
2014年冬季浙江中部海域网采浮游植物群集
2016-07-25 09:52刘亚林黄秀清蒋晓山王航俊齐安翔
海洋科学 2016年4期
关键词:冬季
刘亚林,黄秀清,蒋晓山,王航俊,邹 清,齐安翔
(1. 国家海洋局 温州海洋环境监测中心站,浙江 温州 325000; 2. 上海东海海洋工程勘察设计研究院,上海200137; 3. 国家海洋局 东海预报中心,上海 200137)
2014年冬季浙江中部海域网采浮游植物群集
刘亚林1,黄秀清2,蒋晓山1,王航俊1,邹 清1,齐安翔3
(1. 国家海洋局 温州海洋环境监测中心站,浙江 温州 325000; 2. 上海东海海洋工程勘察设计研究院,上海200137; 3. 国家海洋局 东海预报中心,上海 200137)
摘要:于2014年12月至2015年1月,在121°42′E~126°18′E,28°43′N~30°02′N的浙江中部海域,对97个站位采集的浮游植物的种类组成、群落结构、细胞丰度进行了研究。共鉴定浮游植物6门53属163种(含19未定种),其中以硅藻为主,甲藻其次; 浙江中部海域冬季浮游植物群落可分为4种生态类型: 半咸水类群、沿岸广温型、近海广温广盐型、外海高温高盐型,其中最主要生态类型为近海广温型; 浮游植物优势种为中华齿状藻(Odentella sinensis)、琼氏圆筛藻(Coscinodiscus jonesianus) 、伏氏海线藻(Thalassionema frauenfeldii)、虹彩圆筛藻(Coscinodiscus oculus-iridis)、太阳双尾藻(Ditylum sol)、洛氏角毛藻(Chaetoceros lorenzianus)。浮游植物的细胞丰度范围为0.3×104个/m3~67.1×104个/m3,平均值为 8.8×104个/m3,呈现近岸>近海>外海的分布特点,高值区位于三门湾至渔山列岛之间的海域; 生物多样性指数平均值为3.36; Pearson相关性分析得出: 调查海域浮游植物的细胞丰度与盐度呈负相关,与磷酸盐、硝酸盐成正相关。
关键词:浙江中部海域; 冬季; 浮游植物; 群集结构
浮游植物是海洋的主要的初级生产者,是海洋食物网的基础环节,在海洋生态系统的物质循环和能量转换的过程中起着重要作用[1]。浮游植物种群变动和群落结构直接影响海洋生态系统的结构和功能。浮游植物的时空变化特征与环境因子关系密切,生态系统中环境因子的改变直接作用于浮游植物群落结构。研究浮游植物群落结构是研究海洋生态环境问题的基础[2]。
浙江中部海域位于东海,属于亚热带和温带气候,利于浮游生物的繁殖和生长,是各种鱼虾繁殖和栖息的良好场所。对东海浮游植物的研究众多,成果显著,郭玉洁[4]、吴玉霖[10]、何青[11]在长江口海域,钟惠英[9]、朱根海[25]在三门湾,陆斗定[12]等在舟山群岛,杨清良[22]、孙军[24]、陆斗定[13-15,18]在黑潮区这些东海典型海域进行了浮游植物群落结构或特定种类的研究,郭玉洁[3]、罗民波[6]、郭术津[7]、秦玉涛[8]、王丹[27]在东海海域进行了不同季节的大尺度浮游植物群落结构研究,但从 2010年以来,对东海特别是浙江海域冬季大尺度的浮游植物研究较少,特别是从近岸到近海,从近海到外海连续性的浮游植物群落结构的研究较少,本次调查于2014年12月至2015 年1月,在121°42′~126°18′E,28°43′~30°02′N的浙江中部海域研究浮游植物的群落结构特征、生态类型和其主要优势种,对比研究了浙江中部从近岸到外海不同海域的浮游植物细胞丰度和生物多样性的分布特征,并对调查区浮游植物与环境因子进行了Pearson 相关性分析。
1 材料和方法
1.1 调查海域和采样方法
调查时间为2014年12月至2015年1月,调查海域为 121°42′~126°18′E,28°43′~30°02′N,共设 97个站位,从三门湾外湾经渔山列岛至东海外海,调查站位见图1。
共采集浮游植物样品97个,样品采集和鉴定均按照《海洋调查规范》[5]进行,采样网具采用小型浮游生物网(水深大于30 m使用,网具规格为: 网口内径37 cm,网长280 cm,筛绢孔径0.077 mm)和浅水
[Foundation: Marine environmental protection and ecological project in Zhejiang Province(ZJMFP[2012]100)]
温度、盐度、营养盐(NO3-N、NO2-N、NH4-N、PO4-P)样品的取样和分析参考海洋调查规范[5]进行测定。

图1 2014年冬季研究海域和调查站位图Fig. 1 Study area and sampling stations in the winter of 2014
1.2 数据处理
1.2.1 优势度
对群落的分析过程中,必须确定优势种的成分,因为优势种是决定群落性质的主要因素。本文各物种的优势度值则根据浮游植物的出现频率及细胞丰度来计算,计算公式为:
Yi= (ni/ N) × fi
式中,Yi为优势度; N为该海区出现的所有种类的总个体数; n为第i种的个体数; fi为该种在各站位出现的频率。
1.2.2 多样性指数
反映群落结构种类多样性的数学模式有许多,本文采用种类和数量信息函数表示的香农-韦弗(Shannon-Weaver,1963)多样性指数。
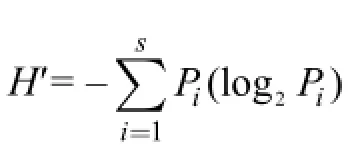
式中,H'为物种的多样性指数; s为样品中的种类总数; Pi为第i种在全部采样中的比例。
2 结果与讨论
2.1 种类组成和优势种
共鉴定浮游植物6门53属163种(含19未定种),其中硅藻门36属121种,甲藻门11属34种,绿藻门3种,蓝藻门3种、金藻门、裸藻门各1种。其中硅藻和甲藻占主要地位,绿藻、金藻、裸藻均有发现,但数量和种类很少。
冬季整个调查海域主要优势种(表1)有中华齿状藻(Odentella sinensis)(Y=0.083)、琼氏圆筛藻(Coscinodiscus jonesianus)(Y=0.053)、伏氏海线藻(Thalassionema frauenfeldii)(Y=0.040)、虹彩圆筛藻(Coscinodiscus oculus-iridis)(Y=0.031)、太阳双尾藻(Ditylum sol)(Y=0.023)、洛氏角毛藻(Chaetoceros lorenzianus)(Y=0.021)。其中罗民波在2000年1月份的东海浮游植物调查研究中[6]报道,东海冬季的主要的浮游植物优势种为中华齿状藻和洛氏角毛藻,而且研究了中华齿状藻和洛氏角毛藻对温度的适应性,发现中华齿状藻和洛氏角毛藻能够在水温较低的冬春季出现,说明其在低温幅范围有较强的适应能力。郭术津、孙军[7]在2009年冬季的东海浮游植物调查研究中报道东海的主要优势物种为具槽帕拉藻(Paralia sulcata)、菱形海线藻(Thalassionema nitzschioides)、柔弱伪菱形藻(Pseudo-nitzschia delicatissima)、伏氏海线藻、虹彩圆筛藻、细弱海链藻(Thalassiosira subtilis)等。秦玉涛[8]在2006年12月浙江及福建北部海域浮游植物调查研究中,发现主要优势种为菱形海线藻、铁氏束毛藻(Trichodesmium thiebautii)、细弱海链藻。
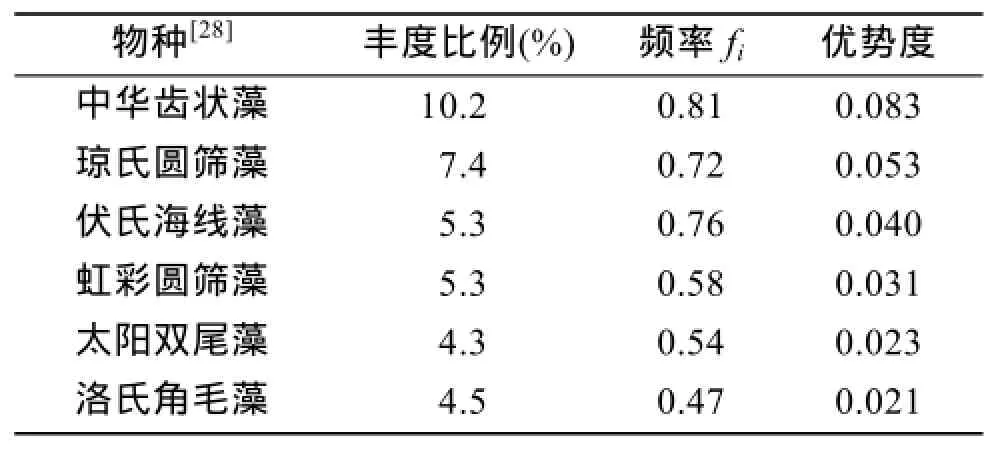
物种[28] 丰度比例(%) 频率fi 优势度中华齿状藻 10.2 0.81 0.083琼氏圆筛藻 7.4 0.72 0.053伏氏海线藻 5.3 0.76 0.040虹彩圆筛藻 5.3 0.58 0.031太阳双尾藻 4.3 0.54 0.023洛氏角毛藻 4.5 0.47 0.021
调查海域从近岸到外海,水系复杂,受到黑潮、台湾暖流、长江冲淡水及钱塘江、甬江、椒江等多重水团影响,浮游植物种类丰富,生态类型多样,按照种类的生境、生态特征,可大体分成如下4个生态类型:
(1) 半咸水类型: 该类型种类和数量都很少,半咸水种主要分布在台州三门湾到宁波象山等海湾河口区域,该区域受陆地冲淡水影响较大。本次调查的代表种为波罗的海布纹藻(Gyrosigma balticum)和蛇目圆筛藻(Coscinodiscus argus)。钟慧英[9]在1986年2月在三门湾健跳港浮游植物调查中曾发现淡水藻种盘星藻(Pediastrum sp.)。本次调查在近岸站位中也发现了该藻种。
(2) 沿岸广温型: 该类型种类数量不多,但常在沿岸海域形成优势种,该类群主要出现在象山石浦到渔山列岛附近海域,本次调查发现的此种类型的代表种有中肋骨条藻(Skeletonema costatum)、具槽帕拉藻、派格棍形藻(Bacillaria paxillifera)等。吴玉霖[10]在2002年1月,何青[11]在2006年2月长江口及其邻近海域进行的浮游植物调查中,均发现具槽帕拉藻、中肋骨条藻在长江口海域是主要的优势种。
(3) 近海广温广盐型: 该类型种类数最多,数量较大,在各调查站位中出现的频率较高,广泛的分布在从近岸到远海的大部分海域,大多数优势种是该类型的,该类型的代表种为中华齿状藻、琼氏圆筛藻、虹彩圆筛藻、旋链角毛藻(Chaetoceros curvisetus)、伏氏海线藻、菱形海线藻、三角角藻(Ceratium tripos)和梭角藻(Ceratium fusus)。陆斗定[12]在马鞍列岛的秋冬季浮游植物调查中发现主要优势种有中华齿状藻、菱形海线藻、梭角藻。秦玉涛[8]在2006冬季浙江及其福建北部海域优势种中有菱形海线藻。
(4) 高温高盐型类型:该类群的种类和数量均不多,代表种为中华齿状藻热带型、大鸟尾藻(Ornithocercus magnificus)、和舒氏方形鸟尾藻(Ornithocercus quadratus v.quadratus.f.)、束毛藻、马西里亚角藻(Ceratium massilense)、波状角藻(Ceratium trichoceros)、科氏角藻(Ceratium kofoidii)等,该类型的在外海即受台湾暖流和黑潮影响的海域常见,尤其是上述藻类中的甲藻可作为黑潮指示种[13-15]。中华齿状藻热带型在6~7月出现于29°N以南和东南的黑潮暖流区,8~9月中国台湾暖流北上,可达 30°N(舟山外海)是良好的暖水指示种[16]; 大鸟尾藻和舒氏方形鸟尾藻仅在外海站位出现,是高温高盐的代表种,刘东艳等[17]在琉球群岛及其邻近海域甲藻的分布特征的研究中指出: 鸟尾藻可作为黑潮的指示种; 陆斗定[18]在研究东海黑潮区鸟尾藻的分布上报道,春夏两季,黑潮主轴一般在200 m等深线附近,秋冬两季,黑潮大规模入侵陆架区,尤其冬季更加明显,黑潮暖水可达 100 m 等深线附近[19],而本次冬季调查所发现鸟尾藻的站位水深范围在60~80 m,鸟尾藻的分布清晰地反映暖水边界位置的推移变化动态,根据此次调查鸟尾藻分布区域来推断,2014年冬季黑潮入侵陆架区的现象依然存在且有所加剧。束毛藻一直是东海常见种,且一直作为黑潮和台湾暖流的指示种[20-21],杨清良[22]统计南黄海和东海陆架区束毛藻的密度为1977年冬季(12月至1月)2 600条/m3,本次调查发现束毛藻的密度为3 200条/m3。《浙江及福建北部海域环境调查与研究》[23]一书中提到 2008年冬季的浮游植物调查中,束毛藻为510条/m3。
关于东海浮游植物的生态类型划分,郭术津、孙军[7]将东海浮游植物分成三部分: 位于长江口近岸和闽浙沿岸海域的近岸群集、位于东海外侧的东海-黑潮群集及占据剩余大部分东海海域的近海群集。近岸群集以具槽帕拉藻为主要优势种,东海-黑潮群集以铁氏束毛藻、科氏角藻和裸甲藻等为优势种,而近海群集则以菱形海线藻和伪菱形藻为优势种。比较本文划分的4种生态类型和郭术津、孙军划分的3个群集,半咸水类型和近岸广温型的集合和近岸群集高度相似,高温高盐性类型和东海-黑潮群集高度相似,近海广温型和近海群集高度相似。生态类型或群集的名称不同,但划分的核心依据是以浮游植物对温度和盐度的适应性以及所分布的地理特征来有所区别。
2.2 细胞丰度
因调查站位呈带状从近岸到外海布设,即从三门湾外湾经渔山列岛至外海,为科学客观的了解调查海域浮游植物的分布趋势,以经度为界,把调查海域划分为122°E以西(含9个站位)、122°~123°E(含23个站位)、123°~124°E(含16个站位)、124°~125°E(含16个站位)、125°E以东(含33个站位)五个区块,每个区块所含站位的浮游植物细胞丰度分布如图2所示。
冬季浮游植物细胞丰度平均值为 8.8×104个/m3,从图 2可以看出,浮游植物细胞丰度的分布趋势呈现近岸>近海>外海的规律。浮游植物分布的高值区在三门湾外湾至渔山列岛之间的海域。此海域岛屿众多,渔山列岛、大陈岛、头门岛分布其中,也是东海传统的渔场大陈渔场所在地。外海浮游植物细胞丰度较低,孙军[24]在研究琉球群岛及其邻近海域浮游硅藻分布的研究中报道: 琉球群岛岛链西侧浮游硅藻细胞丰度一般大于 5.0×104个/m3,岛链东侧浮游硅藻细胞丰度一般小于 5.0×104个/m3,在大洋中浮游硅藻的细胞丰度偏低。
比较历年东海区冬季浮游植物调查资料,见表 2,浮游植物细胞丰度分布基本呈现河口海湾等近岸近海海域大于外海的特点。
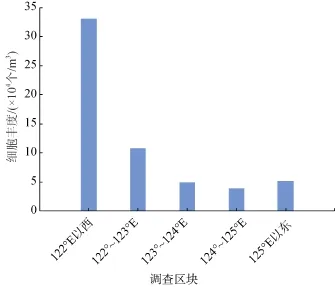
图2 调查海域浮游植物细胞丰度分布图Fig. 2 Phytoplankton cell abundance in the survey area
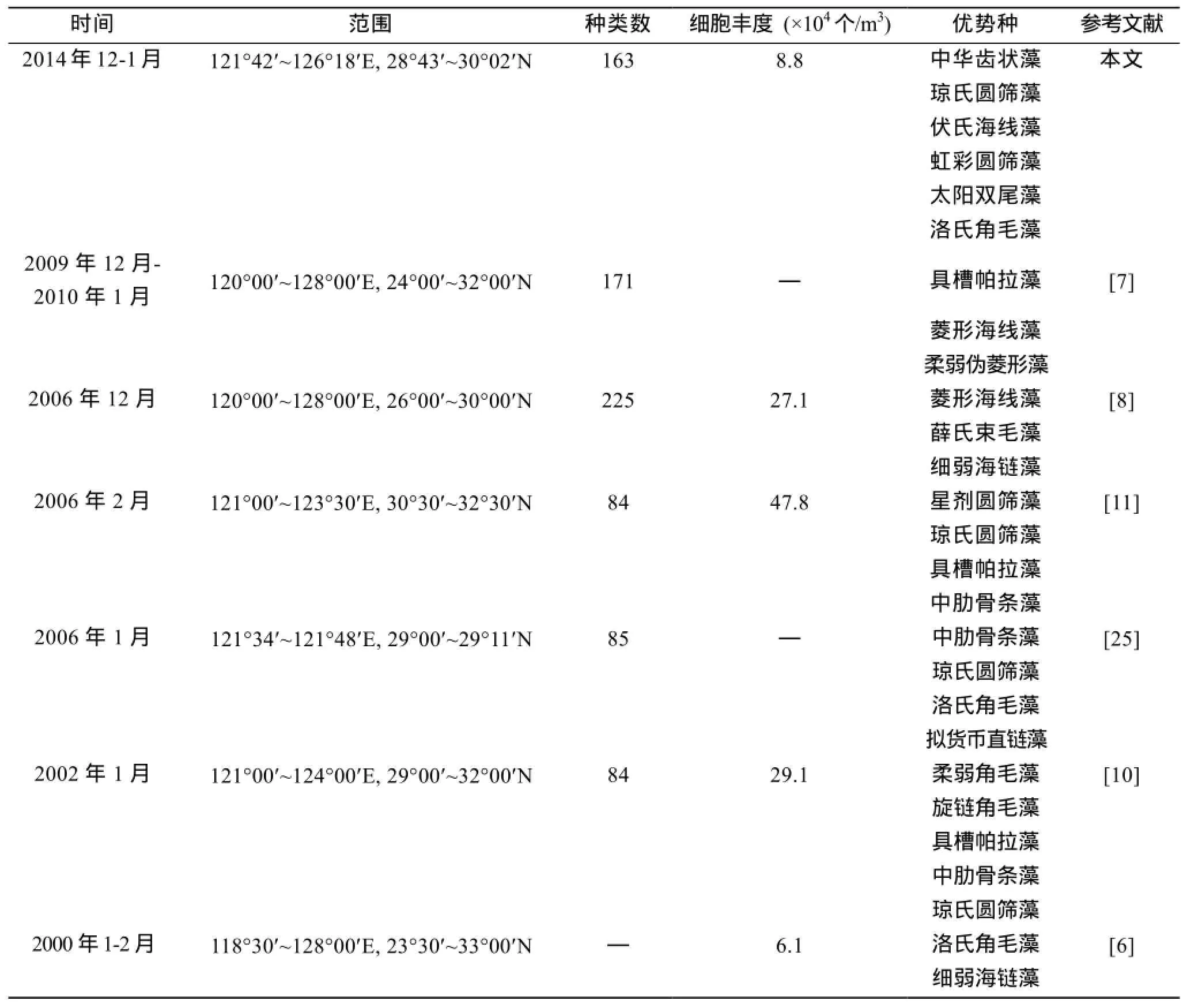
时间 范围 种类数2014年12-1月 121°42'~126°18'E,28°43'~30°02'N 163 2009年12月-2010年1月 120°00'~128°00'E,24°00'~32°00'N 171 2006年12月 120°00'~128°00'E,26°00'~30°00'N 225 2006年2月 121°00'~123°30'E,30°30'~32°30'N 84 2006年1月 121°34'~121°48'E,29°00'~29°11'N 85 2002年1月 121°00'~124°00'E,29°00'~32°00'N 84 2000年1-2月 118°30'~128°00'E,23°30'~33°00'N —细胞丰度 (×104个/m3) 优势种 参考文献8.8 中华齿状藻 本文琼氏圆筛藻伏氏海线藻虹彩圆筛藻太阳双尾藻洛氏角毛藻 — 具槽帕拉藻 [7]菱形海线藻柔弱伪菱形藻27.1 菱形海线藻 [8]薛氏束毛藻细弱海链藻47.8 星剂圆筛藻 [11]琼氏圆筛藻具槽帕拉藻中肋骨条藻 — 中肋骨条藻 [25]琼氏圆筛藻洛氏角毛藻拟货币直链藻29.1 柔弱角毛藻 [10]旋链角毛藻具槽帕拉藻中肋骨条藻琼氏圆筛藻6.1 洛氏角毛藻 [6]细弱海链藻
2.3 生物多样性
冬季浮游植物生物多样性指数的变化范围为1.72~4.72,均值为 3.36,本次调查海域浮游植物生物多样性指数较高,这主要因为调查海域受到的水团影响更加复杂,受到长江冲淡水、浙闽沿岸流以及黑潮、台湾暖流的影响,浮游植物的生态类型比较丰富。同样以经度为界,把调查海域分成5个区块,浮游植物多样性指数的区域差别较小,浮游植物多样性指数的高值区在122°50′~123°00′E,29°00′~29°15′N之间的海域,即渔山列岛以东约 50海里海域。孙军[24]在琉球群岛附近海域的调查研究表明,浮游硅藻生物多样性指数在3.4~3.6之间。外海的大洋水系,环境条件基本稳定,浮游植物多样性指数波动较小; 近海,受到沿岸水系的影响大,多样性指数的波动较大。
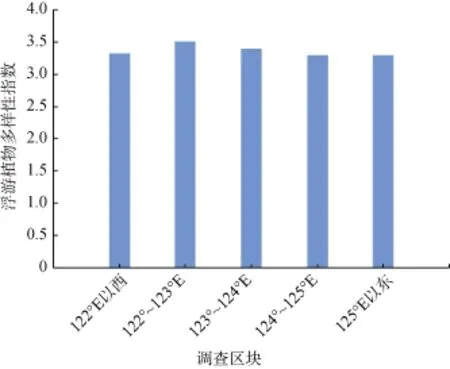
图3 调查海域浮游植物多样性指数Fig. 3 Shannon-Wiener diversity index in the survey area
2.4 浮游植物和环境因子的 Pearson相关性分析
浮游植物的生长和光照、温度、盐度、营养盐等环境因子息息相关,本文应用SPSS11. 0对冬季浙江中部海域浮游植物细胞丰度与环境因子进行Pearson相关性分析(表3)可以看出,浮游植物、硅藻、甲藻的细胞丰度和盐度、磷酸盐、硝酸盐相关性明显,其中和盐度呈明显负相关; 和磷酸盐、硝酸盐成明显正相关,沿岸及其近海海域盐度较小,磷酸盐和硝酸盐的浓度较高,为浮游植物的生长提供了充足的物质基础,利于浮游植物的生长; 甲藻和硅藻的细胞丰度和铵盐呈负相关,浮游植物和亚硝酸盐的相关性不明显。主要优势种中华齿状藻和温度成负相关,和盐度呈负相关,和磷酸盐、硝酸盐成明显正相关。

表3 细胞丰度和主要环境因子的Pearson相关性分析Tab. 3 Pearson correlation analysis between phytoplankton cell abundance and environmental factors
郭术津[7]、何青[11]、王丹[23]等对东海区浮游植物与环境因子的 Pearson相关性分析的研究均表明:浮游植物的细胞丰度和磷酸盐呈正相关。罗民波[6]对东海浮游植物与环境因子的多元回归分析表明:温度是影响东海四季浮游植物分布的主要影响因子,盐度是影响秋季浮游植物分布的主要影响因子。
3 结论
2014年冬季浙江中部海域浮游植物调查,共发现浮游植物 163种,其中以硅藻为主,甲藻其次。浮游植物优势种为中华齿状藻、琼氏圆筛藻、伏氏海线藻、虹彩圆筛藻、太阳双尾藻、洛氏角毛藻。最主要生态类型为近海广温型类群,半咸水类型浮游植物主要出现在三门湾至象山石浦沿岸,高温高盐性浮游植物类群在外海站位常见,其中主要代表种为中华齿状藻热带型、鸟尾藻、束毛藻等,可作为暖流指示种。从鸟尾藻的地理分布特点分析,2014年冬季黑潮入侵陆架区的现象依然存在且有所加剧。细胞丰度平均值为8.8×104个/m3,分布趋势呈现近岸>近海>外海的规律。细胞丰度的高值区在三门湾外湾至渔山列岛之间的海域。浮游植物生物多样性指数平均值为3.36,浮游植物的生态类型较为丰富。浮游植物的细胞丰度与磷酸盐、硝酸盐的浓度呈明显正相关,与盐度呈明显负相关。
参考文献:
[1] 谭书杰,宫相忠,孙军,等. 春节东海产卵场及其邻近海域的浮游植物群落[J]. 海洋科学,2009,33(8): 5-10. Tan Shujie,Gong Xiangzhong,Sun Jun,et al. The phytoplankton community in spawning ground of the East China Sea and its adjacent waters in spring[J]. Marine Sciences,2009,33(8): 5-10.
[2] 孙军,刘东艳,杨世民,等. 渤海中部和渤海海峡及邻近海域浮游植物群落结构的初步研究[J]. 海洋与湖沼,2002,33(5): 461-471. Sun Jun,Liu Dongyan,Yang Shimin,et al. The preliminary study on phytoplankton community structure in the central Bohai and Bohai Strait and Its adjacent area[J]. Oceanologia et Limnologia Sinica,2002,33(5):461-471.
[3] 郭玉洁,杨则禹. 1976年夏季东海陆架区浮游植物生态的研究[J]. 海洋科学集刊,1982,19: 11-32. Guo Yujie,Yang Zeyu. The Study of Phytoplankton ecology on the continental shelf of the East China Sea in summer 1976[J]. Studia Marina Sinica,1982,19: 11-32.
[4] 郭玉洁,杨则禹. 长江口浮游植物的数量变动及生态分析[J]. 海洋科学集刊,1992,33: 167-189. Guo Yujie,Yang Zeyu.The analysis of number variation and ecology of phytoplankton in the Changjiang estuary[J]. Studia Marina Sinica,1992,33: 167-189.
[5] GB/T12763-2007,海洋调查规范[S]. GB/T12763-2007,Specifications for oceanographic survey[S].
[6] 罗民波,陆健健,王云龙,等. 东海浮游植物数量分布与优势种[J]. 生态学报,2007,27(12): 5066-5085. Luo Minbo,Lu Jianjian,Wang Yunlong,et al. Horizontal distribution and dominant species of phytoplankton in the East China Sea[J]. Acta Ecologica Sinica,2007,27(12): 5066-5085.
[7] 郭术津,孙军,戴民汉,等. 2009 年冬季东海浮游植物群集[J]. 生态学报,2012,32( 10) : 3266-3278. Guo Shujin,Sun Jun,Dai Minhan,et al. Phytoplankton assemblages in East China Sea in winter in 2009[J]. Acta Ecologica Sinica,2012,32( 10) : 3266-3278.
[8] 秦玉涛,宋晨瑶,孙亚伟,等. 浙江海域夏冬两季浮游植物分布现状[C]//黄秀清.东海海洋环境调查与研究论文集,北京: 海洋出版社,2010,75-82. Qin Yutao,Song Chenyao,Sun Yawei,et al. Distribution of phytoplankton in Zhejiang in summer and winter[C]//Huang X Q. Investigation and research of marine environmental in the East China Sea,Beijing,Ocean Press,2010,75-82.
[9] 钟惠英. 三门湾健跳港浮游植物初探[J]. 浙江水产学院学报,1990,9(2): 126-130. Zhong Huiying. Preliminary study on the phytoplankton of Jian Tiao Bay of Zhangjiang[J]. Journal of Zhejiang college of fisheries,1990,9(2): 126-130.
[10] 吴玉霖,傅月娜,张永山,等. 长江口海域浮游植物分布及其与径流的关系[J]. 海洋与湖沼,2004,35(3):246-251. Wu Yulin,Fu Yuena,Zhang Yongshan,et al. Phytoplankton on distribution and its relation to the runoff in the Changjiang(Yangzte) estuary[J]. Oceanologia et Limnologia Sinica,2004,35(3): 246-251.
[11] 何青,孙军. 长江口及其邻近水域网采浮游植物群落[J].生态学报,2009,29(7): 3928-3938. He Qing,Sun Jun. The Netz-phytoplankton community in Changjiang(Yangzte) estuary.[J]. Acta Ecologica Sinica,2009,29(7): 3928-3938.
[12] 陆斗定,张志道. 浙江马鞍列岛附近海域浮游植物与赤潮生物研究[J]. 东海海洋,1996,14(1): 44-51. Lu Douding,Zhang Zhidao. Studies on Phytoplankton and red tide causative species in the sea area near Ma’an Islands of Zhejiang[J]. Donghai Marine Science,1996,14(1): 44-51.
[13] 陆斗定,蒋加伦,徐芝敏. 1986年春季东海黑潮区浮游甲藻种类组成及其分布特征的初步分析[C]//国家海洋局科技司. 黑潮调查研究论文选(一). 北京: 海洋出版社,1990: 229-238. Lu Douding,Jiang Jialun,Xu Zhimin. Preliminary analysis of composition and distribution of Dinoflagellates in the East China Sea in spring 1986[C]// Department of Science and Technology,State Oceanic Administration. Proceedings of investigation in the Kuroshio(NO.1),Beijing: Ocean Press,1990: 229-238.
[14] 陆斗定,徐芝敏,蒋加伦. 1986年日本南部浮游甲藻群落组成特点[C]//国家海洋局科技司. 黑潮调查研究论文选(二). 北京: 海洋出版社,1990,278-283. Lu Douding,Xu Zhimin,Jiang Jialun. Community composition of Dinoflagellate in the southern part of Japan in 1986[C]//Department of Science and Technology,State Oceanic Administration. Proceedings of investigation in the Kuroshio(NO.2),Beijing: Ocean Press,1990: 278-283.
[15] 陆斗定. 东海黑潮指示性甲藻的分布特征[C]//国家海洋局科技司. 黑潮调查研究论文选(三). 北京: 海洋出版社,1991,287-296. Lu Douding. Distribution of indicative Dinoflagellate in the Kuroshio area of the East China Sea[C]// Department of Science and Technology,State Oceanic Administration. Proceedings of investigation in the Kuroshio(NO.3). Beijing: Ocean Press,1991: 287-296.
[16] 郭玉洁,钱树本. 中国海藻志第五卷硅藻门第一册中心纲[M]. 北京: 海洋出版社,2003: 389-391. Guo Yujie,Qian Shuben. Flora Algarum Marinarum Sinicarum,TomusV Bacillariophyta NO.1 Centricae[M]. Beijing: Science Press,2003: 389-391.
[17] 刘东艳,孙军,钱树本.琉球群岛及其邻近海域的浮游甲藻—1997年夏季的种类组成和丰度分布[C]//国家海洋局科技司. 中国海洋学文集,2000,12: 170-182. Liu Dongyan,Sun Jun,Qian Shuben. Planktonic Dinoflagellates in Ryukyu-Gunto and Its adjacent waters species composition and their abundance distribution in the summer of 1997[C]//Department of Science and Technology,State Oceanic Administration.Chinese Marine science,Beijing: Ocean Press,2000,12: 170-182.
[18] 陆斗定,Jeanette G. 东海陆架黑潮区鸟尾藻的分类及其生态分布特点[J]. 东海海洋,2001,19(3): 11-18. Lu Douding,Jeanette G. Study on taxonomy and distribution of Ornithocercus in the East China Sea[J]. Donghai Marine Science,2001,19(3): 11-18.
[19] 潘玉球,苏纪兰,苏玉芬.东海南部水文的季节特性[C]//国家海洋局科技司. 黑潮调查研究论文选(五).北京: 海洋出版社,1993: 186-200. Pan Yuqiu,Su Jilan,Su Yufen. Seasonal characteristics of Hydrology in the south of the East China Sea[C]//Department of Science and Technology,State Oceanic Administration,Proceedings of investigation in the Kuroshio(NO.5). Beijing: Ocean Press,1993: 186-200.
[20] Marumo R,Asaoka O. Distribution of pelagic blue-green algae in the North Pacific Oeean.[J]. Oceanogr·Soc·Jap·,1974,30,77~85.
[21] Marumo R,Asaoka O. Trichodesmium in the East China sea 1.Distribution of Trichodesmium thiebautii Gomont during 1961-1967[J]. Oceanogr Soc Jap,1974,30: 289-303.
[22] 杨清良. 南黄海和东海陆架区束毛藻(Trichodesmium)的分布特征[J]. 海洋学报,1998,20(5): 93-100. Yang Qingliang.Characteristics of Trichodesmium distribution in waters over continental shelves of the South Hunaghai SeaAnd the East China Sea[J]. Acta Oceanologia Sinica,1998,20(5): 93-100.
[23] 徐韧. 浙江及福建北部海域环境调查与研究[M]. 北京: 科学出版社,2014: 324. Xu Ren.Investigation and Research on the marine environment of Zhejiang and northern waters of Fujian[M]. Beijing: Science Press,2014: 324.
[24] 孙军,刘东艳,钱树本.琉球群岛及其邻近海域的浮游硅藻—1997年夏季的种类组成和丰度分布[C]//国家海洋局科技司. 中国海洋学文集,北京: 海洋出版社,2000,12: 158-169. Sun Jun ,Liu Dongyan,Qian Shuben. Planktonic Diatoms in Ryukyu-Gunto and Its adjacent waters species composition and their abundance distribution in the summer of 1997[C]//Chinese Marine science,State Oceanic Administration,Beijing: Ocean Press,2000,12:158-169.
[25] 朱根海,陈立红,钱健. 三门湾核电站附近海域浮游植物季节变化研究[J],环境科学与技术,2010,33(12):36-45. Zhu Genhai,Chen Lihong,Qian Jian. A Study on Seasonal Viriation of Phytoplankton in the Sea Near Sanmen Nuclear Power Station[J]. Environmental Science and Technology,2010,33(12): 36-45.
[26] 王金辉. 长江口邻近水域的赤潮生物[J]. 海洋环境科学,2002,21(2): 37-41. Wang Jinhui.HAB alga nearby Changjiang Estuary[J]. Marine Environmental Science ,2002,21(2): 37-41.
[27] 王丹,孙军,安佰正,等. 2006年秋季东海陆架浮游植物群集[J]. 应用生态学报,2008,19(11): 2435-2442. Wang Dan,Sun Jun,An Baizheng,et al. Phytoplankton assemblages on the continental shelf of East China Sea in autumn 2006[J]. Chinese Journal of Applied Ecology,2008,19(11): 2435-2442.
[28] 孙军,刘东艳. 中国海区常见浮游植物种名更改初步意见[J]. 海洋与湖沼,2002,33(3): 271-286. Sun Jun,Liu Dongyan.The preliminary notion on nomenclature of common phytoplankton in China Sea waters[J]. Oceanologia et Limnologia Sinica,2002,33(3): 271-286.
(本文编辑: 康亦兼)
中图分类号:X826
文献标识码:A
文章编号:1000-3096(2016)04-0046-08
doi:10.11759/hykx20150518003
收稿日期:2015-05-18; 修回日期: 2015-07-28
基金项目:浙江省海洋环保与生态项目(浙海渔计[2012]100号)
作者简介:刘亚林(1982-),男,山东曲阜人,高级工程师,硕士,主要从事海洋浮游植物生态学研究,电话0577-28777166,E-mail: liuyalin@eastsea.gov.cnIII型浮游生物网(水深小于30 m使用,网具规格为网口内径37 cm,网长140 cm,筛绢孔径0.077 mm)。采样方式为从底至表垂直拖网,加中性甲醛溶液固定。
Phytoplankton assemblages in offshore water in mid-Zhejiang province during the winter of 2014
LIU Ya-lin1, HUANG Xiu-qing2, JIANG Xiao-shan1, WANG Hang-jun1, ZOU Qing1,QI An-xiang3
(1. Wenzhou marine environmental monitoring center,State Oceanic Administration,Wenzhou 325000,China;2. Shanghai East Sea Marine Engineering Survey & Design Institute,Shanghai 200137,China. 3. East China Sea Forecast Center,State Oceanic Administration,Shanghai 200137,China)
Received: May 18,2015
Key words:offshore water of mid-Zhejiang province; winter; phytoplankton; assemblage structure
Abstract:A total of 97 phytoplankton samples were collected in a multidisciplinary investigation from offshore waters in mid-Zhejiang province (28°43'-30°02'N,121°42'-126°18'E) from December 2014 to January 2015. Moreover,the phytoplankton species composition,assemblage structure,and cell abundance of phytoplankton were studied. A total of 163 species belonging to 53 genera of 6 phyla were identified (including uncertain species). The phytoplankton community mainly comprised bacillariophyta and dinophyta. Moreover,in the study area,the phytoplankton community was divided into four ecological types: a true estuarine component,coastal component,eurythermal and euryhaline marine component,and a high salinity component. of these,the eurythermal and euryhaline marine component was the major ecotype. Dominant phytoplankton species were Odentella sinensis,Coscinodiscus jonesianus,Thalassiothrix frauenfeldii.,Coscinodiscus oculus-iridis,Ditylum sol,and Chaetoceros lorenzianus. Cell abundance ranged from 0.3×104cells/m3to 61.7×104cells/m3with an average of 8.8×104cells/m3. The higher values occurred in the area between Sanmen Bay and the Yushan Islands. The average Shannon-Wiener indices of phytoplankton assemblages was 3.36,and according to Pearson correlation analysis,the distribution of phytoplankton cell abundance was positively correlated with phosphate and nitrate but negatively correlated with salinity.
猜你喜欢
农民致富之友(2016年22期)2017-02-07
建筑建材装饰(2016年11期)2016-12-29
中国绿色画报(2016年7期)2016-12-26
天津农业科学(2015年6期)2015-06-16
