
横断山区干旱河谷川滇蔷薇生长与繁殖特征的地理空间差异
2016-07-21 01:07周志琼包维楷
生态学报 2016年8期
周志琼,包维楷
中国科学院成都生物研究所,中国科学院山地生态恢复与生物资源利用重点实验室,生态恢复与生物多样性保育四川省重点实验室, 成都 610041
横断山区干旱河谷川滇蔷薇生长与繁殖特征的地理空间差异
周志琼,包维楷*
中国科学院成都生物研究所,中国科学院山地生态恢复与生物资源利用重点实验室,生态恢复与生物多样性保育四川省重点实验室, 成都610041
摘要:通过大面积的野外踏查、种群调查和室内实验,分析了横断山区干旱河谷乡土灌木川滇蔷薇在岷江流域、大渡河流域、雅砻江流域和金沙江流域的分布、生长繁殖特征及其在流域间的差异。结果表明,川滇蔷薇广泛分布于四大流域,海拔范围1700 m至3700 m,所调查的18个种群生长状况良好,株高2 m以上,明显高于群落中的其他灌木。主成分分析表明,地理位置(流域、纬度、经度、海拔)和土壤(全碳、总有机碳、全氮、pH值)是影响川滇蔷薇生长繁殖的主要因子,其中地理位置的影响更大。生长特征包括株高、地径和冠幅在18个种群间差异显著,但在不同的流域间差异不显著。丛萌生数在种群间差异显著,其差异性较其他生长特征弱,但丛萌生数在不同流域间差异显著:岷江流域显著大于雅砻江流域,金沙江流域显著大于大渡河流域。与生长和无性繁殖相比,有性繁殖特征表现出更强的空间差异性,果实重量、瘦果重量以及种子活力在居群间差异极显著,果实重量和种子活力在不同流域间差异显著,但瘦果重量在流域间的差异不显著。综合分析表明,川滇蔷薇生长和繁殖特征在横断山区干旱河谷存在较强空间差异性,较强无性繁殖是种群持续存活的保证,而有性繁殖及其强烈的空间差异为种群适应多变环境以及地理空间扩展提供了遗传基础。
关键词:干旱河谷;地理位置;土壤特征;川滇蔷薇
干旱河谷是横断山区众多河流深切后形成的著名自然景观之一,主要分布在金沙江、怒江、岷江、大渡河等河流及其支流峡谷的下段[1]。受自然灾害与过度人为干扰的影响,干旱河谷局部地段植被严重退化,水土流失严重,地质灾害频繁。急剧恶化的生态环境不断削弱着区域社会经济发展成果,并直接威胁整个山地生态系统以及成都平原和中下游地区的生态安全[2]。
促进植被恢复,提高植被覆盖率,改善脆弱生态系统是该区生态建设的一项迫切任务[3]。 恶劣的土壤环境条件( 水分不足和养分贫乏)和气候条件是该区植被恢复的关键障碍,植被恢复与重建应选择耐旱、耐土壤瘠薄和盐碱性强的灌木和草种[3-5]。而干旱河谷的乡土灌木和草本植物是优先考虑的植被恢复物种[2]。目前,干旱河谷植物生态学的研究主要集中在生物量[6-7]、种间关系和群落特征[8-14]、植被与环境因子关系[15-17],优势灌木对干旱适应的生理生态机制[18-20]等方面。乡土植物生长与繁殖特性及其空间差异性研究较为缺乏,仅有的少数研究集中在特定流域内[3-4],缺乏对不同流域间乡土植物生长繁殖特征比较。研究干旱河谷的优势灌木和草本在不同流域的生长繁殖特征及差异,是干旱河谷植被恢复过程中适宜物种及优良种源筛选的关键。
川滇蔷薇(Rosasoulieana)是横断山区干旱河谷广泛分布乡土灌木,在部分地段已成为优势种,具有较明显的耐干旱和耐贫瘠的生态适应特点[2,21]。同时,它也是一种重要的资源植物,其植株含有能治疗心血管疾病的化学成分[22];花中含有酚类抗氧化剂[23],果实含有多种对人体有益的营养成分,具有较高的药用价值和经济价值。因此,研究川滇蔷薇生长与繁殖特性的地理空间差异可为蔷薇野生资源保护、优良种质资源的筛选提供依据。
川滇蔷薇为直立灌木,高2—4 m,萌生株数量可达20 株/丛,灌丛体积大;伞房花序,花瓣黄白色,花期5—7月,果期8—10月,成熟果实浅黄色至橘黄色,成熟后不立即脱落,一般能在枝头保持2—3个月;大量花与果实为昆虫和食果动物提供了丰富的食物来源[2,21]。本研究通过对横断山区干旱河谷川滇蔷薇分布、生长繁殖特征、地理要素及土壤因子调查,试图揭示川滇蔷薇在横断山区干旱河谷的分布现状,阐明影响生长繁殖的主要环境因素,揭示不同流域间川滇蔷薇生长繁殖特征的差异,为干旱河谷植被恢复过程的苗木母体种群选择,野生蔷薇资源利用与开发提供科学依据。
1研究区域
研究区域为横断山区干旱河谷的4个主要流域:岷江流域、大渡河流域、雅砻江流域以及金沙江流域(28°51′37.18″—32°15′55.56″N, 99°24′36.95″—103°44′47.52″E)河谷两侧山地,海拔1200—3700 m。该区域年均气温6—16℃,降水量308—720 mm,气候干燥[1]。植被为典型干旱河谷灌草丛,在不同地段植被覆盖率差异较大。大部分地段覆盖率为20%—40%之间,部分地段达60%以上。所有植物种类基本上为阳生,大多数种类具有叶片角质层发达、多刺、多毛的特点。土壤类型以山地褐土和山地棕壤为主。调查范围,岷江流域主河道:汶川绵虒至松潘镇江关,支流黑水河及杂谷脑河,约360 km,海拔1200—2460 m;大渡河流域:马尔康白湾乡至金川德威乡,约400 km,海拔1780—2260 m;雅砻江流域:雅江县马扎乡至米龙乡,约70 km,海拔2560—3060 m;金沙江流域:乡城县沙贡乡至德荣县松麦镇,约300 km,海拔2700—3700 m。
2研究方法
2.1样地设置与数据收集
2011年6月至7月,在川滇蔷薇的盛花期开展环境因子及生长状况调查。在全面踏查的基础上,针对不同流域不同海拔的典型分布区域进行样点布设,共布设18个样点,各样点的布局如图1。记录样点的8个地理位置参数,包括流域、经度、纬度、海拔、坡度、坡向、坡位和距离河谷的高度(表 1),然后选取10丛具有代表性的川滇蔷薇灌丛,测定灌丛高度、冠幅、萌生数、萌生株地径。2011年10月1日至20日,在川滇蔷薇果实及种子成熟期,开展川滇蔷薇繁殖性状调查。每个样点在10丛蔷薇(已调查生长特征)中选取5丛,在冠丛顶端的3个方向各选取一个当年生小枝,果实重量和种子重量的测定以小枝作为重复,测定每个小枝上果实数量及总重量,果实重量=果实总重量/果实数量。然后,将种子从果实中分离出来,测定种子数量与总重量,种子重量=种子总重量/种子数量。每丛蔷薇随机选取20粒种子,TTC染色法测定种子活力。
2.2土壤分析
在进行繁殖性状调查的灌丛下采集土壤样品,采集0—10 cm 土层土样。将取得的土壤样品进行常规理化性质分析。土壤样品测定的理化性质指标[24]包括:石砾含量(直径>1mm),pH 值(5∶1) 、全碳(%)、土壤有机碳(%)、全氮(%) 、全磷(mg/kg)。
2.3数据统计分析
坡向的量化方法是:以北为0,顺时针和逆时针两个方向都逐渐增大,值在0—180之间[3]。采用混合线性模型分析川滇蔷薇生长与繁殖参数在种群和流域之间的差异。分析种群间差异时,将种群作为固定因素,种群所在流域作为随机因素;分析流域间差异时,将流域作为固定因素,不同种群为随机因素嵌套入流域内。用Person相关系数分析各参数之间的相关关系。计算过程均在SPSS 21.0以及EXCEL 2010下完成。
主成分分析采用18个样地内的14个环境因子进行分析,环境因子分别为: 流域、经度、纬度、海拔、坡度、坡向、坡位、距离河谷的距离、pH值、全碳、有机碳、全氮、全磷、石砾含量。主成分分析可以根据川滇蔷薇各种群生长繁殖的差异,并与环境对生长繁殖参数的影响相结合,去除多余信息,简化数据。主成分分析中,首先对种群与环境因子构成的原始矩阵X(18,14)进行标准化,再分别计算出各因子的负荷量、特征根和贡献率,最后完成排序。主成分分析及相关(Person)分析过程在SPSS 11. 0下完成。
3结果与分析
3.1川滇蔷薇在干旱河谷的分布状况
川滇蔷薇在岷江、大渡河、雅砻江以及金沙江流域广泛分布,其主要分布区为各流域的典型干旱河谷地段以及干旱河谷过渡区两侧的山地灌丛。在岷江流域主要分布在茂县县城以北至松潘镇江关以南的主河道两侧,以及支流黑水河(黑水至两河口)和杂谷脑河(理县至汶川)两侧的山地灌丛中,河谷长度约为360 km;在大渡河流域主要分布在马尔康以南,丹巴县孔玉乡以北的主河道,以及支流小金河(小金至丹巴)与东谷河(八美至丹巴)两侧的山地中,河谷总长度约320 km;在雅砻江流域主要分布在雅江县城南北两个方向的河道两侧,以及高尔士山至雅江县城的河谷两侧,河谷长度约130 km;在金沙江流域分布于硕曲河、定曲河以及金沙江河谷两侧山地,以及附近山地中,总河谷长度约260 km,分布纬度范围为北纬28°51′37.18″至32°15′55.56″,东经99°24′36.95至103°44′47.52″,海拔范围1700 m至3700 m (图1,表 1)。

图1 横断山区干旱河谷川滇蔷薇分布示意图Fig.1 Distribution of Rosa soulieana in the dry valleys of the Hengduan mountain region

样号Plot地名Site流域Drainagebasin经度Longitude纬度Latitude海拔/mAltitude坡度/(°)Slopegradient坡向Slopeaspect距离河谷高度/mHeightfromriverP1镇江关岷江 103°44'47.52″32°15'55.56″24575SW20°40P2红岩 岷江 103°05'15.48″32°04'31.32″223710EN20°30P3较场 岷江 103°40'21.42″32°00'12.00″202318SE14°20P4飞虹 岷江 103°44'32.64″30°47'48.98″172515WS24°70P5渡口 大渡河101°57'41.16″31°46'13.20″225518EN24°100P6杨家湾大渡河102°03'59.34″31°25'45.42″214530NE5°60P7巴底 大渡河101°53'22.56″31°08'55.20″205510ES38°10P8甲居 大渡河101°52'48.36″30°57'18.48″201025NE16°80P9小金河大渡河101°59'45.24″30°57'13.8″201435WN45°100P10孔玉 大渡河102°02'54.48″30°42'9.42″17822EN30°2P11雅江北雅砻江101°02'00.19″29°56'45.62″26575SE22°10P12八角楼雅砻江101°13'48.09″30°02'30.07″30625WS13°20P13雅江南雅砻江101°02'00.16″29°56'45.60″256325NW15°10P14马熊沟金沙江99°56'01.11″29°09'09.10″36205SW5°600P15雨洼 金沙江99°53'34.22″29°04'38.75″29403NW3°15P16青德 金沙江99°49'54.24″28°51'37.18″270510NW14°30P17正斗 金沙江99°31'58.93″29°11'17.77″296630SW20°25P18茨巫 金沙江99°24'36.95″29°04'13.76″32355ES28°500
3.2影响川滇蔷薇生长繁殖的环境因子

表2 14个环境因子的主成分分析
14个环境因子可以简化为2个主成分,其贡献率分别为43.04%和17.19%,累积贡献率为60.23%,这2个轴可较好地反映出种群生长、繁殖和环境因子的差异。累计贡献率较低,主要因为流域、经度、纬度和海拔之间具有较高的相关性,但这是本研究中特殊地理环境特征引起,在分析时不能去除某一因子。第1轴负荷量较大的因子分别是流域(-0.953)、纬度(0.942)、经度(0.930)、海拔(- 0.840)、全碳(0.721)、总有机碳(0.687)、全氮(0.673);第2轴负荷较大的因子分别是pH值(-0.693)、全氮(0.673)、总有机碳(0.651)、全碳(0.645)(表 2)。8个因子可归为两类:地理因子(流域、纬度、经度和海拔)和土壤因子(全碳、总有机碳、全氮、pH值),其中地理因子影响更大。
环境因子间的相关分析表明,对川滇蔷薇生长和繁殖影响最大因素是流域和海拔,这两个因素除与坡位无显著相关外,与其他各环境因子都具有显著相关(表 3)。流域的变化造成了经纬度的变化、并与海拔共同影响土壤性质。
3.3川滇蔷薇生长特征的地理空间差异
所调查的4个流域中,川滇蔷薇18个种群生长状况良好,株高2m以上,明显高于群落中的其他灌木。对18个种群生长特征的分析表明,川滇蔷薇生长特征表现出显著的空间差异(图2),其中,株高、地径和冠幅的空间差异为极显著(P<0.01);萌生数为显著差异(0.01
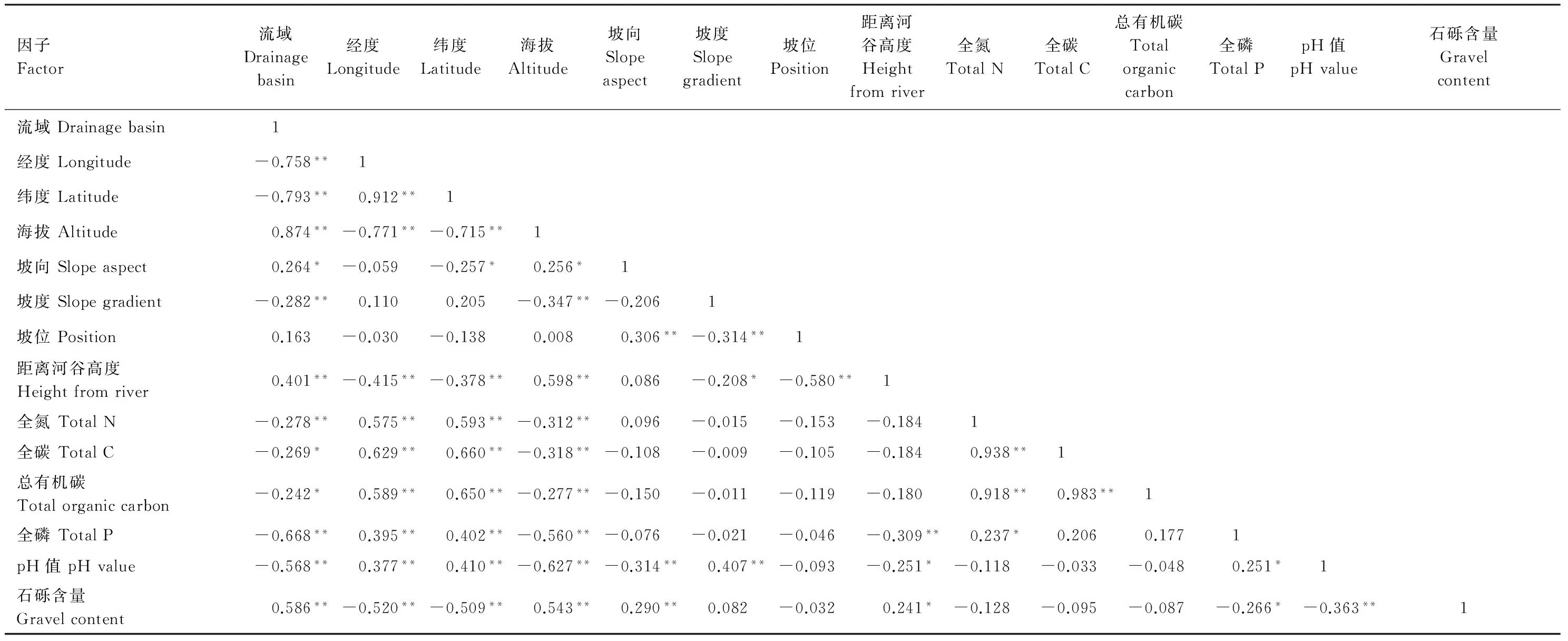
表3 川滇蔷薇18个种群的14个环境因子的相关性

图2 川滇蔷薇在横断山区4个干旱河谷流域中的生长特征Fig.2 Growth characteristics of Rosa soulieana in the four dry valleys of Hengduan mountain region (mean±S.E)岷江流域P1—P4,大渡河流域P5—P10,雅砻江河流域P11—P13,金沙江流域P14—P18
3.4川滇蔷薇繁殖特征的地理空间差异
与生长特征相比,川滇蔷薇的繁殖参数表现出更强的空间差异性。果实重量、瘦果重量和种子活力在18个居群间表现出极显著差异,其中果实重量差异最小,而瘦果重量差异最大(表 4,图 3)。不同流域间果实重量差异极显著(F=14.45, df=3,P<0.001),表现为:果实重量在雅砻江流域显著大于其他三流域;种子活力也有显著差异(F=5.43, df=3, 0.05
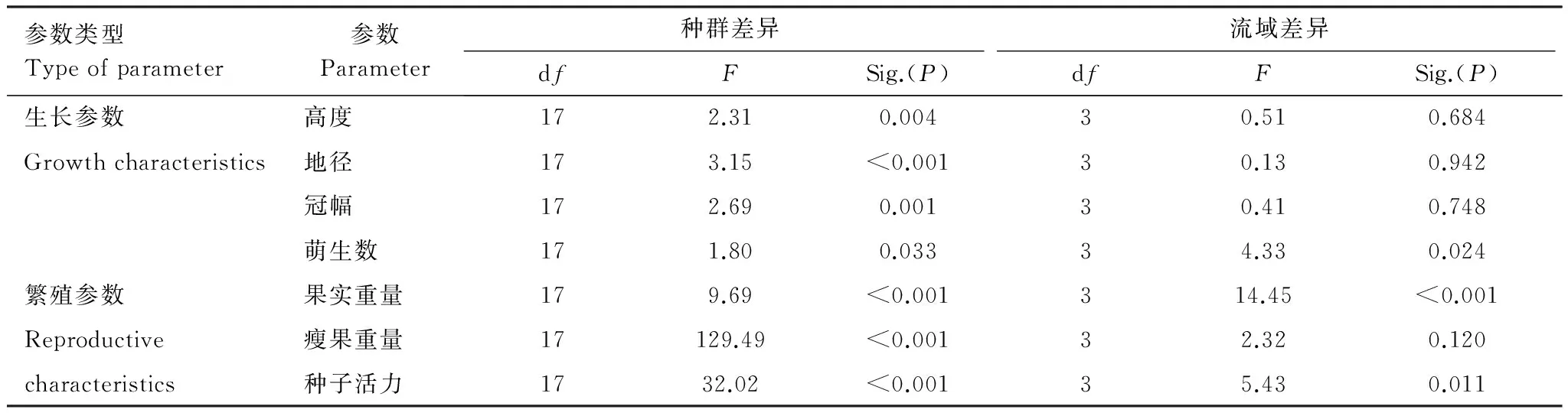
表4 川滇蔷薇18个种群各生长特征的混合线性模型分析

图3 川滇蔷薇在横断山区四大干旱河谷流域中的繁殖特征Fig.3 Reproductive characteristics of Rosa soulieana in dry valleys of Hengduan mountain region (mean±S.E)岷江流域P1—P4,大渡河流域P5—P10,雅砻江河流域P11—P13,金沙江流域P14—P18
4讨论
4.1横断山区干旱河谷川滇蔷薇生长与繁殖的基本特征
本研究表明川滇蔷薇在横断山区干旱河谷分布范围广,在所调查的4个流域的典型干旱区以及过渡区皆有分布,且生长状况良好,表明其适应干旱河谷缺水和高度异质的环境条件。另外,干旱河谷植被类型为低矮的灌草丛,植物高度一般在2m以下,而调查范围内川滇蔷薇平均高度为(2.64±0.17) m。因此,川滇蔷薇灌丛高大,为干旱河谷幼苗更新和存活提供适宜的微生境,利于植被覆盖率的保持与提高。
繁殖能力是衡量植物种群延续和扩展能力的重要指标,其中无性繁殖与有性繁殖是植物应对不同生存环境的重要策略[25]。研究发现,川滇蔷薇具有较强的无性萌生能力,灌丛平均萌生数为(12.5±1.9)株/丛,最多可达25株/丛,远高于干旱河谷同属植物多苞蔷薇(5.07±0.39)株/丛和黄蔷薇(6.48±0.97)株/丛[3-4]。灌丛中10%—30%为一至两年生的萌生株,表明川滇蔷薇当前萌生更新能力强。无性萌生具有高成活率特点,是植物在干旱恶劣环境中保持种群稳定的重要方式。在有性繁殖方面,由于本研究持续时间较长,在调查期间已有部分果实脱落,因此没有进行结实量调查。据前期调查表明,川滇蔷薇结实量巨大,岷江干旱河谷结实量为(9822±2129)粒/丛[2]。本研究中,川滇蔷薇果实中种子数量大,单果种子数量为(18.5±4.8)粒;种子活力高,大部分种群的种子活力大于80% (图3)。总体而言,横断山区干旱河谷川滇蔷薇活力种子产量高,有性繁殖能力强。此外,与干旱河谷广泛分布的黄蔷薇(深度休眠)和多苞蔷薇(中度休眠)相比,川滇蔷薇种子休眠程度较低,为非深度休眠类型;种子库中活力种子数量相对更多,存在持久种子库[5]。Grime系统地将植物繁殖对策归纳为:营养扩展、植被空隙中季节性更新、持久种子(或传播体)库更新、大量风媒种子(或传播体)更新和持久幼苗库更新。据此,川滇蔷薇的繁殖对策有两种:营养扩展和持久种子库更新。川滇蔷薇较强无性萌生能力,保证其在干旱贫瘠的环境保持种群持续存活,而较强的有性繁殖能力为保持种群遗传多样性,应对多变的干旱河谷环境提供遗传物质基础,同时也是种群在空间上不断扩展的重要方式。
4.2横断山区干旱河谷川滇蔷薇种群生长繁殖的空间格局
主成分和相关分析结果表明地理位置和土壤共同作用影响川滇蔷薇种群生长与繁殖。地理因素中流域、经度、纬度和海拔影响最大,表明川滇蔷薇不同种群地理环境差异是导致其生长繁殖空间差异的最主要因素;而土壤因子对其生长繁殖差异性贡献相对较小。地理因素控制多种环境因子,如气象、土壤和植被状况等,因此是影响植物生长繁殖及相关功能性状的最重要因素[26],直接控制植物的养分与水分供给状况,因此也对植物功能性状有着强烈影响,但土壤在小空间尺度范围内变异较大,对种群生长繁殖的影响力相对较弱。
横断山区干旱河谷区受高山峡谷地形和大陆性季风气候的影响,生态因子特别是水热条件空间变化表现为:从南至北气温降低,湿度和降水量增加的趋势;在同一流域内表现为随海拔高度升高气温降低,湿度和降水量增加,而蒸发量减小,辐射增强,这是干旱河谷生态环境因子空间变化的基本特点[14]。川滇蔷薇生长特征(高度、地径和冠幅)在18个种群间存在显著差异,表现出较强的形态可塑性,而表型可塑性是植物适应异质环境的重要策略。但本研究中,川滇蔷薇的生长特征在不同流域间没有显著差异,表明该灌木的生长特征主要受所在位置的海拔、纬度、气候及土壤因子等差异的影响,而各流域的大气候条件影响相对较弱。一般情况下高海拔种群比低海拔种群表现出更低的株高[27],本研究中,只有岷江流域川滇蔷薇株高表现出类似的海拔格局,但其他3个流域中株高没有显著差异或明显海拔变化趋势,这可能是由于干旱河谷主要的环境限制因素为水分,海拔的影响相对较弱。
繁殖特征在种群间以及流域间都表现出显著差异,表明与生长特征相比,繁殖特征的空间差异性更强。种子大小是植物最重要生活史特征之一,影响种子散布、幼苗定居以及种群的分布格局,种子大小随环境的变化而改变是植物适应异质环境的重要策略[28]。本研究中,与果实重量和种子活力相比,瘦果(种子)重量在种群间的差异性更为显著,证明了种子大小是植物适应环境的重要方式。川滇蔷薇瘦果重量与海拔呈显著负相关关系,即种子大小随海拔升高而呈降低的趋势,该结果与岷江干旱河谷同属植物多苞蔷薇种子大小随海拔(1700—2500 m)梯度变化的格局相反[29];但与瑞士东南部的多年生草本植物Eritrichiumnanum种子大小在海拔梯度(2435—3055 m)[30]上的变化趋势一致。因此,不同地区不同物种种子大小地理空间差异性存在很大不确定性,受多种环境因素如气候、土壤,同时也受到植物自身特征(如植株大小)的影响。
致谢:衷心感谢成都生物所李甫、杨春燕、李金容、何大海博士等在野外调查中的大力协助,感谢成都生物所茂县生态站在野外调查中提供方便。
参考文献(References):
[1]张荣祖. 横断山区干旱河谷. 北京: 科学出版社, 1992.
[2]包维楷, 庞学勇, 李芳兰, 周志琼. 干旱河谷生态恢复与持续管理的科学基础. 北京: 科学出版社, 2012.
[3]周志琼, 包维楷, 吴福忠, 庞学勇, 何晓, 吴宁. 岷江干旱河谷多苞蔷薇生长与繁殖特征. 应用生态学报, 2007, 18(7): 1407- 1413.
[4]周志琼, 包维楷, 吴福忠, 何晓, 吴宁. 岷江干旱河谷黄蔷薇生长与繁殖特征及其空间差异. 生态学报, 2008, 28(4): 1820- 1828.
[5]周志琼, 包维楷, 吴福忠, 吴宁. 岷江干旱河谷黄蔷薇和川滇蔷薇更新能力及其限制因素. 生态学报, 2009, 29(4): 1931- 1939.
[6]刘国华, 马克明, 傅伯杰, 关文彬, 康永祥, 周建云, 刘世梁. 岷江干旱河谷主要灌丛类型地上生物量研究. 生态学报, 2003, 23(9): 1957- 1964.
[7]刘国华, 张洁瑜, 张育新, 周建云, 关文彬, 马克明, 傅伯杰. 岷江干旱河谷三种主要灌丛地上生物量的分布规律. 山地学报, 2003, 21(1): 24- 32.
[8]关文彬, 冶民生, 马克明, 刘国华, 汪西林, 谭辉. 岷江干旱河谷植物群落物种周转速率与环境因子的关系. 生态学报, 2004, 24(11): 2367- 2373.
[9]关文彬, 冶民生, 马克明, 刘国华, 汪西林. 岷江干旱河谷植被分类及其主要类型. 山地学报, 2004, 22(6): 679- 686.
[10]冶民生, 关文彬, 白占雄, 马克明, 刘国华, 汪西林. 岷江干旱河谷植物群落生态梯度分析. 中国水土保持科学, 2005, 3(2): 70- 75.
[11]刘文彬. 岷江上游半干旱河谷的主要类型. 山地研究, 1994, 12(1): 27- 31.
[12]冶民生, 关文彬, 谭辉, 马克明, 刘国华, 汪西林. 岷江干旱河谷灌丛α多样性分析. 生态学报, 2004, 24(6): 1123- 1130.
[13]李丽光, 何兴元, 李秀珍, 问青春, 赵永华, 胡志斌, 常禹, 朱亚平. 岷江上游干旱河谷农林边界影响域的研究. 应用生态学报, 2004, 15(10): 1804- 1808.
[14]刘庆, 包维楷, 乔永康, 钱能斌. 岷江上游茂县半干旱河谷灌丛优势种间关系的研究. 应用与环境生物学报, 1996, 2(1): 36- 42.
[15]张文辉, 卢涛, 马克明, 周建云, 刘世梁. 岷江上游干旱河谷植物群落分布的环境与空间因素分析. 生态学报, 2004, 24(3): 552- 559.
[16]刘世梁, 傅伯杰, 马克明, 刘国华. 岷江上游高原植被类型与景观特征对土壤性质的影响. 应用生态学报, 2004, 15(1): 26- 30.
[17]何其华, 何永华, 包维楷. 岷江上游干旱河谷典型阳坡海拔梯度上土壤水分动态. 应用与环境生物学报, 2004, 10(1): 68- 74.
[18]Li F L, Bao W K, Wu N. Morphological, anatomical and physiological responses ofCampylotropispolyantha(Franch.)Schindl. seedlings to progressive water stress. Scientia Horticulturae, 2011, 127(3): 436- 443.
[19]Gong M G, Tang M, Chen H, Zhang Q M, Feng X X. Effects of two Glomus species on the growth and physiological performance ofSophoradavidiiseedlings under water stress. New Forests, 2013, 44(3): 399- 408.
[20]Wu F Z, Bao W K, Li F L, Wu N. Effects of drought stress and N supply on the growth, biomass partitioning and water-use efficiency ofSophoradavidiiseedlings. Environmental and Experimental Botany, 2008, 63(1/3): 248- 255.
[21]中国科学院中国植物志编辑委员会. 中国植物志 第三十七卷. 北京: 科学出版社, 1985: 442- 442.
[22]陈封政, 赵卫权, 何永华, 丁立生, 王明奎. 川滇蔷薇和多苞蔷薇化学成分的研究. 应用与环境生物学报, 2000, 6(4): 334- 336.
[23]Yang C Y, Li F, Zhang X L, Wang L, Zhou Z Q, Wang M K. Phenolic antioxidants fromRosasoulieanaflowers. Natural Product Research, 2013, 27(21): 2055- 2058.
[24]中国科学院南京土壤研究所. 土壤理化分析. 上海: 上海科学技术出版社, 1978.
[25]刘志民, 蒋德明, 高红瑛. 植物生活史繁殖对策与干扰关系的研究. 应用生态学报, 2003,14(3): 418-422.
[26]Zhou W , Wang Z X, Davy A J, Liu G H. Geographic variation and local adaptation inOryzarufipogonacross its climatic range in China. Journal of Ecology, 2013, 101(6): 1498- 1508.
[27]Premoli A C, Brewer C A. Environmental v. genetically driven variation in ecophysiological traits ofNothofaguspumiliofrom contrasting elevations. Australian Journal of Botany, 2007, 55(6): 585- 591.
[28]Wang Y J, Wang J J, Lai L M, Jiang L H, Zhuang P, Zhang L H, Zheng Y R, Baskin J M, Bakin C C. Geographic variation in seed traits within and among forty-two species ofRhododendron(Ericaceae) on the Tibetan plateau: relationships with altitude, habitat, plant height, and phylogeny. Ecology and Evolution, 2014, 4(10): 1913- 1923.
[29]Zhou Z Q, Bao W K. Changes in seed dormancy ofRosamultibracteataHemsl. & E. H. Wilson with increasing elevation in an arid valley in the eastern Tibetan Plateau. Ecological Research, 2014, 29: 693- 700.
[30]Wirth L R, Graf R, Gugerli F, Landergott U, Holderegger R. Between-year variation in seed weights across altitudes in the high-alpine plantEritrichiumnanum. Plant Ecology, 2010, 207(2): 227- 231.
Geographic variation in growth and reproduction ofRosasoulieanain the dry valleys of the Hengduan Mountain region
ZHOU Zhiqiong, BAO Weikai*
KeyLaboratoryofMountainEcologicalRestorationandBioresourceUtilization&EcologicalRestorationBiodiversityConservationKeyLaboratoryofSichuanProvince,ChengduInstituteofBiology,ChineseAcademyofSciences.,Chengdu610041,China
Abstract:Understanding the geographic variation of the growth and reproduction characteristics of native plants may help to develop effective strategies for vegetation restoration in degraded regions. Rosa soulieana, a native shrub widely distributing in the dry valleys of the Hengduan Mountain region, appears to have a great potential to rehabilitate the vegetation because of its ecological and economic advantages. To determine the distribution, growth and reproduction of this species, 18 plots were investigated along the dry valleys of four rivers (Minjiang, Daduhe, Yalongjiang and Jinshajiang) in June and October 2011. Eight location parameters of each plot were recorded including drainage, latitude, longitude, altitude, slope gradient, slope aspect, position, and height from river. Ten clumps were randomly selected for measuring the growth parameter, and five clumps for reproductive parameters in each plot. Soil parameters were measured using samples collected from the five clumps that had been used to record reproductive parameters. The results indicated that R. soulieana was widely distributed in each dry valley of Hengduan Mountain region at altitudes from 1,700 to 3,700 m above sea level. Mean plant height was (2.64 ± 0.17) m, which was higher than that of other species in the community in the dry valley region. PCA and correlation analyses showed that location (drainage, latitude, longitude, and altitude) and soil traits (total carbon, total organic carbon, total nitrogen, and pH) were the principal factors influencing growth and reproduction, and location had a stronger influence than that of soil traits. Growth traits (height, base diameter, and crown diameter) differed significantly among the 18 populations, but not among drainages. Vegetative reproduction was high with a mean of (2.5 ± 1.9) ramets per individual, with a maximum of 25 ramets on a single individual. Ramet number per individual differed less among the populations than growth traits, but it was significantly different among drainages. R. soulieana also exhibited strong sexual reproduction. Seed vitality of 83.3% of the populations was >80%. The reproductive parameters exhibited stronger spatial variation than growth traits. Fruit mass, achene mass, and seed vitality were highly significantly different among the 18 populations, but only fruit mass and seed vitality were significantly different among the four drainages. Fruit mass was significantly greater in the Yalongjiang drainage than in the other three. Seed vitality was greater in Minjiang and Jinshajiang drainages than in the Yalongjiang and Daduhe drainages. Comprehensive analysis showed pronounced geographic variation in growth and reproduction in R. soulieana in the dry valleys of the Hengduan Mountain region. Strong vegetative reproduction ensures the persistence of the populations, and sexual reproduction provides genetic material for this species to adapt to the environment and expand their distribution in dry valleys.
Key Words:dry valley; location; Rosa soulieana; soil characteristics
基金项目:国家自然科学基金项目(31200297);植被与环境变化国家重点实验室开发课题(LVEC-2012KF04)
收稿日期:2014- 10- 27;
修订日期:2015- 12- 10
*通讯作者
Corresponding author.E-mail: baowk@cib.ac.cn
DOI:10.5846/stxb201410272102
周志琼,包维楷.横断山区干旱河谷川滇蔷薇生长与繁殖特征的地理空间差异.生态学报,2016,36(8):2192- 2201.
Zhou Z Q, Bao W K.Geographic variation in growth and reproduction ofRosasoulieanain the dry valleys of the Hengduan Mountain region.Acta Ecologica Sinica,2016,36(8):2192- 2201.
猜你喜欢
散文诗世界(2021年12期)2021-12-17
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
经济研究导刊(2019年22期)2019-09-17
Zoological Research(2019年3期)2019-06-28
岷峨诗稿(2019年4期)2019-04-20
阿来研究(2018年1期)2018-07-10
乡村地理(2018年4期)2018-03-23
——唐天马长篇小说《横断山》简析
边疆文学(文艺评论)(2018年3期)2018-03-21
廉政瞭望(2016年13期)2016-08-11
