
水稻叶片早衰突变体ospls3的生理特征和基因定位
2016-07-15 05:45盼黎坤瑜黄福灯韦荔全杨茜程方民潘
作物学报 2016年5期
关键词:水稻
龚 盼黎坤瑜黄福灯韦荔全杨 茜程方民潘 刚,*
1浙江大学农业与生物技术学院, 浙江杭州310058;2浙江省农业科学院, 浙江杭州310021
水稻叶片早衰突变体ospls3的生理特征和基因定位
龚 盼1,**黎坤瑜1,**黄福灯2韦荔全1杨 茜1程方民1潘 刚1,*
1
浙江大学农业与生物技术学院, 浙江杭州310058;2浙江省农业科学院, 浙江杭州310021
摘 要:叶片衰老是作物叶片发育的最后阶段, 功能叶早衰将影响作物产量和品质, 因此, 研究叶片早衰的分子与生理机制对于培育耐早衰优良品种具有重要意义。本研究利用60Co-γ辐射诱变籼稻N142, 获得叶片早衰突变体ospls3,其叶片早衰始于分蘖期, 最先表现为叶尖变褐及叶中上部出现褐色斑点, 并向叶基部蔓延而使叶片枯死。生理分析表明, 野生型剑叶的叶绿素含量显著低于倒二叶和倒三叶, 而突变体的含量则分别低于野生型且依次显著降低; 野生型剑叶、倒二叶和倒三叶间的超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、丙二醛(MDA)含量、O2-含量和
H2O2含量基本不变, 而突变体的这些活性和含量则依次显著升高; 野生型剑叶、倒二叶和倒三叶的可溶性蛋白含量和过氧化氢酶(CAT)活性变化不显著, 而突变体则依次降低。遗传分析表明, ospls3突变性状受1对隐性基因控制。借助图位克隆技术, 将该基因定位于第12染色体长臂的RM6953与RM28753之间, 物理距离为294 kb, 该结果为进一步克隆OsPLS3基因并研究其功能奠定了基础。
关键词:水稻; ospls3; 叶片早衰; 生理分析; 基因定位
本研究由国家自然科学基金项目(31271691)和国家转基因生物新品种培育重大专项(2013ZX08001-002)资助。
This study was supported by the National Natural Science Foundation of China (31271691) and the Major Project of China on New Varieties of GMO Cultivation (2013ZX08001-002).
URL∶ http∶//www.cnki.net/kcms/detail/11.1809.S.20160119.1327.014.html
叶片衰老是植物生长发育的必经阶段, 也是植物适应环境的一种重要表现[1-2]。水稻等主要农作物在灌浆中后期功能叶早衰, 将导致其结实率显著降低、空秕率增加[3]、产量下降及品质性状(垩白和整精米率等)变劣等“负面”效应[4-5]。据报道, 水稻灌浆后期功能叶每推迟1 d衰老, 理论上可增产2%左右,实际能增产1%左右[6], 而且还能改善稻米品质[7]。
关于植物叶片衰老成因, 现已提出许多重要的学术观点或理论假说, 包括自由基损伤说、基因调控说、光碳失衡说、营养胁迫说和激素平衡说等[8]。尽管这些假说都在一定程度上解释了叶片的衰老启动及衰老进程, 但还未真正探明植物叶片衰老的机制。利用化学诱变及转基因T-DNA插入技术获得叶片早衰突变体是研究叶片早衰分子机制的重要手段之一。基于突变体策略, 至今已从水稻中克隆了14个叶片早衰基因, 包括UAP1[9]、OsNAP[10]、OsSIK2[11]、OsWRKY42[12]、OsABC1-2[13]、SGR[14-15]、OsAkαGal[16]、RLS1[17]、nol1[18]、nyc1[19]、NYC3[20]、NYC4[21]、SPL28[22]和OsLMS[23]。
本课题组利用60Co-γ辐射诱变中籼恢复系N142,获得一个隐性叶片早衰突变体, 暂命名为ospls3 (Oryza s ativa p recocious leaf s enescence 3)。该突变体叶片早衰表型在水稻分蘖期(约4~5片叶龄时)就开始显现, 之后随着叶龄的增加, 除心叶外, 其他叶片均不同程度早衰; 孕穗后期后则所有叶片均早衰。本文研究突变体ospls3的基本表型、生理变化及基因定位, 以期为进一步克隆该基因并揭示其早衰的分子和生理机制提供理论基础。
1 材料与方法
1.1 试验材料
叶片早衰突变体ospls3, 经杭州和海南连续8代回交和自交, 获得突变性状稳定株系。之后以ospls3为母本, 分别与N142和粳稻02428杂交获得F2群体并用于遗传分析。所有材料均种植于浙江大学农业试验站。水稻成熟后, 分别取 ospls3及其野生型对照N142各20株, 调查它们的株高、有效穗数、每穗粒数、结实率、千粒重和单株产量等主要农艺性状。
1.2 生理指标测定
选取ospls3及其野生型对照N142各12株处于孕穗期(剑叶叶枕与倒二叶叶枕平齐的分蘖)的剑叶、倒二叶和倒三叶分别测定叶绿素含量、可溶性蛋白含量、H2O2含量、超氧阴离子含量(O2-)、MDA含量、CAT活性、SOD活性和POD活性。用碧云天过氧化氢检测试剂盒(s0038)测定H2O2含量, 提取方法参见说明书。参考《植物生理学实验技术》测定其他生理指标[24], 每个指标测定3个重复。
1.3 遗传分析与基因定位
在大田分单株剪取 2014年 ospls3/02428的 F2定位群体中具有突变体性状的早衰单株、双亲以及F2群体中 10株正常植株的叶片, 采用 CTAB法[25]提取基因组总DNA。利用均匀分布于水稻12条染色体的500对SSR标记, 其引物序列来自Gramene数据库(http∶//www.gramene.org/), 以及Shen等[26]所开发的50对InDel标记进行基因定位。PCR总反应体系为20 μL, 含0.8 U Taq DNA聚合酶、1 × PCR buffer (Mg2+Plus)、1 mmol L-1dNTP Mixture、50 ng DNA、上下游引物各0.25 μmol L-1。PCR反应条件为95℃预变性5 min; 94℃, 30 s; 55℃, 30 s; 72℃, 30 s, 共35个循环; 72℃, 10 min。PCR产物经8%非变性聚丙烯酰胺凝胶电泳, 快速银染后观察[27]。
1.4 遗传图谱构建
从786株的ospls3/02428 F2群体中获得190个具有早衰性状的单株组成基因定位群体, 利用上述筛选到的基因连锁分子标记进行基因定位。同时在Gramene数据库查找基因连锁分子标记在染色体上的具体物理位置以确定其排列顺序, 结合每个标记的重组交换单株确定OsPLS3基因与连锁标记的顺序, 构建基因定位图谱。
2 结果与分析
2.1 突变体ospls3与野生型N142的表型及其主要农艺性状
突变体 ospls3的叶片在分蘖期就表现早衰性状(图1-A), 除心叶外, 各叶位叶片完整展开5~7 d后,从叶尖和叶中上部开始出现褐色斑点(图1-D), 并向叶基部迅速蔓延而使叶片枯死。孕穗后期所有叶片均不同程度早衰(图1-B, C)。叶片早衰导致突变体ospls3的主要农艺性状, 如株高、有效穗数、每穗粒数、结实率、千粒重和单株产量分别比野生型下降27.74%、32.03%、69.08%、71.35%、36.03%和95.12%(表1)。
2.2 突变体ospls3与野生型对照N142的生理性状
2.2.1 叶绿素含量 孕穗期突变体 ospls3的剑叶、倒二叶和倒三叶的叶绿素含量及叶绿素a/b的值依次显著下降(表2), 倒二叶和倒三叶的总叶绿素含量分别比剑叶下降7.02%和53.24%。与野生型相比,突变体ospls3的剑叶、倒二叶和倒三叶的总叶绿素含量分别下降22.78%、47.63%和74.66% (表2)。
2.2.2 SOD、POD和CAT的活性 图2表明突变体倒二叶和倒三叶的SOD和POD活性显著高于野生型,而CAT的活性则极显著低于野生型。相对于突变体剑叶, 倒二叶和倒三叶的SOD活性分别增加12.03% 和21.79%, POD活性分别升高100.00%和100.76%,而CAT活性则分别下降23.04%和47.76%。
2.2.3 超氧阴离子(O2-)含量和H2O2含量 图3-A 和B显示, 野生型剑叶、倒二叶和倒三叶间的O2-含量和H2O2含量无显著性差异, 与野生型相比, 突变体ospls3剑叶、倒二叶和倒三叶的O2-含量分别增加15.88%、49.08%和194.30%, 而H2O2含量则分别增加28.46%、22.79%和88.18%。
2.2.4 MDA和可溶性蛋白含量 图3-C显示, 野生型的剑叶、倒二叶和倒三叶间的MDA含量和可溶性蛋白含量无显著性差异, 与野生型相比, 突变体ospls3剑叶、倒二叶和倒三叶的MDA含量分别增加38.25%、39.96%和125.31% (图3-C); 而突变体除剑叶外, 其倒二叶和倒三叶的可溶性蛋白含量分别下降13.17%和31.83% (图3-D)。

图1 不同生育期突变体ospls3及其野生型的表型Fig.1 Phenotype of ospls3 and its wild-type (WT) plants at different growth stages
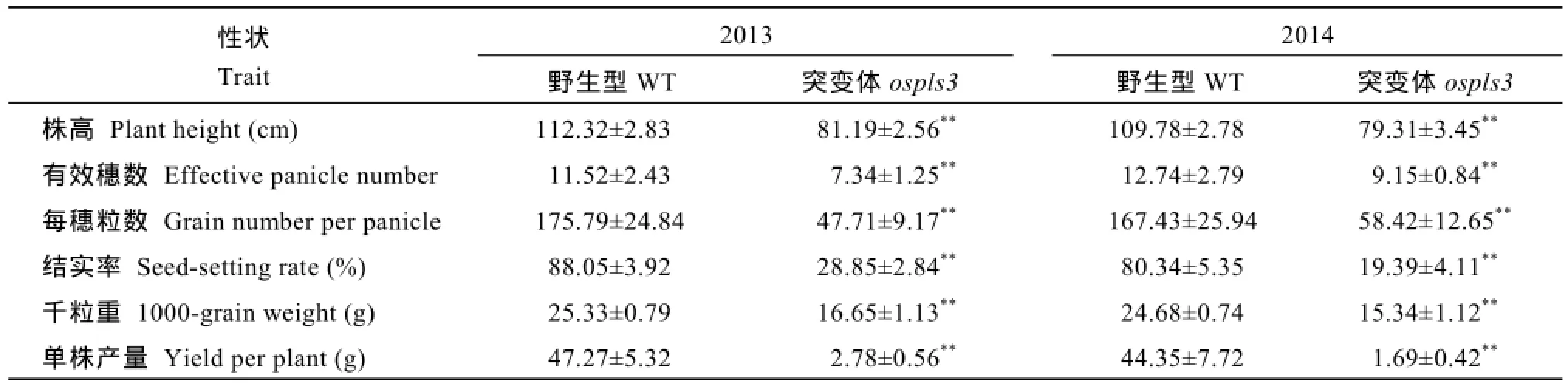
表1 突变体ospls3及其野生型的农艺性状Table 1 Agronomic traits of ospls3 and its wild-type (WT) plants

表2 孕穗期突变体ospls3及其野生型叶片的叶绿素含量Table 2 Chlorophyll content of leaves in ospls3 and its wild-type (WT) plants at booting stage

图2 孕穗期突变体ospls3及其野生型叶片的SOD、POD和CAT的活性Fig.2 SOD, POD, and CAT activities of ospls3 and its wild-type (WT) plants at booting stage
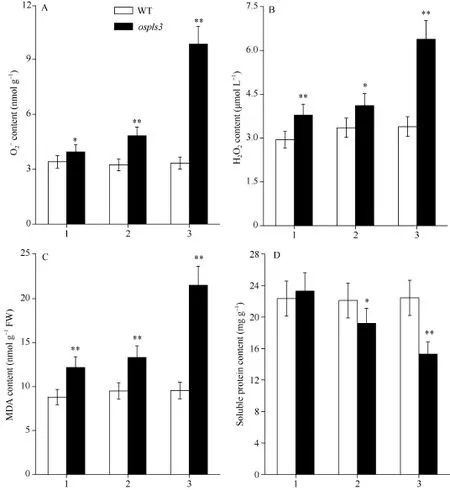
图3 孕穗期突变体ospls3及其野生型叶片的O2-、H2O2、MDA和可溶性蛋白含量Fig.3 O2-, H2O2, MDA, and soluble protein contents of ospls3 and its wild-type (WT) plants at booting stage
2.3 遗传分析与基因定位
ospls3/N142和ospls3/02428两个杂交F1单株的叶色正常, 说明突变体叶片早衰性状是由隐性位点控制。在 ospls3/N142的 638个 F2单株中, 有 153株表现叶片早衰, 485株表现正常, 正常单株∶早衰单株=3∶1 [χ2=0.30 < 3.84 (χ2(0.05, 1))]; 而在ospls3/02428的786个F2定位群体中, 正常单株为596株,早衰单株为 190株, 正常单株∶早衰单株=3∶1 [χ2=0.24 < 3.84 (χ2(0.05, 1))]。遗传结果均符合孟德尔单基因隐性遗传的分离比例, 从而证实突变体 ospls3的叶片早衰症状受单隐性核基因控制。
为了定位OsPLS3基因, 分别选取ospls/02428 F2群体中10株正常株和10株早衰突变株构建正常基因池和突变基因池, 选取均匀分布于水稻12条染色体上的SSR标记与InDel标记, 确定各标记与OsPLS3基因的连锁关系。结果发现水稻第12条染色体上的SSR标记RM28466、RM3739、RM1194及RM28826 与OsPLS3连锁。利用这4个标记对ospls3/02428的F2群体中的190个早衰单株进行基因型分析, 发现RM28466、RM3739、RM1194和RM28826的交换单株分别是50、19、16和21株。进一步在RM1194和RM28826之间设计了16个SSR标记, 利用其中10个在定位群体亲本间具多态性的标记将OsPLS3基因定位在RM6953和RM28753之间, 物理距离为294 kb, 横跨BX000457、AL731885和AL845347三个BAC (图4)。其间有EST支持的开放阅读框(ORF)为21个(http∶//rapdb.dna.affrc.go.jp/viewer/gbrowse/build5/) (表3)。
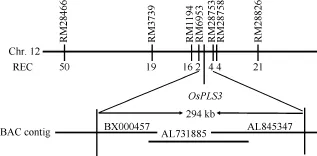
图4 OsPLS3基因在第12染色体长臂上的分子定位Fig.4 Molecular mapping of OsPLS3 gene on the long arm of chromosome 12
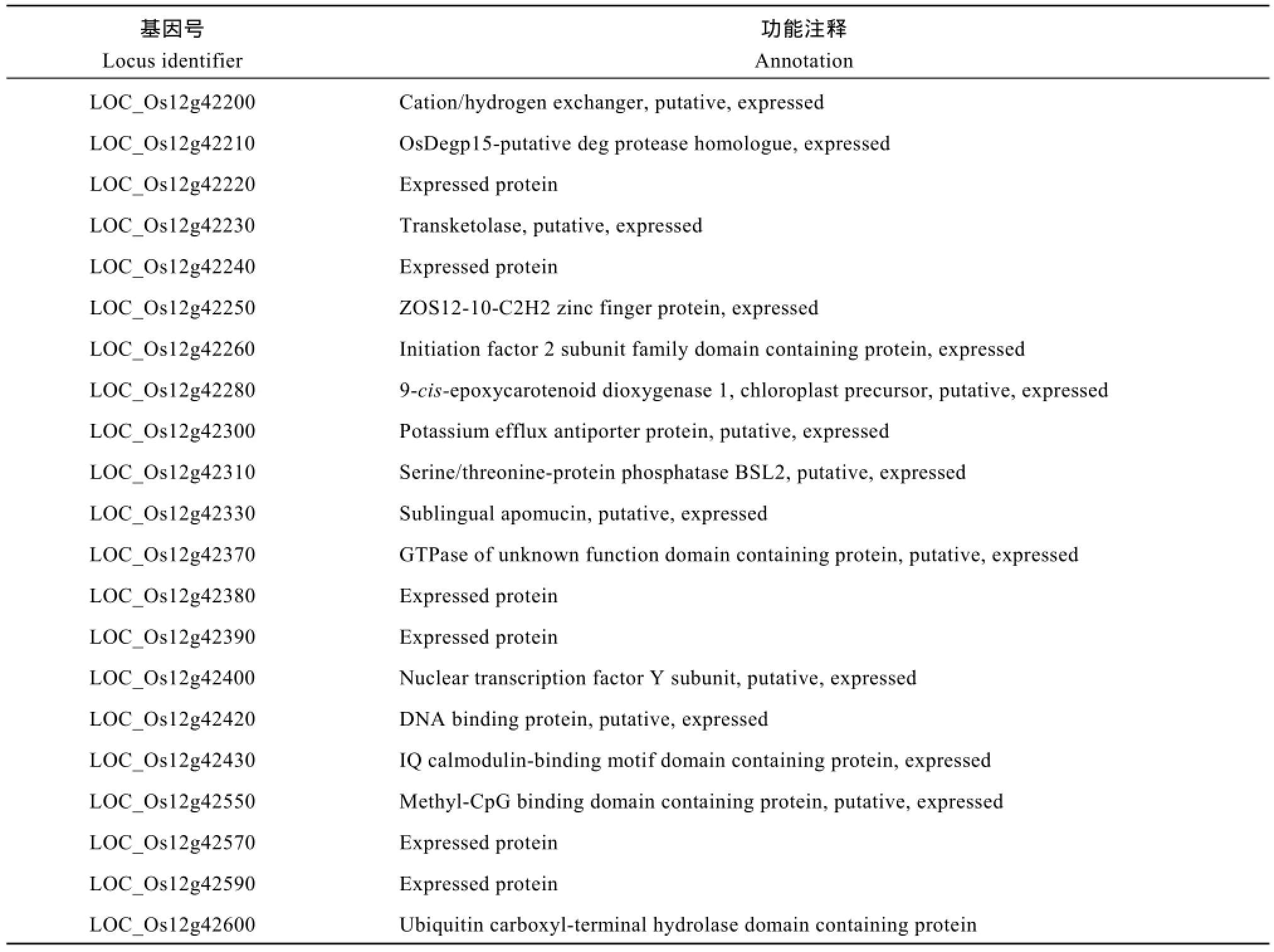
表3 定位区间内的基因及功能注释Table 3 Gene names and their annotations in the target interval
3 讨论
叶片衰老最明显的外在表现是叶片黄化, 而内在生理生化变化则包括叶绿体降解、蛋白水解酶活性增加、蛋白质降解、活性氧清除系统受到抑制、自由基含量及丙二醛(MDA)含量急剧增加等[1,28]。本研究利用辐射诱变获得叶片早衰突变体ospls3, 其叶片早衰症状始于分蘖期(图1-A), 孕穗后期则所有叶片均早衰(图1-B)。相对于野生型, 突变体ospls3的剑叶、倒二叶和倒三叶的总叶绿素含量分别下降22.78%、47.63%和74.66% (表2)。说明突变体的剑叶、倒二叶和倒三叶均已衰老, 且衰老程度依次加重。研究表明, 叶绿素a/b的值是衡量植物叶片光合效率的重要生理指标之一[29]。本研究中与突变体剑叶相比, 其倒二叶和倒三叶的叶绿素a含量下降率均比叶绿素b高(表2)。结果说明突变体叶片的光合效率将受到抑制, 且叶绿素a比叶绿素b下降更快, 究其原因, 可能是倒二叶和倒三叶中的活性氧离子含量显著高于剑叶(图3-A), 导致叶绿素a对活性氧离子的反应比叶绿素b更敏感[2,30]。
叶绿体是植物ROS (reactive oxygen species,ROS)形成的主要场所, 伴随衰老叶片叶绿体的解体以及其他逆境胁迫, 叶绿体中的电子传递链将受到抑制, 致使1O2、H2O2和O2-大量形成[31]。同时, 高浓度的ROS不仅作用于脂质, 使其发生过氧化反应而产生大量MDA, 加剧膜的损伤, 还通过细胞氧化应激反应诱导细胞凋亡甚至坏死[32]。本研究中野生型剑叶、倒二叶和倒三叶间的H2O2、O2-和MDA含量无显著差异, 而突变体的含量分别显著高于野生型并依次增加(图3)。此外, 蛋白质含量特别是可溶性蛋白质含量的下降是衡量叶片衰老的重要指标之一[1]。本研究中野生型倒二叶和倒三叶的可溶性蛋白含量极显著高于突变体, 说明突变体倒二叶和倒三叶明显衰老。作为可溶性蛋白的抗氧化系统酶SOD、POD 和CAT, 担负着清除细胞内ROS并保护膜系统的重任。野生型剑叶的CAT活性低于突变体(图-C), 其原因可能是野生型剑叶尚未发育完全, 前人研究也显示叶片完全发育前其CAT活性持续增加[33], 而对于突变体来说, 要及时清除过多的H2O2, 势必增加CAT活性。而突变体的CAT活性依次显著下降(图2-C), 致使细胞中H2O2不能及时被清除而累积(图3-B)。至于突变体剑叶、倒二叶和倒三叶的SOD和POD活性显著高于野生型且依次显著升高的原因,可能是叶片在衰老初期的自我保护, 诱导了SOD和 POD合成相关基因的表达(图2-A和B), 该结果进一步证实了汪媛[34]和赵晨晨等[35]的研究。当然, 叶片中高浓度的H2O2和O2-也将导致突变体倒二叶和倒三叶中的MDA含量显著升高, 分别比剑叶增加8.29%和76.93% (图3-C)。
叶片衰老既受生物和非生物等因素的影响, 也受植物内源激素的调节。研究已证实, 乙烯、油菜素甾醇、脱落酸(ABA)、水杨酸、茉莉酸及其衍生物茉莉酸甲酯等均是叶片衰老促进激素[36]。其中ABA合成的关键限速步骤是9-顺-环氧类胡萝卜素(9-cis-epoxycarotenoids)被裂解成C15的ABA前体黄质醛, 此步骤涉及的酶是9-顺-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenase, NCED)[36]。一旦内源ABA水平增加, 将诱导NAC类转录因子如AtNAP[37]和OsNAP[10]的表达, 并进一步促进其靶基因SAG113的表达[38], 进而抑制叶片气孔关闭, 加快叶肉细胞呼吸及水分蒸发, 最终导致叶片衰老。此外, 叶片衰老还受其他衰老相关基因的精细调控,其中水稻DNA结合蛋白OsWRKY42可以与OsMT1d的启动子区域结合, 进而抑制其表达并诱导ROS反应, 促进叶片衰老[12]; 抑制水稻锌指蛋白基因OsDOS[39]或OsTZF1[40]的表达则会引起水稻叶片早衰; 过量表达水稻GTP酶基因OsRab7B3也导致叶片早衰[41]。在本研究的OsPLS3基因定位区间内, 尚没有衰老相关的基因被定位或克隆, 而与报道相关的导致水稻叶片衰老的基因分别为锌指蛋白基因(LOC_ Os12g42250)、9-顺-环氧类胡萝卜素双加氧酶基因(LOC_Os12g42280)、GTP酶基因(LOC_Os12g42370)和DNA结合蛋白基因(LOC_Os12g42420)。研究表明锌指蛋白转录因子[42]及DNA结合因子[43]可以通过与清除ROS的CAT互作, 调控叶肉细胞的ROS反应来控制叶片衰老。本研究突变体的MDA含量、CAT活性、O2-含量、H2O2含量等结果表明叶片衰老很可能与ROS反应密切相关, 因此初步推测候选基因很可能是锌指蛋白基因(LOC_Os12g42250)或DNA结合蛋白基因(LOC_Os12g42420)。当然, 候选基因的最终确定要依赖于这两个基因的测序分析及遗传互补验证。
4 结论
ospls3是一个新的叶片早衰突变体。与野生型N142相比, 孕穗期突变体叶片中的叶绿素总量显著下降。伴随ospls3叶绿体的降解, 致使叶片中的O2-和H2O2大量产生。同时, 由于ospls3的CAT活性显著下降, 清除 H2O2的能力也显著下降, 并进一步引起MDA含量的极显著增加; 然而, 在叶片衰老的初期,SOD和POD活性显著升高。该突变受1对核隐性基因控制, 定位于第 12染色体 SSR标记 RM6953和RM28753之间约294 kb的范围内。
References
[1]Lim P O, Kim H J, Nam H G.Leaf senescence.Annu Rev Plant Biol, 2007, 58∶ 115-136
[2]伍泽堂.超氧自由基与叶片衰老时叶绿素破坏的关系.植物生理学通讯, 1991, 27∶ 277-279 Wu Z T.Relationship between superoxide radical and destruction of chlorophyll during leaf senescence.Plant Physiol Commun,1991, 27∶ 277-279 (in Chinese with English abstract)
[3]段俊, 梁承邺, 黄毓文.杂交水稻开花结实期间叶片衰老.植物生理学报, 1997, 23∶ 139-144 Duan J, Liang C Y, Huang Y W.Studies on leaf senescence of hybrid rice at flowering and grain formation stage.Acta Phytophysiol Sin, 1997, 23∶ 139-144 (in Chinese with English abstract)
[4]Zhang C J, Chu H J, Chen G X, Shi D W, Zuo M, Wang J, Lu C G,Wang P, Chen L.Photosynthetic and biochemical activities in flag leaves of a newly developed superhigh-yield hybrid rice (Oryza sativa) and its parents during the reproductive stage.J Plant Res, 2007, 120∶ 209-217
[5]Inada N, Sakai A, Kuroiwa H, Kuroiwa T.Senescence program in rice (Oryza sativa L.) leaves∶ analysis of the blade of the second leaf at the tissue and cellular levels.Protoplasma, 1999, 207∶222-232
[6]刘道宏.植物叶片的衰老.植物生理学通讯, 1983, (2)∶ 14-19 Liu D H.Plant leaf senescence.Plant Physiol Commun, 1983, (2)∶14-19 (in Chinese)
[7]Thomas H, Smart C M.Crops that stay green.Ann Appl Biol,1993, 123∶ 193-219
[8]魏道智, 戴新宾, 许晓明, 张荣铣.植物叶片衰老机理的几种假说.广西植物, 1998, 18(1)∶ 90-97 Wei D Z, Dai X B, Xu X M, Zhang R X.Several hypotheses on the mechanism of the plant leaf senescence.Guihaia, 1998, 18(1)∶90-97 (in Chinese with English abstract)
[9]Wang Z, Wang Y, Hong X, Hu D, Liu C, Yang J, Li Y, Huang Y,Feng Y, Gong H, Li Y, Fang G, Tang H, Li Y.Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1)induces early leaf senescence and defence responses in rice.J Exp Bot, 2015, 66∶ 973-987
[10]Liang C, Wang Y, Zhu Y, Tang J, Hu B, Liu L, Ou S, Wu H, Sun X, Chu J, Chu C.OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice.Proc Natl Acad Sci USA, 2014, 111∶ 10013-10018
[11]Chen L, Wuriyanghan H, Zhang Y, Duan K, Chen H, Li Q, Lu X,He S, Ma B, Zhang W, Lin Q, Chen S, Zhang J.An S-domain receptor-like kinase, OsSIK2, confers abiotic stress tolerance and delays dark-induced leaf senescence in rice.Plant Physiol, 2013,163∶ 1752-1765
[12]Han M, Kim C, Lee J, Lee S, Jeon J.OsWRKY42 represses OsMT1d and induces reactive oxygen species and leaf senescence in rice.Mol Cells, 2014, 37∶ 532-539
[13]Gao Q, Yang Z, Zhou Y, Yin Z, Qiu J, Liang G, Xu C.Characterization of an Abc1 kinase family gene OsABC1-2 conferring enhanced tolerance to dark-induced stress in rice.Gene, 2012,498∶ 155-163
[14]Park S, Yu J, Park J, Li J, Yoo S, Lee N, Lee S, Jeong S, Seo H S,Koh H, Jeon J, Park Y, Paek N.The senescence-induced stay green protein regulates chlorophyll degradation.Plant Cell, 2007,19∶ 1649-1664
[15]Jiang H, Li M, Liang N, Yan H, Wei Y, Xu X, Liu J, Xu Z, Chen F, Wu G.Molecular cloning and function analysis of the stay green gene in rice.Plant J, 2007, 52∶ 197-209
[16]Lee R, Hsu J, Huang H, Lo S, Chen S G.Alkaline alpha-galactosidase degrades thylakoid membranes in the chloroplast during leaf senescence in rice.New Phytol, 2009, 184∶ 596-606
[17]Jiao B, Wang J, Zhu X, Zeng L, Li Q, He Z.A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice.Mol Plant, 2012, 5∶ 205-217
[18]Sato Y, Morita R, Katsuma S, Nishimura M, Tanaka A, Kusaba M.Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice.Plant J, 2009, 57∶ 120-131
[19]Kusaba M, Ito H, Morita R, Iida S, Sato Y, Fujimoto M, Kawasaki S, Tanaka R, Hirochika H, Nishimura M, Tanaka A.Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence.Plant Cell, 2007, 19∶ 1362-1375
[20]Morita R, Sato Y, Masuda Y, Nishimura M, Kusaba M.Defect in non-yellow coloring 3, an alpha/beta hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice.Plant J, 2009, 60∶ 1110-1110
[21]Yamatani H, Sato Y, Masuda Y, Kato Y, Morita R, Fukunaga K,Nagamura Y, Nishimura M, Sakamoto W, Tanaka A, Kusaba M.NYC4, the rice ortholog of Arabidopsis THF1, is involved in the degradation of chlorophyll protein complexes during leaf senescence.Plant J, 2013, 74∶ 652-662
[22]Qiao Y, Jiang W, Lee J, Park B, Choi M, Piao R, Woo M, Roh J,Han L, Paek N, Seo H S, Koh H.SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunitμ1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa).New Phytol, 2010, 185∶ 258-274
[23]Undan J R, Tamiru M, Abe A, Yoshida K, Kosugi S, Takagi H,Yoshida K, Kanzaki H, Saitoh H, Fekih R, Sharma S, Undan J,Yano M, Terauchi R.Mutation in OsLMS, a gene encoding a protein with two double-stranded RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (Oryza sativa L.).Genes Genet Syst, 2012, 87∶ 169-179
[24]张治安, 陈展宇.植物生理学实验技术.长春∶ 吉林大学出版社, 2008.p 7 Zhang Z A, Chen Z Y.Experiment Technology of Plant Physiology.Changchun∶ Jilin University Press, 2008.p 7 (in Chinese)
[25]Rogers S O, Bendich A J.Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues.Plant Mol Biol, 1985, 5∶ 69-76
[26]Shen Y J, Jiang H, Jin J P, Zhang Z B, Xi B, He Y Y, Wang G,Wang C, Qian L L, Li X, Yu Q B, Liu H J, Chen D H, Gao J H,Huang H, Shi T L, Yang Z N.Development of genome-wide DNA polymorphism database for map-based cloning of rice genes.Plant Physiol, 2004, 135∶ 1198-1205
[27]Panaud O, Chen X, McCouch S R.Development of microsatellite markers and characterization of simple sequence length polymorphism (SSR) in rice (Oryza sativa L.).Mol Gen Genet, 1996,252∶ 597-607
[28]李晴, 朱玉贤.植物衰老的研究进展及其在分子育种中的应用.分子植物育种, 2003, 1∶ 289-296 Li Q, Zhu Y X.The progress of plant senescence research and plant molecular breeding.Mol Plant Breed, 2003, 1∶ 289-296 (in Chinese with English abstract)
[29]刘贞琦, 刘振业, 马达鹏, 曾淑芬.水稻叶绿素含量及其与光合速率关系的研究.作物学报, 1984, 10∶ 57-62 Liu Z Q, Liu Z Y, Ma D P, Zeng S F.A study on the relation between chlorophyll content and photosynthetic rate of rice.Acta Agron Sin, 1984, 10∶ 57-62 (in Chinese with English abstract)
[30]林植芳, 李双顺, 林桂珠, 孙谷畴, 郭俊彦.水稻叶片的衰老与超氧物歧化酶活性及脂质过氧化作用的关系.植物学报,1984, 26∶ 605-615 Lin Z F, Li S S, Lin G Z, Sun G C, Guo J Y.Superoxide dismutase activity and lipid peroxidation in relation to senescence of rice leaves.Acta Bot Sin, 1984, 26∶ 605-615 (in Chinese with English abstract)
[31]Hideg E, Kalai T, Kos P B, Asada K, Hideg K.Singlet oxygen in plants∶ its significance and possible detection with double (fluorescent and spin) indicator reagents.Photochem Photobiol, 2006,82∶ 1211-1218
[32]华春, 王仁雷.杂交稻及其三系叶片衰老过程中 SOD、CAT活性和MDA含量的变化.西北植物学报, 2003, 23∶ 406-409 Hua C, Wang R L.Changes of SOD and CAT activities and MDA content during senescence of hybrid rice and three lines leaves.Acta Bot Boreali-Occident Sin, 2003, 23∶ 406-409 (in Chinese with English abstract)
[33]金杨, 周丽芬, 陈析丰, 刘峰, 马伯军.水稻类病变突变体spl5细胞坏死机制的分析.浙江师范大学学报(自然科学版),2009, 32∶ 326-331 Jin Y, Zhou L F, Chen X F, Liu F, Ma B J.Mechanisms of cell death in rice lesion mimic mutant spl5.J Zhejiang Norm Univ (Nat Sci), 2009, 32∶ 326-331 (in Chinese with English abstract)
[34]汪媛.水稻叶片衰老过程生理变化及蛋白质降解与蛋白酶活性变化研究.扬州大学硕士学位论文, 江苏扬州, 2010 Wang Y.The Research of Physiological Changes, Protein Degradation and Protease Activity in the Process of Leaf Senescence in Rice.MS Thesis of Yangzhou University, Yangzhou, China, 2010 (in Chinese with English abstract)
[35]赵晨晨, 黄福灯, 龚盼, 杨茜, 程方民, 潘刚.水稻叶片早衰突变体 osled的生理特征与基因定位.作物学报, 2014, 40∶1946-1955 Zhao C C, Huang F D, Gong P, Yang Q, Cheng F M, Pan G.Physiological characteristics and gene mapping of a leaf early-senescence mutant osled in rice.Acta Agron Sin, 2014, 40∶1946-1955 (in Chinese with English abstract)
[36]许智宏, 薛红卫.植物激素作用的分子机理.上海∶ 上海科学技术出版社, 2012.pp 403-417 Xu Z H, Xue H W.Molecular Mechanism of Plant Hormones.Shanghai∶ Shanghai Scientific and Technical Publisher, 2012.pp 403-407 (in Chinese)
[37]Guo Y F, Gan S S.AtNAP, a NAC family transcription factor, has an important role in leaf senescence.Plant J, 2006, 46∶ 601-612
[38]Zhang K, Xia X, Zhang Y, Gan S.An ABA-regulated and Golgi-localized protein phosphatase controls water loss during leaf senescence in Arabidopsis.Plant J, 2012, 69∶ 667-678
[39]Kong Z, Li M, Yang W, Xu W, Xue Y.A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice.Plant Physiol, 2006, 141∶ 1376-1388
[40]Jan A, Maruyama K, Todaka D, Kidokoro S, Abo M, Yoshimura E, Shinozaki K, Nakashima K, Yamaguchi-Shinozaki K.OsTZF1,a CCCH-tandem zinc finger protein, confers delayed senescence and stress tolerance in rice by regulating stress-related genes.Plant Physiol, 2013, 161∶ 1202-1216
[41]Pitakrattananukool S, Kawakatsu T, Anuntalabhochai S, Takaiwa F.Overexpression of OsRab7B3, a small GTP-binding protein gene, enhances leaf senescence in transgenic rice.Biosci Biotechnol Biochem, 2012, 76∶ 1296-1202
[42]Li Y, Chen L, Mu J, Zuo J.LESION SIMULATING DISEASE1 interacts with catalases to regulate hypersensitive cell death in Arabidopsis.Plant Physiol, 2013, 163∶ 1059-1070
[43]Smykowski A, Zimmermann P, Zentgraf U.G-Box binding factor1 reduces CATALASE2 expression and regulates the onset of leaf senescence in Arabidopsis.Plant Physiol, 2010,153∶ 1321-1331
DOI:10.3724/SP.J.1006.2016.00667
*通讯作者(
Corresponding author)∶ 潘刚, E-mail∶ pangang12@126.com**同等贡献(Contributed equally to this work)
收稿日期Received()∶ 2015-09-12; Accepted(接受日期)∶ 2016-01-11; Published online(网络出版日期)∶ 2016-01-19.
Physiological Characteristics and Gene Mapping of a Precocious Leaf Senescence Mutant ospls3 in Rice
GONG Pan1,**, LI Kun-Yu1,**, HUANG Fu-Deng2, WEI Li-Quan1, YANG Xi1, CHENG Fang-Min1, and PAN Gang1,*
1College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310058, China;2Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
Abstract:Leaf senescence is the final stage of leaf development.However, premature aging of functional leaves leads to yield reduction and quality decline.Thus, it is very important for developing novel crop germplasms with delayed leaf-senescence characteristics through investigating the molecular mechanism of leaf senescence.In this study, an ospls3 (Oryza s ativa p recocious leaf senescence 3) mutant, produced by60Co γ-radiation treatment of indica cultivar N142, was identified.The symptoms of the premature senescence mutant presented firstly at tillering stage showing brown leaf tip and brown spots in top part of leaf blade, then spread rapidly to basal part of leaf blade and led leaf to die.The physiological analysis indicated that, in the ospls3 mutant, the content of chlorophyll was the highest in the flag leaf, the following was in second-top and third-top leaves, but all of them were significantly lower than those in the wild type.The contents of MDA, O2-, and H2O2and the activities of SOD and POD among the top three leaves in the wild type maintained similar levels, which were significantly lower than those in the mutant.The soluble protein contents and the activity of CAT had no significant difference among top three leaves in the wild type while significantly decreased in the mutant.Genetic analysis verified that the ospls3 is a recessive mutant and was mapped in a 294 kb interval between RM6953 and RM28753 on the long arm of chromosome 12, which establishes a solid foundation for further cloning and functional studies of this gene.
Keywords:Rice; ospls3; Precocious leaf senescence; Physiological analysis; Gene mapping
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27
中国土壤与肥料(2021年5期)2021-12-12
今日农业(2021年21期)2021-11-26
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-10-14
疯狂英语·初中天地(2021年6期)2021-08-06
金桥(2021年7期)2021-07-22
好孩子画报(2021年3期)2021-04-02
今日农业(2020年20期)2020-11-26
文苑(2020年6期)2020-06-22
