值要高于第1波谷值,而后随时间的变化出现缓慢上升趋势;气孔导度(Gs)日变化随时间呈“W”字形变化规律,其第1波峰出现在08:00,在10:00出现第1个波谷,第2波峰出现在13:00,但第2波峰明显低于第1波峰,16:00出现第1次峰谷,而后随时间的变化缓慢上升;蒸腾速率(EVAP)日变化规律与光合速率(Pn)保持一致,均呈双峰曲线,局部峰值推迟或者提前。
2.1.3绿地生态系统植物叶片光合特性与生理响应由表1可知,通过SPSS最佳曲线拟合(分别拟合光合速率Pn与PAR、CO2浓度和温度之间的曲线关系),筛选出R2最大(最佳拟合度及校准以后的最大R2)和P值(显著性)最小的曲线组合,并在此基础上检验其F值和差异显著性(P<0001)。福州市绿地生态系统植物的Pn-PAR呈一致的二次曲线关系,曲线拟合方程见表3。
2.2绿地生态系统土壤呼吸特性
2.2.1绿地生态系统土壤呼吸日变化由图3可知,4种绿地生态系统土壤呼吸存在明显的日变化规律, 均表现为单峰型,且峰值出现的时间基本一致,土壤呼吸速率都在约14:00达到最大,最低值出现在06:00,在06:00—10:00,4种绿地生态系统土壤呼吸升高缓慢,12:00以后急剧上升,达到最大值以后急剧降低。草地土壤呼吸日变化范围为0.73~1.85 μmol/(m2·s),林地土壤呼吸日变化范围为0.84~2.06 μmol/(m2·s),耕地土壤呼吸日变化范围为0.52~217 μmol/(m2·s),园地土壤呼吸日变化范围为0.46~162 μmol/(m2·s)。相同时间土壤呼吸速率基本表现为耕地>林地>草地>园地。
2.2.2土壤温度和湿度对土壤呼吸的影响为进一步探讨绿地生态系统土壤呼吸与其显著影响因子土壤温度和湿度的关系,国内外学者一般采用线性模型、二次方程、指数模型等多种方法进行拟合,本试验结果中将土壤呼吸强度与土壤 10 cm 温度进行曲线拟合,筛选出最佳拟合度的方程(R2最大,P值最小),得出的不同拟合方程;由表4可知,土壤呼吸与土壤温度之间关系以指数方程拟合最好。4种绿地生态系统土壤呼吸与土壤温度的指数关系达到了极显著水平(P<0.01),且指数模型的决定系数最大,故指数模型的拟合效果最好。4种绿地生态系统土壤呼吸与土壤湿度之间关系也以指数方程拟合最好。
2.3土壤呼吸与光合的相关关系
由绿地生态系统土壤呼吸与植物叶片光合特征参数标准化经SPSS双变量Pearson相关系数法分析的结果(表5)可知,土壤呼吸主要是植物的根以及土壤微生物的呼吸,植物的生长、微生物的氧化分解等活动都要受周围环境如土壤的温度、土壤的水分、气温、空气湿度等的影响,因此,这些环境因子也是影响土壤呼吸强度的重要因素。由表5可知,4种绿地生态系统土壤呼吸强度与各环境因子的Pearson相关关系均显著,但有时由于某一共同变量的干扰会导致2个变量间的假相关,为排除其他因素的干扰,对各因子分别作偏相关分析,控制其他几个变量以分析土壤呼吸与另一个变量的净相关关系。结果表明,草地、林地、耕地和园地土壤呼吸均与Pn、EVAP和WUE呈显著(P<0.05)或极显著(P<0.01)的相关关系,而与其他光合生理指标基本没有显著相关关系(P>0.05)。
3讨论
3.1绿地生态系统光合特性特征分析
光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)、羧化效率(CE)、气孔限制值(Ls)和水分利用效率(WUE)均能衡量植物对强光或弱光的利用能力。本研究中4种绿地生态系统植物LSP、LCP、AQY、CE、Ls和WUE基本表现出一致的变化规律,综合比较可知林地和耕地显著高于草地和园地,主要是受植被类型及植物本身特性的影响,林地和耕地植物叶片对低CO2浓度和低光强的利用率最强,具有较强的再生速率。此外,本研究中光合生理指标属于瞬间测定的结果,受环境影响较大,所测定的结果有一定的偏差,后续还需要进行反复大量的野外监测来验证本试验的研究结果。
植物的光合特性反映了对外界环境的响应和适应,受到众多环境因素的影响,其中影响光合速率的因素主要是气孔限制和非气孔限制因素,即气孔的开张和叶肉细胞的光合活性[17]。4种绿地生态系统Pn日变化表现为双峰曲线,有明显的“午休”现象,气孔导度(Gs)和胞间CO2浓度(Ci)变化方向与净光合速率(Pn)变化方向相反,1 d之中相同时间段净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和空气中CO2浓度基本表现为林地和耕地显著高于草地和园地,主要是由于林地和耕地(灌木乔木为主)比草地和园地(草本为主)能够利用更多的时间合成有机物而增产,中午持续的强光使气温、蒸腾速率急剧增加,叶片水分代谢失调,叶面温度过高而抑制了参与光合过程酶的活性,气孔导度(Gs)、胞间CO2浓度(Ci)和空气中CO2浓度导致Pn下降,从而造成了光合“午休”现象[18-19]。气孔是水分与叶片进行气体交换的通道,也是CO2进入细胞的门户,控制着叶片和大气之间的CO2和水分交换,气孔的闭合程度对光合与蒸腾作用会产生直接的影响,本研究中4种绿地生态系统气孔导度(Gs)日变化与蒸腾速率(EVAP)曲线相似,说明气孔的开闭程度直接影响蒸腾作用,主要是由水蒸气浓度差及水蒸气扩散阻力所决定[20];EVAP与Gs变化趋势一致,主要是由于叶内外水蒸气压差的增大和水蒸气浓度减少,加剧了植物体蒸腾扩散阻力,从生理生态的角度看,光合和蒸腾分别是CO2和H2O通过叶片气孔的内外物质交换过程,气孔行为对光合与蒸腾将产生直接影响,叶内外水蒸气压差的增大和水蒸气浓度减少,加剧了植物体蒸腾扩散阻力[21-22];本研究中在绝大多数时段Ci和Gs变化方向与其Pn变化方向相反,气孔主导其光合速率的变化,光合速率绝大多数取决于气孔限制因素,即叶肉细胞光合能力的变化主导其光合速率的变化。同时,绿地生态系统植物也能采取不同的适应策略应对环境的变化,叶片水分饱和亏缺逐渐增加的同时,净光合速率和蒸腾速率逐渐减少,光合限制因素由气孔限制为主逐渐转变为非气孔限制为主[23]。此外,水分状况也是影响小麦光合作用最重要的因子之一,4种绿地生态系统具有较高的EVAP,说明其需水量较多,因此要加强对水分的管理。
通过SPSS最佳曲线拟合(分别拟合光合速率Pn与PAR、CO2浓度和温度之间的曲线关系),福州市4种绿地生态系统植物的Pn与PAR、CO2浓度和温度变化呈一致的二次曲线关系,由曲线计算可知绿地生态系统植物光合作用最适温度为23~25 ℃,这与前人的研究结论[20-21]一致。研究还发现,虽然绿地生态系统植物光合适温相差不大,但在最适温度时的光合速率却相差较大,说明温度的变化对4种绿地生态系统植物光合作用有较大的影响。不同植物的光合作用对温度的响应不同,可能与物种的温度适应性(原产地的气候、环境)有关。
3.2绿地生态系统土壤呼吸特征分析
福州市4种绿地生态系统大气温度存在显著的日变化,气温的日较差在早、晚可达10 ℃以上,白天较高的温度有利于植物进行光合作用,下午较低的温度下植物呼吸作用渐弱,有利于植物进行有机物质的积累,大气温度变化幅度大,14:00 温度达到最高,土壤呼吸速率对地表温度响应较快,因此土壤呼吸速率在此时段达到峰值。从4种绿地生态系统土壤呼吸的日变化分布特征可知,在白天林冠的调节作用趋向于在昼间热量充足时从外界获取能量,夜间则趋向于保持环境稳定,将林内外气温保持在一个可控的范围内。大量研究结果显示,森林、草原以及湿地等的土壤呼吸速率呈现单峰型特点,本研究中绿地生态系统土壤呼吸日和季节变化同样呈现明显的单峰曲线,日变化峰值出现在14:00,最小值约出现在06:00,此时段地表温度较低、湿度较大,此外,根系活动和呼吸作用较弱,因此土壤呼吸速率低,这与前人的研究结果[24-25]相一致。
土壤呼吸速率主要由温度、湿度、环境因子等共同作用所驱动,控制因子随生态系统类型和气候类型变化而变化。对于城市绿地生态系统,水热环境因子是影响其土壤呼吸最为主要的两大因素,土壤湿度在一定程度上降低土壤呼吸速率对土壤温度的响应,土壤湿度的限制作用可能是导致土壤呼吸速率的敏感性降低[24,26]。因此,土壤温度、湿度和养分含量相互作用影响着绿地生态系统土壤呼吸,同时,在本研究中坡度和坡向的变化也影响着绿地生态系统土壤呼吸。本研究中4种绿地生态系统土壤呼吸作用受土壤湿度和温度的影响较为明显,土壤呼吸速率与土壤(0~10 cm)湿度呈负指数相关关系,与土壤(0~10 cm)温度呈正指数相关关系,土壤温度和土壤湿度是影响福州市4种绿地生态系统土壤呼吸的重要环境因子。相关性分析可知,草地、林地、耕地和园地土壤呼吸均与Pn、EVAP和WUE呈显著(P<0.05)或者极显著(P<0.01)的相关关系,而与其他光合生理指标基本没有显著相关关系(P>0.05),由此表明土壤温度、土壤湿度、Pn、EVAP和WUE是影响福州市4种绿地生态系统土壤呼吸的主要环境因子。
参考文献:
[1]李锋,王如松. 城市绿地系统的生态服务功能评价、规划与预测研究——以扬州市为例[J]. 生态学报,2003,23(9):1929-1936.
[2]张浩,王祥荣. 城市绿地的三维生态特征及其生态功能[J]. 中国环境科学,2001,21(2):101-104.
[3]周廷刚,郭达志. 基于GIS的城市绿地景观空间结构研究——以宁波市为例[J]. 生态学报,2003,23(5):901-907.
[4]Harawa N T,Greenland S,Bingham T A,et al. Associations of race/ethnicity with HIV prevalence and HIV-related behaviors among young men who have sex with men in 7 urban centers in the United States[J]. Journal of Acquired Immune Deficiency Syndromes,2004,35(5):526-536.
[5]Tajima K. New estimates of the demand for urban green space:implications for valuing the environmental benefits of Bostons big dig project[J]. Journal of Urban Affairs,2003,25(5):641-655.
[6]何兴元,陈玮,徐文铎,等. 沈阳城区绿地生态系统景观结构与异质性分析[J]. 应用生态学报,2003,14(12):2085-2089.
[7]蔺银鼎,韩学孟,武小刚,等. 城市绿地空间结构对绿地生态场的影响[J]. 生态学报,2006,26(10):3339-3346.
[8]Erickson R A,Jimenez R. Microfluidic cytometer for high-throughput measurement of photosynthetic characteristics and lipid accumulation in individual algal cells[J]. Lab on a Chip,2013,13(15):2893-2901.
[9]Yuan L,Shu S,Sun J,et al. Effects of 24-epibrassinolide on the photosynthetic characteristics,antioxidant system,and chloroplast ultrastructure in Cucumis sativus L. under Ca(NO3)2 stress[J]. Photosynthesis Research,2012,112(3):205-214.
[10]Gomez-Casanovas N,Matamala R,Cook D R. Net ecosystem exchange modifies the relationship between the autotrophic and heterotrophic components of soil respiration with abiotic factors in prairie grasslands[J]. Global Change Biology,2012,18(8):2532-2545.
[11]Reid J P,Adair E C,Hobbie S E,et al. Biodiversity,nitrogen deposition,and CO2 affect grassland soil carbon cycling but not storage[J]. Ecosystems,2012,15(4):580-590.
[12]Lorenz K,Lal R. Biogeochemical C and N cycles in urban soils[J]. Environment International,2009,35(1):1-8.
[13]Kaye J P,Mcculley R L,Burke I C. Carbon fluxes,nitrogen cycling,and soil microbial communities in adjacent urban,native and agricultural ecosystems[J]. Global Change Biology,2005,11(4):575-587.
[14]李房英,吴少华,雷波.福州市园林绿地植物应用调查研究[J]. 城市环境与城市生态,2002,15(1):50-52.
[15]Cramer M,Myers J. Growth and photosynthetic characteristics of euglena gracilis[J]. Archives of Microbiology,1952,17(1/2/3/4):384-402.
[16]Talling J F. Photosynthetic characteristics of some freshwater plankton diatoms in relation to underwater radiation[J]. New Phytologist,1957:29-50.
[17]Van T K,Haller W T,Bowes G. Comparison of the photosynthetic characteristics of three submersed aquatic plants[J]. Plant Physiology,1976,58(6):761-768.
[18]Koehl M R,Alberte R S. Flow,flapping,and photosynthesis of Nereocystis leutkeana:a functional comparison of undulate and flat blade morphologies[J]. Marine Biology,1988,99(3):435-444.
[19]Karlsson P S. Photosynthetic characteristics and leaf Carbon economy of a deciduous and an evergreen dwarf shrub:Vaccinium uliginosum L.and V.vitisidaea L.[J]. Ecography,1985,8(1):9-17.
[20]张璐,张纪林,教忠意,等. 不同光照条件下3种冬青属植物的光合特征日变化研究[J]. 西北植物学报,2006,26(3):490-495.
[21]郑淑霞,上官周平. 不同功能型植物光合特性及其与叶氮含量、比叶重的关系[J]. 生态学报,2007,27(1):171-181.
[22]龚吉蕊,赵爱,苏培玺,等. 黑河流域几个主要植物种光合特征的比较研究[J]. 中国沙漠,2005,25(4):587-592.
[23]常宗强,冯起,苏永红,等. 额济纳绿洲胡杨的光合特征及其对光强和CO2浓度的响应[J]. 干旱区地理,2006,29(4):496-502.
[24]刘绍辉,方精云. 土壤呼吸的影响因素及全球尺度下温度的影响[J]. 生态学报,1997,17(5):19-26.
[25]李玉宁,王关玉,李伟. 土壤呼吸作用和全球碳循环[J]. 地学前缘,2002,9(2):351-357.
[26]王淼,姬兰柱,李秋荣,等. 土壤温度和水分对长白山不同森林类型土壤呼吸的影响[J]. 应用生态学报,2003,14(8):1234-1238.
猜你喜欢
——福州市冯宅中心小学简介(二)
福建基础教育研究(2020年9期)2020-10-21
——福州市冯宅中心小学简介(一)
福建基础教育研究(2020年9期)2020-10-21
红土地(2019年10期)2019-10-30
福建基础教育研究(2019年12期)2019-05-28
福建基础教育研究(2019年5期)2019-05-28
福州党校学报(2018年4期)2018-12-29
防护林科技(2017年1期)2017-02-13
——为悼念福州市政协原副主席、福州市老促会原副会长高伯英同志而作
红土地(2017年4期)2017-01-25
亚热带资源与环境学报(2015年4期)2015-01-22
