
商陆及其同源四倍体叶绿素荧光参数的日变化
2016-07-25 15:52陈梦华赵盈丽游少鸿刘杰
江苏农业科学 2016年6期
陈梦华++赵盈丽++游少鸿+++刘杰++杨明匀++何昌杰
摘要:为选育用于植物修复的更具优异光合特性的超富集植物,以商陆(Phytolacca acinosa)及其同源四倍体为研究对象,采用PAM-2500便携式调制叶绿素荧光仪测定其植株叶片的叶绿素荧光参数日变化。测定结果表明:初始荧光(Fo)、最大荧光产量(Fm′)、PSⅡ的最大光合量子产量(Fv/Fm)、PSⅡ的实际光合量子产量[Y(Ⅱ)]及通过PSⅡ的电子传递速率(ETR)均存在明显的日变化。其中Fm′、Fv/Fm、Y(Ⅱ)值随时间呈先降后升的变化趋势,最小值出现在12:00或14:00左右,至18:00时商陆及其同源四倍体Fv/Fm值分别恢复到08:00时的97.4%、98.8%;而Fo、ETR随着时间呈先升后降的变化规律,四倍体较二倍体ETR平均值高35.49%。综上,二者在强光下均表现出可逆的光抑制。商陆同源四倍体较二倍体具有更优良的光合特性,对强光环境表现出更强的耐性,用于锰污染植物修复领域更具研究价值。
关键词:商陆;四倍体;叶绿素荧光参数;日变化
中图分类号: X173;Q945.11文献标志码: A文章编号:1002-1302(2016)06-0278-03
收稿日期:2015-05-14
基金项目:国家自然科学基金(编号:41001186);广西自然科学基金(编号:2011GXNSFF018003、2013GXNSFBA019210)。
作者简介:陈梦华(1989—),女,广东阳江人,硕士,研究方向为重金属污染修复。E-mail:chanmenghua@163.com。
通信作者:游少鸿,副教授,研究方向为重金属污染修复。E-mail:646761963@qq.com。植物生长基于光合作用,其光合作用能力的高低直接决定植物的总生长力。植物体内发出的叶绿素荧光与光合作用的反应过程紧密相关[1],通过叶绿素荧光分析技术测得的叶绿素荧光参数是研究植物光合特性的重要指标。目前,叶绿素荧光分析技术已经成为研究植物光合作用与环境关系以及抗逆生理的重要手段[2-4]。随着社会的发展和科技的进步,多倍体育种技术广泛应用于培育观赏性更强的花卉[5]、产量更高的农作物[6-7]及药用成分含量更高的药材[8]。植物倍性差异导致生理特性的改变,随之产生不同的生理生化现象[9]。当今重金属污染日渐严重,关于植物修复更具优异特性的超富集植物的研究迫在眉睫。商陆(Phytolacca acinosa)作为药用植物及重金属锰超富集植物,其同源四倍体(4x)的诱导研究已见报道[10],并已初步证实四倍体具有修复优势。但是其二倍体(2x)及四倍体叶绿素荧光参数的相关研究未见报道。
本研究以商陆二倍体和经秋水仙素诱导所得商陆同源四倍体为材料,对二者叶绿素荧光参数Fo、Fm′、Fv/Fm、Y(Ⅱ)、ETR的日变化进行了比较研究。为进一步深入探讨商陆同源多倍体特性提供依据,并为商陆同源四倍体作为锰超富集植物提供参考。
1材料与方法
1.1试验材料
试验于2014年6月于桂林理工大学重金属污染植物修复试验基地进行,供试材料为试验基地内培植的商陆以及经秋水仙素诱导所得商陆同源四倍体。
1.2试验方法
选取长势一致的商陆二倍体、四倍体植株各5株为试验材料,每株取全光照环境下高度和方位一致的生长健壮的中部功能叶1张,标记并于每张叶片中部大约相同位置(避开叶脉)进行活体测试。采用便携式调制叶绿素荧光仪(PAM-2500,泽泉科技有限公司&泽泉生态开放实验室)于晴天测定各选定叶片的叶绿素荧光参数的日变化(08:00—18:00),每2 h测定1次,其中叶夹测量的光合有效辐射在 1 200~1 300 μmol/(m2·s)范围内,温度为(31±2) ℃。测定前用特定夹子将各选定叶片暗适应30 min,而后将夹子对准探头拉下金属遮光片使叶片暴露在饱和脉冲光下1 s,从仪器中直接读取初始荧光(Fo)、最大荧光(Fm)等值。2个反映光系统Ⅱ(PSⅡ)光能利用效率的参数计算如下:
PSⅡ的最大光合量子产量(反映了样品的光合潜能):
Fv/Fm=(Fm-Fo)/Fm;
PSⅡ的实际光合量子产量:
Y(Ⅱ)=(Fm′-F)/Fm′。
1.3数据分析
数据采用Excel 2003和DPS v7.55软件进行分析, 采用Duncans新复极差法进行显著性检验。
2结果与分析
2.1光合有效辐射(PAR)的日变化
由表1可知,1 d中的光合有效辐射从08:00开始逐渐上升,12:00达到1 d中的最大值,为1 299.83 μmol/(m2·s),为最初时的1.03倍,18:00 时降至1 189.50 μmol/(m2·s)。
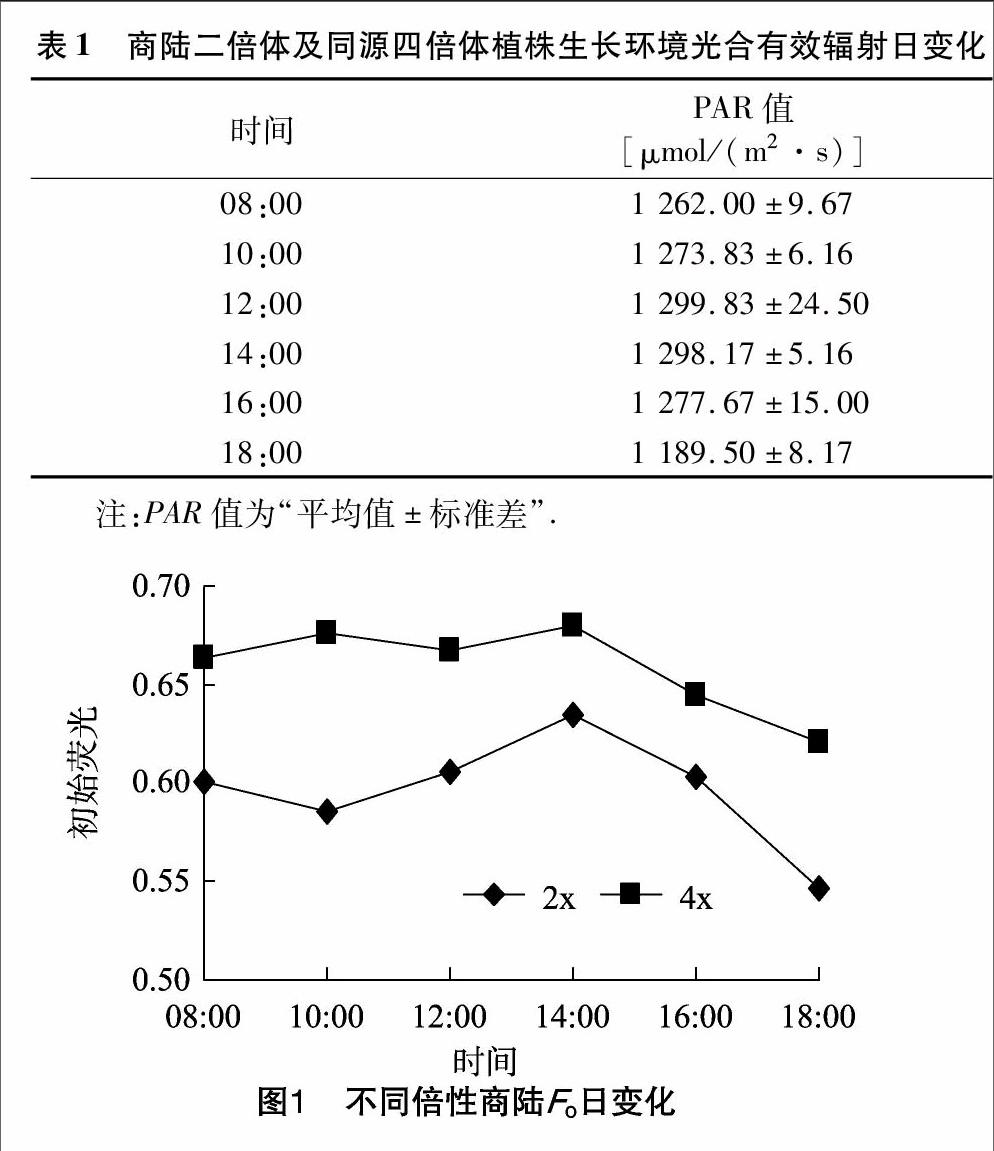
2.2初始荧光的日变化
初始荧光是光系统Ⅱ反应中心全部开放时的荧光水平[11]。PSⅡ天线的热耗散增加常导致Fo降低,而PSⅡ反应中心的破坏或可逆失活则引起Fo增高,因此可根据Fo的变化推测反应中心的状况和可能的光保护机制。商陆二倍体和四倍体Fo的日变化整体上均呈先上升后下降的趋势(图1)。从08:00—12:00,二者Fo略有下降,显示在上述条件下的光抑制并非是叶片光合机构受到强光破坏的结果,而是一种光保护作用的调节机制。14:00时,商陆二倍体和四倍体Fo均达到峰值,分别为0.634、0.680,表明光抑制是叶片光合机构受到强光破坏或可逆失活的结果。商陆四倍体的平均Fo值为二倍体的1.11倍,表明二者Fo的日变化差异并不大。
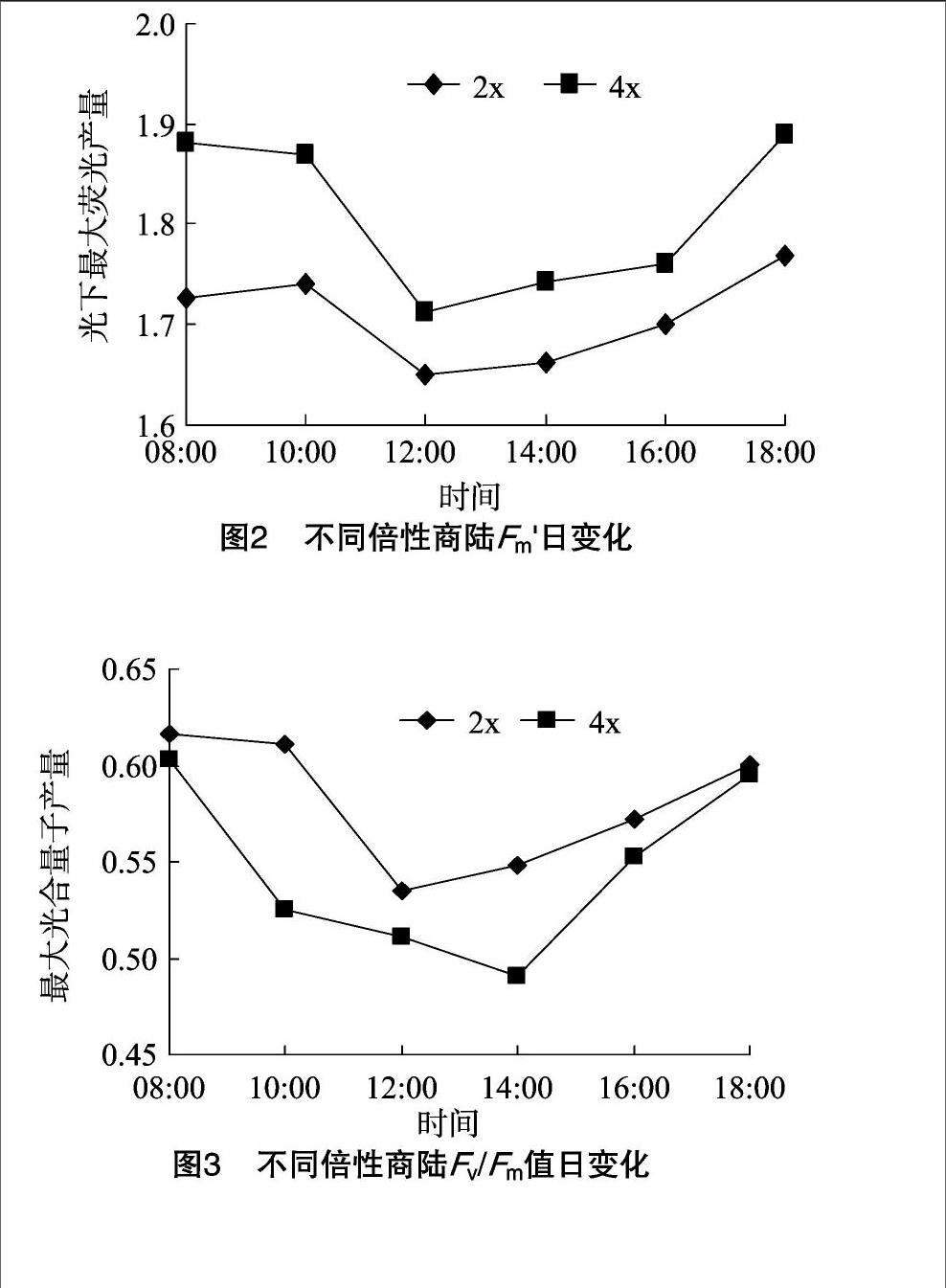
2.3最大荧光产量的日变化
光下最大荧光产量是光适应的植株在打开饱和脉冲时得到的最大荧光产量,反映自然状况下植物体光合机构开放程度[12]。商陆二倍体和四倍体Fm′的日变化大体上均呈先下降后上升的变化规律(图2)。二者Fm′均在12:00降到最低,分别为1.651、1.712。08:00—12:00,随光照强度的不断增强,引起了电子在光系统Ⅰ和Ⅱ之间累积,叶绿素分子得到的量子减少,Fm′不断下降,12:00降到最小值,表明其在午间均出现了光抑制。12:00—18:00,二者Fm′又逐渐上升,说明午间强光并未造成其 PSⅡ反应中心受到破坏,表现出植物在长期生境中形成的一种自我保护机制。不同倍性商陆Fm′的日变化幅度不同,18:00时Fm′恢复到初始值的程度也不同,四倍体的下降及恢复的幅度均比较大;而二倍体光抑制现象后该值回升但幅度较缓,表明二倍体叶片光合机构受强光损伤后恢复较慢,与不同植物受到的光抑制不同,自我保护抑制也存在差异有关。
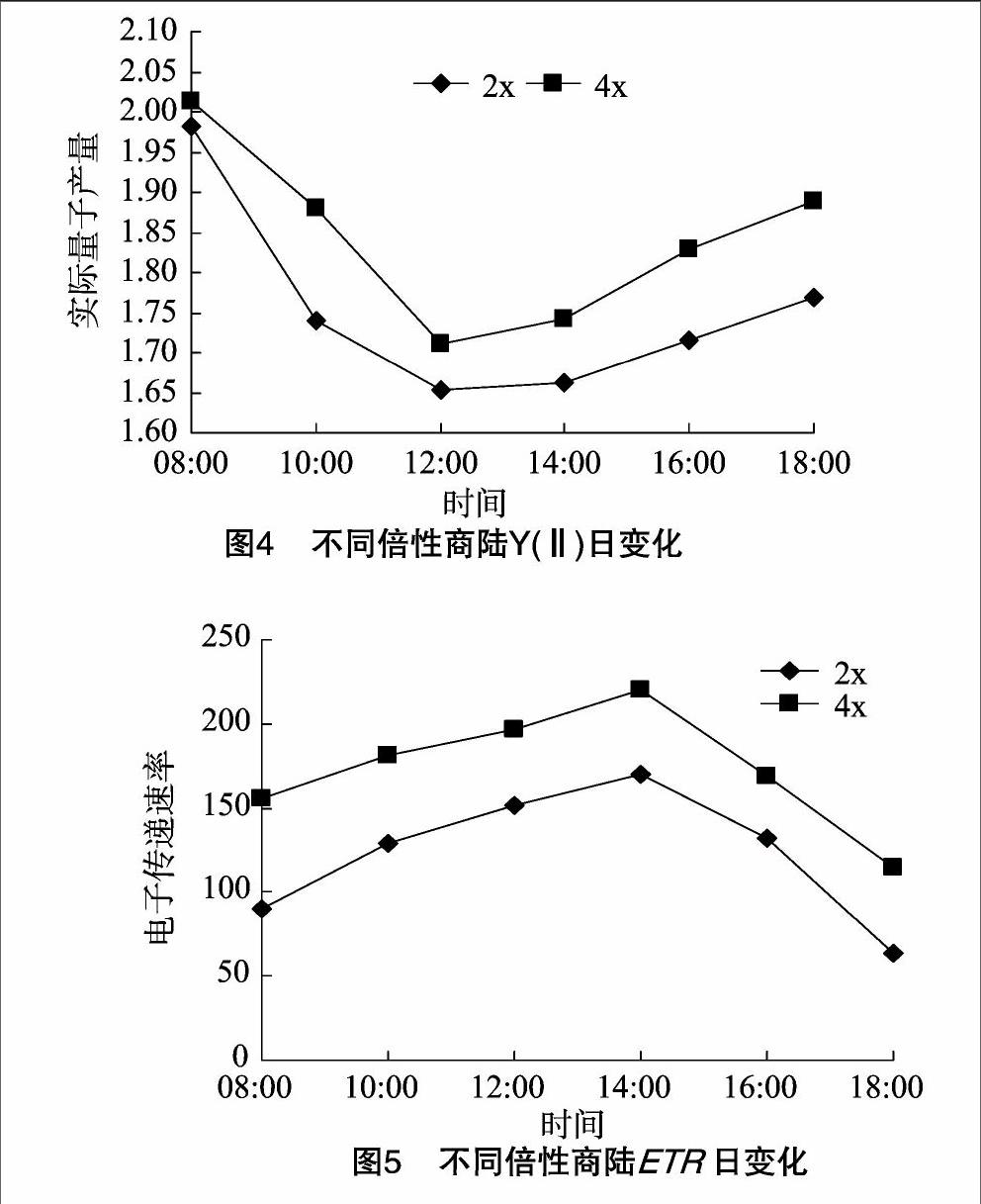
2.4最大光合量子产量的日变化
PSⅡ的最大光合量子产量变化是研究最为广泛的光抑制指标。从图3可以看出,商陆二倍体、四倍体叶片Fv/Fm值的日变化大体上均呈先降后升的趋势。8:00时Fv/Fm值较高,随着光照度及温度升高,二倍体、四倍体Fv/Fm值不同程度地下降,并先后在12:00、14:00左右达到谷底,最小值分别为0.535、0.491,均明显低于各自08:00的Fv/Fm值。表明在强光、高温下叶片PSⅡ光化学效率降低,并出现光合作用的光抑制。二者Fv/Fm值波谷的降幅及持续时间差别各异,恢复幅度及快慢程度也不尽相同,相对而言,四倍体Fv/Fm值恢复幅度大、速率快、所需时间短。至18:00时,二倍体、四倍体分别恢复至08:00时Fv/Fm值的97.4%%、98.8%,说明商陆四倍体叶片在弱光下恢复速率和程度及其PSⅡ功能的可逆效果都优于二倍体,对避免强光伤害所作出的反应也更加灵敏。Fv/Fm值逐渐回升到初始水平,表明中午PSⅡ的功能下降是可逆的,PSⅡ的功能下降可能是避免中午光强伤害的一种适应方式。
2.5实际光化学量子产量的日变化
实际光化学量子产量是指植物光合作用下PSⅡ总的光化学量子产量,它反映PSⅡ反应中心在部分关闭情况下的实际原初光能捕获效率[13]。Y(Ⅱ)值较高更有利于光能转化效率的提高,同时可为暗反应中的光合碳同化积累更多的能量,以便促进碳同化作用的高效运转和有机物的大量积累(图4)[14]。商陆二倍体、四倍体的PSⅡ的实际量子产量Y(Ⅱ)日变化趋势与Fv/Fm值基本相似,均呈现出先下降然后逐渐上升的倒抛物线型变化趋势,二倍体、四倍体的Y(Ⅱ)最小值均出现在12:00,说明光合机构在强光和高温下其光合活性受到了暂时的抑制。在1 d中相同时刻二倍体的Y(Ⅱ)值均低于四倍体,说明四倍体的光能转化率较高。对比二倍体及四倍体Y(Ⅱ)的降幅,四倍体总体降幅较小,为149%,进一步证明四倍体较好的光合能力。与 Fv/Fm值相比,Y(Ⅱ) 1 d中变化幅度更大,表明Y(Ⅱ)指标对外界光强和气温条件的响应比 Fv/Fm值敏感。
2.6电子传递速率的日变化
通过PSⅡ的最大电子传递速率,是光合机构吸收光能发生电荷分离产生电子,并沿电子传递链向下传递的速率。其高低在一定程度上反映了PSⅡ反应中心的电子捕获效率的高低[15]。从图5可以看出,商陆二倍体、四倍体ETR均表现出先升高后下降的趋势。二者ETR峰值均出现在14:00左右,分别为170、220,与初始值相比其增幅分别达到了889%、41.0%。正午后ETR开始缓慢降低,在18:00达到最低值。1 d中四倍体植株ETR各个时间点的值均相对高于二倍体约35.49%,说明商陆四倍体PSⅡ反应中心的电子捕获效率较高,在一定程度上反映商陆四倍体光合能力较强,能将更多吸收的光能用于光合作用,减少了多余电子的产生,对光合器官起到一定的保护作用,即商陆四倍体的电子传递速率高于商陆二倍体。
3结论与讨论
商陆二倍体及同源四倍体Fo和ETR日变化趋势基本一致,二者均随着时间变化呈先升高后降低的趋势。各测定时间点的四倍体日变化值均高于二倍体,表明商陆四倍体光系统Ⅱ反应中心的电子捕获效率较二倍体高,通过光耗散而降低光抑制的能力强,反映其在强光下能保持较高光合作用效率,更耐光照较强的环境。
商陆二倍体及其同源四倍体的光下最大荧光产量、PSⅡ的最大光合量子产量和PSⅡ的实际光合量子产量均随着时间变化呈先降后升趋势。表明二者均发生了光抑制现象,在日变化过程中商陆四倍体较二倍体Fv/Fm值下降快,受光抑制影响、光合机构受损伤也都比较大,但午后该值又急剧回升,表现其极强的损伤恢复能力,至18:00时商陆及其同源四倍体Fv/Fm值分别恢复到8:00时的97.4%、98.8%。四倍体Fm′、Y(Ⅱ)的变化趋势均较二倍体明显,体现出四倍体较二倍体在自然状况下光合机构开放程度更大、更易于提高光能转化效率。
商陆二倍体及其同源四倍体的各荧光参数的最高、最低值并非全部出现在PAR最高值时的12:00,部分出现在同样光照度较强的14:00,说明植物对强光响应有一定的反应时间。不同倍性商陆的叶绿素荧光参数之间也存在一定差异,商陆同源四倍体较其二倍体表现出更优良的光合特性,用于锰污染植物修复领域更具研究价值。
参考文献:
[1]冯汉青,汤淑珍,李淮. 不同光强下交替氧化酶基因修饰的两种拟南芥叶片叶绿素荧光特性的变化[J]. 广西植物,2013,33(5):679-684.
[2]吴韩英,寿森炎,朱祝军,等. 高温胁迫对甜椒光合作用和叶绿素荧光的影响[J]. 园艺学报,2001,28(6):517-521.
[3]白志英,李存东,赵金锋,等. 干旱胁迫对小麦代换系叶绿素荧光参数的影响及染色体效应初步分析[J]. 中国农业科学,2011,44(1):47-57.
[4]王振磊,陈海江,林敏娟,等. 黄金梨和鸭梨叶片光合作用的光抑制及其恢复的比较研究[J]. 园艺学报,2009,36(9):1261-1268.
[5]郭海滨. 花卉多倍体育种研究进展探析[J]. 绿色科技,2011(1):174-175,179.
[6]谢大森,张钢,彭庆务,等. 秋水仙素诱导冬瓜多倍体的初步研究[J]. 热带作物学报,2011,32(11):2094-2098.
[7]刘喜星,顾玉红,李保国,等. 核桃多倍体诱导技术研究[J]. 北方园艺,2012(2):113-116.
[8]杜艳伟,阎晓光,赵晋锋. 药用植物多倍体育种的研究进展[J]. 生物技术进展,2011,1(4):249-253.
[9]Peng H,Zhang J,Wu X. The ploidy effects in plant gene expression:progress,problems and prospects[J]. Science in China Series C-Life Sciences,2008,51(4):295-301.
[10]常建忠,吴玉香,赵晓明,等. 秋水仙素诱导美洲商陆四倍体的研究[J]. 中国农学通报,2009,25(4):35-38.
[11]李林光,李芳东,王颖,等. 苹果四倍体品种叶绿素荧光参数的日变化[J]. 果树学报,2011,28(1):129-133.
[12]朱琳,潘国富,杜广明,等. 呼伦贝尔草原三种优势植物叶绿素荧光特性的比较[J]. 中国草地学报,2012,34(4):82-86.
[13]李春霞,曹慧. 干旱对苹果属植物叶绿素荧光参数的影响[J]. 安徽农业科学,2008,36(31):13536-13538.
[14]吕芳德,徐德聪,侯红波,等. 5种红山茶叶绿素荧光特性的比较研究[J]. 经济林研究,2003,21(4):4-7.
[15]孙景宽,张文辉,陆兆华,等. 干旱胁迫下沙枣和孩儿拳头叶绿素荧光特性研究[J]. 植物研究,2009,256(2):216-223.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
大众健康(2022年7期)2022-07-30
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
石河子大学学报(自然科学版)(2021年6期)2021-03-03
现代园艺(2017年13期)2018-01-19
中成药(2016年4期)2016-05-17
首都食品与医药(2015年13期)2015-11-03
中国果业信息(2015年6期)2015-01-23
江苏中医药(2015年1期)2015-01-09
