
不同供硅条件下稗草生物量生殖分配及生殖分株数量特征
2016-07-25 15:07赵琦郭伟王存国杨颜熙陈熙
江苏农业科学 2016年6期
关键词:生物量
赵琦++郭伟+王存国++杨颜熙+++陈熙


摘要:采用盆栽试验方法分别测定5种供硅条件下稗草[Echinochloa crusglli (L.) beauv.]分蘖期、拔节期、盛花期、结实期各个器官的生物量以及株丛高、分株高、分蘖数等指标。结果表明:不同供硅条件下,分蘖期稗草各个器官的生物量、生物量分配之间差异基本不显著;拔节期施硅量为300 kg/hm2时,根、叶、叶鞘、全株生物量显著低于对照,根生物量分配呈先下降后上升的趋势,叶生物量分配先上升后下降;盛花期和结实期施硅量为240 kg/hm2时,稗草总生物量最大,施硅量为300 kg/hm2时,稗草总生物量最小,不同硅处理下的根、叶、叶鞘生物量分配之间差异不显著;盛花期,施硅量为60、120 kg/hm2时,茎生物量分配较高,花生物量分配较低;结实期,在不施硅和中硅(240 kg/hm2)处理下,稗草有较高的果生物量分配,施硅量为(180 kg/hm2)时,果生物量分配较小,茎生物量分配较大。分蘖数和分株高随施硅量的增加呈先上升后下降的趋势,生殖分配(Ⅰ)与分株高呈显著负相关。
关键词:稗草;生物量;生物量分配;生殖分配
中图分类号: Q948.1;S451文献标志码: A文章编号:1002-1302(2016)06-0205-05
收稿日期:2016-02-18
基金项目:国家重大科技专项水专项(编号:2014ZX07201-009-04)。
作者简介:赵琦(1989—),女,山西忻州人,硕士研究生,研究方向为植物种群生态学。E-mail:18802485361@163.com。
通信作者:郭伟,副教授。E-mail:guowei_233@163.com。植物种群生态学是种群生态学研究的前沿,而种群的生殖分配是植物种群生态学研究中的一个重要方面[1-3],生殖分配(reproductive allocation,RA),亦称生殖配置,是指植物一年所同化的资源中用于生殖的比例,实际上指总资源供给生殖器官的比例,它控制着植物生殖和平衡[4]。生殖分配在国外已有40多年的研究历史[5],而国内起步较晚且研究大都集中在木本植物、草原等地区特有的草本植物,对农田环境条件下肥力对杂草生殖分配的研究较少[6-13],郭伟等在此方面作了初步探讨[14-15]。稗草原产于欧洲和亚洲,广泛分布于温带和亚热带,是禾本科稗属的一年生草本植物,是具有发达根系的无性系植物,作为稻田的主要伴生植物,稗草是全球分布最广的恶性杂草[16-21]。控制农田杂草的本质是控制农田杂草的生殖,因为生殖是生物繁衍后代及其延续种群最基本的行为和过程。硅是继氮、磷、钾之后的第4种营养元素,影响植物的生长发育和生殖分配格局,而目前硅的使用和研究大都集中在水稻,对其他植物研究较少[22-23]。因此研究不同外源硅浓度条件下稗草的生殖分配和生殖分株数量特征,既丰富了生殖分配的理论,又为稻田稗草的生态控制提供了理论依据。
1材料与方法
1.1试验材料
稗草:采自沈阳市周边稻田。
1.2试验方法
采用盆栽试验,盆直径30 cm、高26 cm,每盆1穴,硅施偏硅酸钠(以SiO2计),设5个处理:Si1,60 kg/hm2;Si2,120 kg/hm2;Si3,180 kg/hm2;Si4,240 kg/hm2;Si5,300 kg/hm2。溶入等体积的蒸馏水中,用稀硫酸调为中性,为了排除盐分的干扰,施硫酸钠作为对比试验,S1:137.2 kg/hm2;S2:277.4 kg/hm2;S3:414.7 kg/hm2;S4:553.4 kg/hm2;S5:692.1 kg/hm2。施氮肥(按纯量计)225 kg/hm2,磷肥(P2O5)79.5 kg/hm2,钾肥(K2O)63 kg/hm2,每个处理3次重复,同时设对照CK。
分别在分蘖期、拔节期、盛花期、结实期取样,记录株丛高、单株分蘖数,在结实期将采集的植株各分蘖株进行高度分级,测定每个高度级的平均质量,并将植株各器官分离,于105 ℃杀青30 min,80 ℃烘干至恒质量。计算公式:
生物量分配=某器官干质量+附属结构干质量植株总生物量干质量×100%;
生殖分配(Ⅰ)[RA(Ⅰ)]=某一高度级籽粒质量生殖分蘖株总质量×100%。
2结果与分析
2.1不同供硅条件下稗草生物量生殖分配
2.1.1不同供硅条件下稗草生物量从表1可以看出,不同供硅条件下,生殖器官生物量总体上呈增长趋势,稗草生物总量从分蘖期到盛花期迅速增长,盛花期稗草在生殖生长的同时还进行营养生长。结实期稗草营养生长停止,主要进行生殖生长。
由表1还可看出,随着施硅量的增加,分蘖期各处理下各个器官的生物量无显著差异。拔节期对照处理的根生物量显著高于其他处理;茎生物量呈先上升后下降的趋势,CK、Si5处理显著低于其他处理;叶、叶鞘、总生物量在Si5处理下较小,显著低于对照。说明拔节期高浓度硅对稗草生物量的积累起抑制作用。
此外,由表1可见,盛花期随施硅量的增加,根质量呈先增加后下降的趋势,叶鞘、花果的变化呈二次曲线。在Si5处理下,根、茎、叶、叶鞘、全株的生物量较小;在Si4处理下茎、叶、花和全株的生物量最大,且两者之间差异显著。结实期不同浓度硅处理下叶鞘的生物量变化呈二次曲线。在Si5处理下,根、茎、叶、叶鞘、花果的生物量最小,Si4处理下的茎、果、全株生物量最大,二者之间差异显著。说明进入生殖生长期后,Si4处理对稗草生物量的积累起促进作用,Si5处理对稗草的生物量积累起抑制作用。
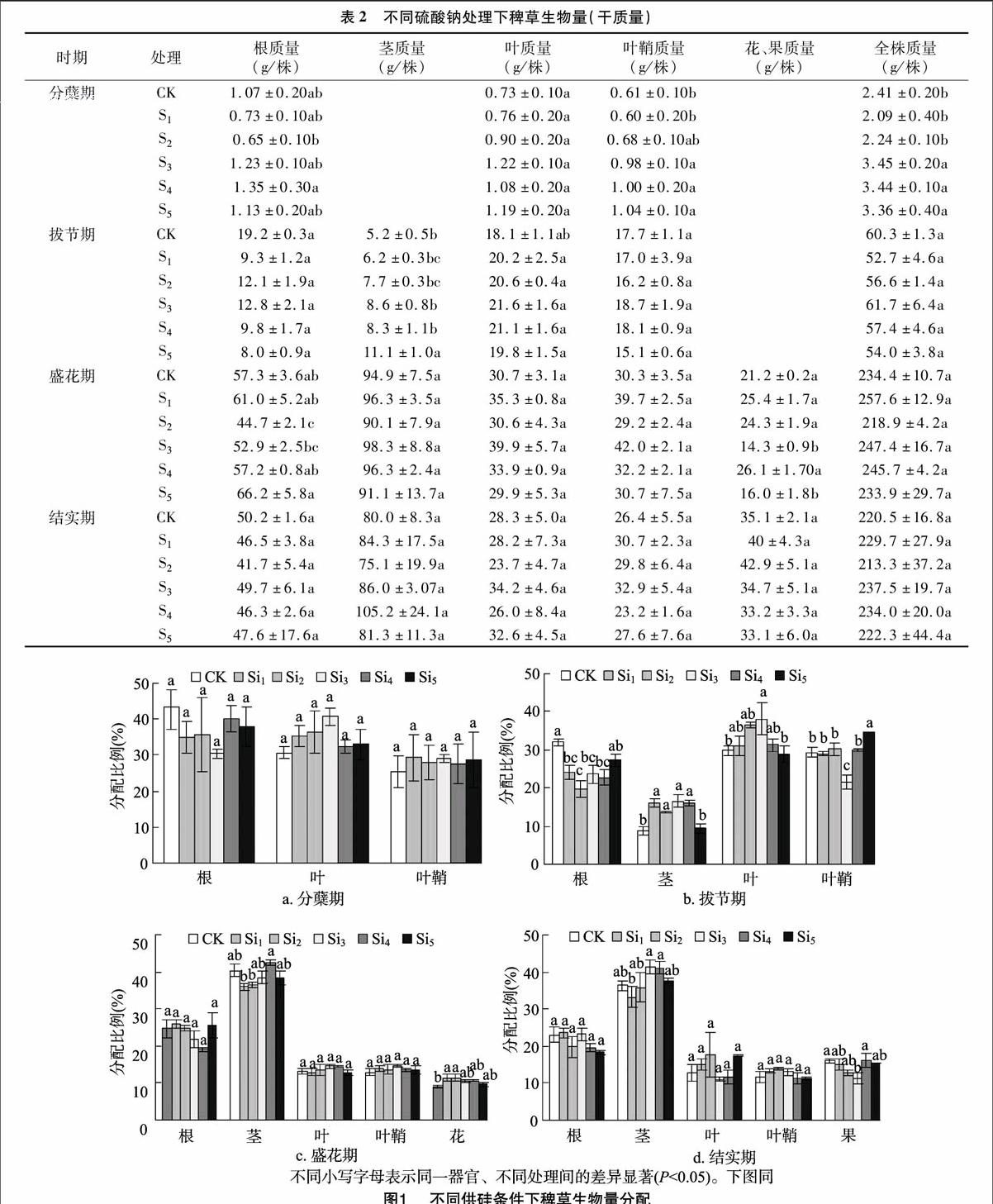
由表2可见,分蘖期不同浓度硫酸钠处理下的叶生物量无显著差异,S3、S4、S5处理下叶鞘、全株生物量显著高于对照。拔节期不同浓度硫酸钠处理下的根、叶、叶鞘、全株生物量之间无显著差异,S5处理下茎生物量最高且显著高于对照。
由表2还可见,盛花期不同浓度硫酸钠处理下的茎、叶、叶鞘、全株生物量之间差异不显著,S2处理下的根生物量最小,显著低于S5处理,S3处理下的花果生物量最小。结实期不同浓度硫酸钠处理下的各个器官之间差异不显著。
通过分析各个时期不同硅、硫酸钠处理下各个器官生物量动态变化发现,影响稗草生物量的是硅,并不是其中的盐分。
2.1.2不同供硅条件下稗草生物量分配生殖分配是由内在基因和外在环境决定的,同种植物在不同生境下会表现出不同的数量特征和生殖对策。从图1可以看出,分蘖期和拔节期叶占有较高的生物量分配比例,说明稗草在营养生长时期叶通过光合作用积累了大量的有机质,为稗草后期生长提供物质需要。稗草进入生殖生长时期后,茎生物量生殖分配明显增大,盛花期和结实期生物量分配大小为:茎>根>叶鞘≈叶,说明生长后期稗草利用根部贮藏的有机物质提供更多的物质供稗草进行生殖生长。随生长期的延长,生殖器官生物量分配增大,但总体来说低于20%,与已有一年生草本植物生殖器官的生物量分配的研究结果高于20%[24]不同,这可能与稗草生长的生态环境有关。
从图1可以看出,分蘖期各个器官生物量分配在各处理间差异不显著。拔节期根生物量分配呈现出先下降后上升的趋势,茎生物量分配在CK、Si5处理下显著低于其他处理。叶生物量分配呈现出先增加后降低的趋势,在Si3处理下显著高于对照和高硅处理Si5,中浓度的硅有助于增大叶面积,提高光合作用,积累有机质。
盛花期营养器官的生物量分配均高于生殖器官的生物量分配,说明这个时期资源大部分用于植物的营养生长而不是生殖生长,有助于提高稗草的竞争力[25]。根、叶、叶鞘生物量分配差异不显著,茎生物量分配在Si1、Si2处理下低于其他处理,花生物量分配在Si1、Si2处理下高于其他处理,并和对照处理差异显著(图1),说明在低硅处理下稗草为了提高潜在的生育率要提供更多的物质保证花的生长。
稗草生长进入结实期后,营养生长基本停止,资源主要用于生殖生长。由图1可见,根、叶、叶鞘生物量分配在各处理间差异不显著;在Si3、Si4处理下的茎生物量分配高于其他处理;果生物量分配在CK、Si4处理下高于其他处理,且显著高于Si3处理。在不施硅条件下稗草的生殖策略发生改变[26],给予生殖器官更多的生物量分配,保证了子代的繁殖。在Si3处理下,将较多的物质分配给茎,保证营养器官能量的吸收,维持稗草的生殖生长,提高稗草的竞争力,维持下一代的绝对数量。在Si4处理下,茎生物量分配和果生物量分配都较大,说明此浓度处理下的稗草最有利于种子的繁殖。
从图2可以看出,分蘖期在不同硫酸钠处理,稗草各个器官不同处理之间差异不显著;拔节期对照处理的根生物量分配比例显著高于其他处理,不同硫酸钠处理叶、叶鞘差异不显著,茎生物量分配呈上升趋势,在S5处理下达到最大值,与
其他处理之间差异显著。
由图2还可以看出,盛花期根生物量分配呈先下降后上升的趋势,S2处理下根生物量分配最低,显著低于S5处理;茎和叶生物量分配在各处理之间差异不显著;S3处理下的叶鞘生物量分配最高显著高于对照,S3处理下花生物量分配最低,显著低于对照。结实期根、茎、叶、叶鞘生物量分配在各处理间差异不显著,和硅处理下的果生物量分配不同,S2处理下果生物量分配的最高,S3处理下果生物量分配最低。硅处理、硫酸钠处理在分蘖期对稗草各个器官分配的影响都不显著。拔节期、盛花期和结实期硫酸钠和偏硅酸钠处理下的生物量分配不同。对生物量分配起作用的是硅,并不是其中的盐分。
2.2不同供硅条件下稗草数量特征和生殖分配
从表3可以看出,Si3、Si4处理下的株丛高大于低硅处理(CK、Si1、Si2)和高硅处理(Si5),但处理之间差异基本不显著。分蘖数随着施硅浓度的增加出现先增加后减少的趋势,与郭正刚等关于硅对紫花苜蓿分枝数影响的研究一致[27]。Si2和Si3处理下,千粒质量和分蘖数的变异系数高于其他处理,说明此浓度下的形态可塑性和生殖器官的可塑性都较大。
同时由表3可以看出,分株高随浓度的增加呈现先增加后降低的趋势,Si3处理下株高为最高,Si5处理下最低,二者之间差异显著;RA(Ⅰ)随浓度的变化出现先降低后升高的趋势,对株高和RA(Ⅰ)进行回归分析可知:y=-0.198x+44809,r2=0538 6,二者之间相关性显著,说明随着施硅浓度的变化,为了保证稗草的生存,生殖分株会对有限的资源进行调节,体现了生殖分株对有限资源的分配策略。
从表4可以看出,不同浓度硫酸钠处理下的分蘖数、株丛高、分株高和千粒质量差异不显著,硫酸钠处理下RA(Ⅰ)、分株高的相关性关系为:y=-0.036 2x+26.562,r2=0.005 3,二者之间呈负相关但相关性不显著。可见影响稗草生殖分株数量特征和生殖分配的是硅并不是其中的盐分。
3结论
(1)稗草进入生殖生长期后,一定范围内施硅有助于稗草生物量的积累,超过一定范围,硅对稗草生物量的积累起抑制作用。
(2)从整个生长周期来看,分蘖期和拔节期硅处理下的根和叶占有较大的生物量分配比例,为稗草后期的生殖生长积累了大量的有机物质。随着生长期的延长,稗草生殖器官生物量分配增大。结实期稗草通过调节有限的资源来保证子代的繁衍,在不施硅的条件下,稗草通过增加生殖器官的投入维持下一代的绝对数量,而Si3处理下,更多的资源分配到营养器官中,保证能量的吸收,维持生长,提高了稗草的竞争力。
(3)一定范围内施硅有助于提高稗草的形态和籽粒的表型可塑性。分蘖数和分株高随施硅量的增加呈先增加后下降的趋势。不同浓度硅处理下的株高、RAⅠ呈显著负相关,体现了不同硅浓度条件下稗草对资源的分配策略。
(4)通过对稗草生物量生殖分配和生殖分株数量特征研究发现,对稗草生长起作用的是硅,并不是其中的盐分。
参考文献:
[1]王仁忠,祖元刚,聂绍荃. 植物种群生殖生态学研究透视[J]. 东北师大学报:自然科学版,1999,10(2):22-23
[2]Murali K S,Sukumar R.Reproductive phenology of a tropical dry forest in Mudumalai,Southern India [J]. Journal of Ecology,1994,82:759-767.
[3]Smith R C,Armesto J.Flowering and fruiting pattern in the forest of chiloe,chile-ecologies and climatic constraints[J]. Journal of Ecology,1994,82:353-365.
[4]Jan Thiele.Flowering does not decrease vegetative competitiveness of Lolium perenne[J]. Basic and Applied Ecology,2009(10):340-348.
[5]Arahamson W G. Patterns of resources allocation in wildflower populations of field and woods[J]. American Journal of Botany,1979,66:71-79.
[6]操国兴,钟章成,谢德体,等. 不同群落中川鄂连蕊茶的生殖分配与个体大小之间关系的探讨[J]. 植物生态学报,2005,29(3):361-366.
[7]陈劲松,苏智先,缙云山. 马尾松种群生物量生殖配置研究[J]. 植物生态学报,2001,25(6):704-708.
[8]徐庆,刘世荣,臧润国,等. 中国特有植物四合木种群的生殖生态特征——种群生殖值及生殖分配研究[J]. 林业科学,2001,37(2):36-41.
[9]周小玲,马新娥,尚可为,等. 不同物候期胀果甘草生物量和营养物质生殖分配研究[J]. Acta Prataculturae Sinica,2012(4):25-32.
[10]包国章,康春莉,李向林.不同放牧强度对人工草地牧草生殖分配及种子重量的影响[J]. 生态学报,2002,22(8):1362-1366.
[11]郭力华,杨允菲,李建东. 松嫩平原不同生境寸草苔种群生殖分株的数量特征及生殖分配动态[J]. 草业学报,2005,14(2):63-68.
[12]张春华,杨允菲. 松嫩平原寸草苔种群生殖分株的种子生产与生殖分配策略[J]. 草业学报,2001,10(2):7-13.
[13]沈紫微,南志标. 甘南地区歪头菜生殖分配对生殖产量的影响[J]. 草业科学,2014,31(5):884-891.
[14]郭伟,潘星极,孙备,等. 不同氮素条件下稗草生物量生殖分配及生殖分株数量特征[J]. 吉林农业大学学报,2011,33(5):478-484.
[15]敖凤志,郭伟,范其阳,等. 不同供氮水平下稗草营养元素(N)的生育期分配研究[J]. 江西农业大学学报,2014(3).
[16]李海波. 水稻和稗草共生土壤微生物生物量碳及酶活性的变化[J]. 应用生态学报,2008,19(10):2234-2238.
[17]姜敏. 中国北方水稻与无芒稗化感作用研究[J]. 沈阳农业大学学报,2007.08,38(41):478-482.
[18]吴声敢,王强,赵学平,等. 稻田稗草生物学特性及其综合防除[J]. 杂草科学,2006(3):1-5.
[19]王宇,黄春艳,郭玉莲,等. 不同密度稗对春玉米产量的影响[J]. 杂草科学,2014,32(1):52-53.
[20]宫海军,陈坤明,王锁民,等. 植物硅营养的研究进展[J]. 西北植物学报,2004,24(12):2385-2392.
[21]温广月,钱振官,李涛,等. 上海地区稻田无芒稗对丁草胺抗药性的测定[J]. 杂草科学,2015,33(3):34-36.
[22]邓接楼,王艾平,何长水,等. 硅肥对水稻生长发育及产量品质的影响[J]. 广东农业科学,2011,38(12):58-61.
[23]龚金龙,胡雅杰,龙厚元,等. 不同时期施硅对超级稻产量和硅素吸收、利用效率的影响[J]. 中国农业科学,2012,45(8):
1475-1488.
[24]李金花,潘浩文,王刚. 草地植物种群繁殖对策研究[J]. 西北植物学报,2004.24(2):352-355.
[25]邢福,郭继勋,王珂. 狼毒种群生殖构件数量特征与生殖配置研究[J]. 草业学报,2005,14(4):111-115.
[26]靳牡丹,邹春静,宋晴. 草本植物生殖分株的数量特征和生殖分配的时空动态研究进展[J]. 山西师范大学学报:自然科学版,2007,21(1):80-85.
[27]郭正刚,田福平,王锁民,等. 硅对紫花苜蓿生物学特性的影响[J]. 生态学报,2006,26(10):3302-3307.
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
今日农业(2020年19期)2020-12-14
农家科技中旬版(2020年7期)2020-07-24
甘肃科技(2020年21期)2020-04-13
中国野生植物资源(2019年2期)2019-06-11
湖南林业科技(2017年1期)2017-02-06
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
