
西伯利亚幼鲟对偶极子电场的避让行为研究
2016-06-29 09:50杨全威张旭光郭弘艺宋佳坤上海海洋大学水产与生命学院海洋生物系统与神经科学研究所上海201306
水生生物学报 2016年3期
杨全威 张旭光 郭弘艺 张 博 宋佳坤(上海海洋大学水产与生命学院,海洋生物系统与神经科学研究所,上海 201306)
西伯利亚幼鲟对偶极子电场的避让行为研究
杨全威 张旭光 郭弘艺 张 博 宋佳坤
(上海海洋大学水产与生命学院,海洋生物系统与神经科学研究所,上海 201306)
摘要:研究利用人工偶极子电场来模拟生物电场刺激,对西伯利亚幼鲟的电感受能力进行了行为探究。结果显示西伯利亚幼鲟对本实验中的偶极子电场产生躲避行为,其平均感受阈值在2月龄为(457.5±32.5)μV/cm,3月龄为(29.5±2.5)μV/cm,7月龄为(10±1.0)μV/cm。这表明西伯利亚鲟鱼的电感受敏感性随个体的发育而增强,而这可能与电感受器官数量的增加有关(2月龄为2234±470,7月龄为5273±523)。
关键词:鲟鱼; 电感受; 避让行为; 阈值; 壶腹器官
电感受系统是一种古老的感觉系统,主要存在于大多数的软骨鱼类(Elasmobranch)[1]和软骨硬鳞鱼类(Chondrostean)[2—4],以及少数硬骨鱼类(Teleost)[5]中。电感受鱼类通过独特的电感受器官(壶腹器官)来感知周围生物体或非生物体的微弱电场信号,生物体的电场信号是由其呼吸、消化、运动等生命活动产生,在短距离内(<50 cm),电感受系统能够依靠感受到的微弱电场信号来进行定位,在其摄食和避害等行为中起着重要作用[1]。
目前,国内对鲟鱼电感受系统形态学和行为学等方面的研究表明,其电感受器官由套细胞、感觉毛细胞和支持细胞组成,中间充满透明的胶质黏液[6],鲟鱼能够对水下金属微弱腐蚀电场产生捕食行为[4,7]。同时,鲟鱼电感受系统的初级处理中枢脑核DON(Dorsal octavolateral nucleus)的信息编码机制也有一些研究,在电生理实验中,DON神经元对模拟饵料鱼生物电场的偶极子信号产生明显的相位耦合反应,并认为是一种鲟鱼捕食的行为机制[3]。研究表明,鱼类的生物电场具有偶极子电场的特征,其电场频率与其呼吸节律相对应[8],偶极子电场的强度与鱼的个体大小正相关,也就是说其敌害生物的电场强度大于其饵料生物电场。不过至今为止,仍不清楚鲟鱼对敌害生物电场的神经处理机制及行为表现。因此,在本研究中,利用人工偶极子产生较强的电场信号,以通过观察鲟鱼对高强度偶极子电场的行为反应特征(避让),来研究西伯利亚幼鲟(Acipenser baeri Brandt)对电场的感受敏感性以及不同发育时期的电场敏感性差异。
1 材料与方法
1.1 实验鱼
实验用西伯利亚幼鲟苗是由受精卵在实验室的孵化箱内孵化、生长而成。幼鲟在开口期主要投喂刚孵化出的卤虫,开口后饲养在200 L的循环水鱼缸内,缸内设置沙石、水草以及小型杂鱼和底栖生物(田螺、蟹等)以模拟自然水域环境。在鲟鱼生长过程中,选择2月龄、3月龄和7月龄的幼鲟进行偶极子电场刺激的行为实验,以及选择2月龄和7月龄的幼鲟进行电感受器官的形态观察。
1.2 行为记录方法
行为实验共分3组,2月龄组[N=13,TL=(6.2± 0.3)cm],3月龄组[N=13,TL=(9.2±0.5)cm],7月龄组[N=13,TL=(24.5±0.9)cm],分别在一个直径70 cm的玻璃水槽内进行,水槽放置在一个全封闭的隔音室内,以消除外界环境干扰。在水槽内壁一侧设置偶极子电场(图 1),并以偶极子为圆心在水槽底绘制不同半径的同心圆,并标记上实际值,以方便定量分析实验鱼的反应距离。水槽上方设置红外摄像头(670A3C,WOSHIDA,China)以记录实验鱼的行为反应,并在隔音室外进行远程观察。行为记录前,幼鲟在水槽中先适应10—20min,并以幼鲟围绕水槽内侧做规则的绕壁运动为适应标准。待实验鱼适应后,游至偶极子对侧时(距离电场最远处)进行偶极子电场刺激,并开始视频记录。

图 1 实验装置示意图
偶极子电场由间距0.5 cm的两根尖端裸露的银丝产生,其电流信号为10 Hz、1 V的正弦波信号,经由数模转换器(D/A converter,Micro 1401,CED,UK)和刺激隔离器(A395,WPI,USA)后输入至偶极子电极产生电场。偶极子在水中产生的实际电场采用一对间隔1 cm的Ag/AgCl电极进行测量和校正,记录的电场信号经过交流信号放大器(DAM 80,WPI,USA)放大1000倍(带通滤波1—300 Hz)后,传入数模转换器,并保存在计算机中。由信号分析软件(Spike 2)直接读取其峰峰幅值,算出电场强度。
为校正鲟鱼所感知的电场,实验前把测量点设置在鲟鱼的绕壁轨迹线附近,即以偶极子为中心的不同辐射半径与游泳轨迹线的交点。若把鲟鱼长吻端视为一条直线,这条线在鲟鱼游行过程中一直切向它的圆形轨迹,因此,本实验中的测量电极也以横切圆形轨迹的方向进行测量(图 1)。
1.3 电感受器官形态观察
先对西伯利亚幼鲟进行亚甲基蓝染色处理,把2月龄[N=5,TL=(6.5±0.3)cm]和7月龄[N=5,TL=(26.8±0.9)cm]西伯利亚幼鲟活体放入曝气的染色缸内,往染色缸里滴入配好的亚甲基蓝母液,直至水体呈深蓝色,但仍可看到鱼体,染色时间一般为24h。鲟鱼染色后再放回正常的养殖系统水中漂洗,并加入0.02%的MS-222麻醉处理。把麻醉后的西伯利亚鲟鱼放置在体式显微镜下观察(SteREO Discovery.V12),并对分布有壶腹器官的区域进行拍照,最后借助计数器进行数量统计。
1.4 数据分析
行为实验的视频数据采用逐帧法进行离线统计分析。分析时,将实验鱼在游向偶极子电场时突然出现停顿、折回、绕过等行为均视作避让行为,此时,实验鱼吻尖端与偶极子的距离则定义为避让距离(Avoiding distance),而实验鱼吻尖端处的电场值则定义为鲟鱼的避让阈值(Avoiding threshold),通过对比不同年龄段间的差异来了解幼鲟的电感受特性(Tukey test)。由于西伯利亚鲟鱼电感受器官数量较多,为方便统计,依据各壶腹器官与侧线管的分布关系将其划分成不同的区域进行计算[9],再比较两个年龄段壶腹器官数量的差异(t test)。所有数据均以平均值±标准误来表示,最后利用Sigmaplot(Version 12.5)进行绘制图表和显著性分析。
2 结果
2.1 电场分布模式
实验中,对鲟鱼绕壁轨迹处的电场强度进行了实际测量。结果表明,实验水槽内偶极子电场沿偶极子中线而对称分布,其中左侧4 cm处最大电场强度为379 μV/cm,右侧4 cm处测量的最大电场强度为438 μV/cm,右侧略大于左侧,这可能与使用的非标准偶极子电场源有关。电场强度随着与偶极子电极之间距离的增加而急剧地下降,在12 cm处平均电场强度已降至20 μV/cm,但此时电场强度减少幅度减弱,最终在36 cm以外的电场已经融合在环境背景电噪声中,无法进行精确测量(图 2)。
2.2 行为反应
实验分别测试了2月龄组(13条)、3月龄组(13条)、7月龄组(13条)3个不同年龄段的西伯利亚幼鲟对偶极子电场刺激的行为反应。结果表明,不同实验组的幼鲟对偶极子电场刺激表现出类似的规律性。在偶极子未通入电流时,幼鲟通常表现为规则的绕壁游动,并很少改变运动方向。当偶极子通入电流时,在远处的(偶极子对侧)鲟鱼并未出现异常反应,表现为正常的绕壁游动,当游向偶极子电场至一定距离时,大部分实验鱼会突然改变方向,加速逃离到远处,而后又开始恢复原来的绕壁行为(图 3)。但发现个别鲟鱼并未出现急剧地逃离行为,而表现突然停止,并在原处不停地徘徊。这些逃离或停止行为都被定义为避让行为。其中2月龄组的平均避让距离为(3.1±0.7)cm,平均避让阈值为(457.5±32.5)μV/cm,3月龄组的平均避让距离为(10.8±1.2)cm,平均避让阈值为(29.5±2.5)μV/cm,7月龄组的平均避让距离为(18.4±3.0)cm,平均避让阈值为(10±1.0)μV/cm。幼鲟对偶极子电场的避让距离随着生长发育而增加(Tukey test,P<0.001),避让阈值随着生长发育而降低,也就是随着年龄增长,幼鲟对偶极子电场的反应敏感性逐渐提高(图 4、图 5)。实验中也发现,实验鱼对偶极子电场存在适应性,即经过几次刺激后幼鲟接近偶极子电场的距离逐渐变小。
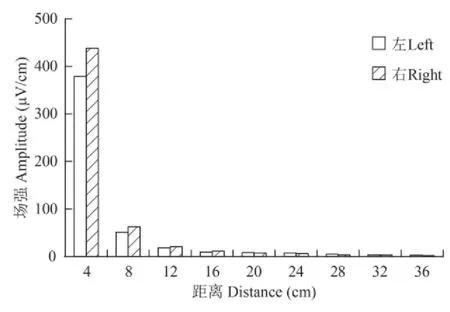
图 2 鲟鱼游泳轨迹处偶极子电场分布图
2.3 电感受器官数量的统计
实验分别选取了2月龄(5条)和7月龄(5条)的鲟鱼进行了形态观察。根据各壶腹分布区与侧线管的关系,西伯利亚鲟鱼壶腹区可分为:眶下管背区(di,dorsal infraorbital ampullary field)、眶下管腹区(vi,ventral infraorbital ampullary field)、眶上管背区(ds,dorsal supraorbital ampullary field)、眶上管腹区(vs,ventral supraorbital ampullary field)、前鳃盖管前区(app,anterior preopercular ampullary field)、前鳃盖管后区(ppp,posterior preopercular ampullary field)、耳壶腹区(ot,otic ampullary field)、颞上壶腹区(st,supratemporal ampullary field)(图 6)。不管是2月龄还是7月龄,其吻部腹面(di+vi)的壶腹器官平均分布数量最多,均占它们总数的一半以上。而分布数量最少的是2月龄的ot区,平均只有14个,甚至在其中个别鲟鱼此处并没有发现壶腹器官。结合两个年龄段鲟鱼壶腹器官的个体总平均值来看,电感受器官在此阶段出现明显增加的现象(t test,P=0.003)(表 1)。
3 讨论
鲟鱼具有电感受功能已经被人们证实,比如国内的梁旭方于九十年代在中华鲟(Acipenser sinensis Gray)的头部发现了大量的与电感受功能相关的感受器官即壶腹器官,同时行为上也表明了其的确能够利用这种特殊感觉系统来帮助捕食[7]。而近年来张旭光等人也发现西伯利亚鲟鱼对水中的微弱电场出现明显的捕食行为[4]。

图 3 鲟鱼对偶极子的避让行为(右侧白色区域为偶极子位置)
一般来说,电感受鱼类可通过电感受系统感知其它生物产生的电场来区分饵料或敌害[1,10,11]。鱼类生物电场具有偶极子电场的特征,其电场的大小与鱼体的长度有关,即体型较大的鱼的生物电场也较大。如张旭光等人描述了2 cm左右的食蚊鱼(Gambusia affinis Baird and Girard)的生物电场特征为头负尾正的偶极子,其最大交流信号约为4 μV[8],但Kalmijn也发现牛眼鲷(Boops salpa Cuvier)的生物电场为嘴负咽正的偶极子电场,其嘴部电势达+2.5 mV,咽部达-7.5 mV[12]。本实验中的偶极子电场的最大电场强度为500 μV/cm,如果以头负尾正的偶极子为例,那么该生物电场可近似为一种体型较大的鱼类。

图 4 不同年龄段西伯利亚幼鲟平均躲避距离(平均值±标准误)
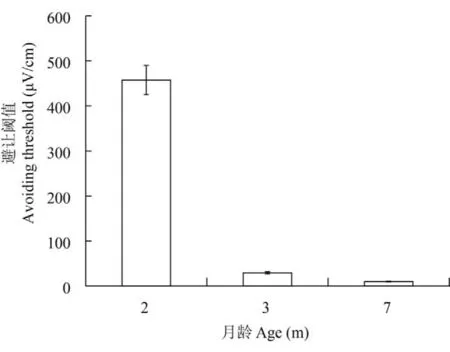
图 5 不同年龄段的西伯利亚幼鲟的避让阈值(平均值±标准误)
铝棒产生的腐蚀电场能引起西伯利亚鲟鱼产生吮吸行为[8],但在本实验中,西伯利亚鲟鱼对偶极子电场表现出了明显的避让反应。一方面,可能是实验鱼存在个体差异和实验鱼的体长差异; 另一方面电场的性质存在差异,同时电场强度也存在差异。距离铝棒表面5 mm处的腐蚀电场的峰值为90 μV,根据测量方法可换算为电场强度1.3—1.8 μV/ cm,鲟鱼的饵料鱼生物电场中的交流信号为4 μV左右,实验中测得最大的偶极子电场强度500 μV/cm(10 Hz),明显大于铝棒的腐蚀电场和饵料鱼的生物电场。然而,在实验中,7月龄幼鲟的平均避让阈值仅为10 μV/cm左右,与小型鱼类的生物电场信号接近。一方面,可能是鲟鱼实际的生理感受阈值要远远低于行为阈值。电生理的研究也表明,5 μV/cm的偶极子电场能引起电感受神经元放电序列的改变[3]; 另一方面,虽然水中电场随距离急剧衰减至一个较弱的信号,但鲟鱼通过电感觉系统能够根据实际感知的电场来分辨或推测出原电场源的性质或大小,从而判断出该电场是否来自饵料或敌害生物,对鲟鱼的电感受定位的初步研究也表明,鲟鱼能通过电感受系统来解析电场和距离的关系。
鲟鱼的侧线系统,包括机械感受和电感受系统,在2月龄时已经发育完善[13],但本次的实验结果表明,鲟鱼电感受器官的数量却在机体发育过程中还在继续增加,其对应的电感受功能也在不断地增强。根据之前提出的一种假说,即电感受的能力与机体电感受器官的分布密度、数量及壶腹器官管道的长度成正比[14],这样,在不同发育阶段的电感受器数量的差异就决定了其电感受敏感性的差异。因此,在本实验中,2月龄至7月龄的西伯利亚幼鲟的电感受器官数量的增加导致了其电感受敏感性的增强。7月龄幼鲟的电感受阈值(10 μV/cm)与其他淡水的被动电感受鱼类的感受阈值相比略大些,如匙吻鲟(Polyodon spathula Walbaum)的感受阈值低于1 μV/cm,鲶鱼(Ictalurus nebulosus Lesueur)的电感受阈值为1 μV/cm[15—17],这可能与鲟鱼的摄食习性相关,如匙吻鲟以浮游动物为食,其生物电场相对来说也较弱[17]。鲟鱼是一种洄游性的鱼类,有些可进行降海洄游,比如中华鲟。一般来说海水电感受鱼类的电感受性要更敏感,比如鲨鱼和鳐鱼的感受阈值为0.005—0.05 μV/cm,这是由于海水极强的导电性,电场在传播过程中会衰减地更严重,海水鱼为适应独特的海洋环境而进化出更敏感的电感受功能[18]。因此,那些降海洄游的鲟鱼(如中华鲟)随着个体发育其电感受敏感性可能还会继续增强。
近年来,随着人类社会的发展,人们对鱼类生存环境的污染越来越大,包括电磁污染,比如在一些河流或海域建设的大坝或发电站,及在水底或水上铺设高压电缆。水利设施中的金属结构在水中能够产生腐蚀电场,而那些发电站或高压电缆由于产生的是高频交流电,这种变化的电流能够导致周围水中也产生同等频率的电场,这些电场强度都远远大于鲟鱼的正常饵料或同类的生物电场,从而干扰它们或其他电感受鱼类的正常生活。Basov在伏尔加河一个水电站的水域附近检测到大于5000 μV/ cm的电场,而在距离其300 m以外的地方还依然能够测到3—4 μV/cm的电场,这些电场改变了在此生活的鲟鱼原有栖息规律[19]。另外,这对洄游性的电感受鱼更是一种新的威胁。如在1984年密西西比河段的一个水闸前聚集了大量的匙吻鲟鱼,因为悬在水中间的金属挡板让它们畏惧不前,而只有当金属挡板完全离开水面时,它们才肯通过水闸[20]。在我国,由于长江流域水利工程的建设,与西伯利亚鲟鱼同属的中华鲟也同样面临着这样的电磁污染的威胁。本研究中关于西伯利亚幼鲟的电感受行为反应能力的结果能够在一个新的角度对中华鲟的保护提供一定的科学参考。
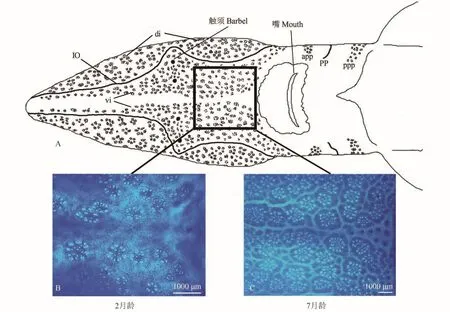
图 6 电感受器官在西伯利亚幼鲟头部腹面的分布图

表 1 壶腹器官在不同年龄段和区域的数量分布Tab.1 Average numbers of ampullary organs at different body regions of 2 age groups of sturgeons
参 考 文 献:
[1]Kajiura S M,Fitzgerald T P.Response of juvenile scal-loped hammerhead sharks to electric stimuli[J].Zoology,2009,112(4):241—250
[2]Hofmann M H,Jung S N,Siebenaller U,et al.Response properties of electrosensory units in the midbrain tectum of the paddlefish(Polyodon spathula Walbaum)[J].Journal of Experimental Biology,2008,211(5):773—779
[3]Zhang X G,Herzog H,Song J K,et al.Response properties of the electrosensory neurons in hindbrain of the white sturgeon,Acipenser transmontanus[J].Neuroscience Bulletin,2011,27(6):422—429
[4]Zhang X G,Song J K,Fan C X,et al.Use of electrosense in the feeding behavior of sturgeons[J].Integrative Zoology,2012,7(1):74—82
[5]Roth A.Ampullary electroreceptors in catfish:Afferent fiber activity before and after removal of the sensory cells[J].Journal of Comparative Physiology,1973,87(3):259—275
[6]Song W,Song J K.Morphological structure and peripheral innervation of the lateral line system in the Siberian sturgeon(Acipenser baerii)[J].Integrative Zoology,2012,7(1):99—109
[7]Liang X F.Study on the structure and function of the ampullae on the ventral part of the snout of Chinese sturgeon(Acipenser sinensis Gray)[J].Oceanologia et Limnologia Sinica,1996,27(1):1—5[梁旭方.中华鲟吻部腹面罗伦氏囊结构与功能的研究.海洋与湖沼,1996,27(1):1—5]
[8]Zhang X G,Song J K,Zhang G S,et al.The bioelectric field of gambusia affinis[J].Acta Hydrobiologica Sinica,2011,35(5):823—828[张旭光,宋佳坤,张国胜,等.食蚊鱼的生物电场特征.水生生物学报,2011,35(5):823—828]
[9]Gibbs M A,Northcutt R G.Development of the lateral line system in the shovelnose sturgeon[J].Brain,Behavior and Evolution,2004,64(2):70—84
[10]Kajiura S M,Holland K N.Electroreception in juvenile scalloped hammerhead and sandbar sharks[J].Journal of Experimental Biology,2002,205(23):3609—3621
[11]Kajiura S M.Electroreception in neonatal bonnethead sharks,Sphyrna tiburo[J].Marine Biology,2003,143(3):603—611
[12]Kalmijn A J.Bioelectric fields in sea water and the function of the ampullae of lorenzini in elasmobranch fishes[J].Scripps Institution of Oceanography Reference Series,1972,72—83
[13]Song W,Song J K.Development of the lateral line system in juvenile Siberian sturgeon(Acipenser baerii)[J].Zoological Research,2012,33(3):261—270[宋炜,宋佳坤.西伯利亚鲟仔鱼侧线系统的发育.动物学研究,2012,33(3):261—270]
[14]Kajiura S.Head morphology and electrosensory pore distribution of carcharhinid and sphyrnid sharks[J].Environmental Biology of Fishes,2001,61(2):125—133
[15]Peters R C,Eeuwes L B,Bretschneider F,et al.On the electrodetection threshold of aquatic vertebrates with ampullary or mucous gland electroreceptor organs[J].Biological Reviews,2007,82(3):361—373
[16]Russell D F,Wilkens L A,Moss F.Use of behavioural stochastic resonance by paddle fish for feeding[J].Nature,1999,402(6759):291—294
[17]Wilkens L A,Hofmann M H,Wojtenek W.The electric sense of the paddlefish:a passive system for the detection and capture of zooplankton prey[J].Journal of Physiology-Paris,2002,96(5—6):363—377
[18]Kalmijn A J.Electric and magnetic field detection in elasmobranch fishes[J].Science,1982,218(4575):916—918
[19]Basov B M.On electric fields of power lines and on their perception by freshwater fish[J].Journal of Ichthyology,2007,47(8):656—661
[20]Southall P D,Hubert W A.Habitat use by adult paddlefish in the upper mississippi river[J].Transactions of the American Fisheries Society,1984,113(2):125—131
THE AVOIDING RESPONSE TO DIPOLE STIMULI IN JUVENILE STURGEON
YANG Quan-Wei,ZHANG Xu-Guang,GUO Hong-Yi,ZHANG Bo and SONG Jia-Kun
(Institute for Marine Biosystem and Neuroscience,Shanghai Ocean University,Shanghai 201306,China)
Abstract:Here,the electrosensory responses of juvenile siberian sturgeon to electric fields simulated by dipole were studied.The results showed that the fish can avoid the dipole electric fields,and significant differences in avoiding thresholds were observed among different age groups.The mean avoiding threshold was(457.5±32.5)μV/cm at 2 months old,and it was(29.5±2.5)μV/cm at 3 months old,and it reached(10±1.0)μV/cm at 7 months old.The morphological observation showed that the electroreceptors increased from 2 month(2234±470)to 7 month(5273±523)old.These results suggest that the electrosensory sensitivity enhanced with increased electroreceptors in the development of the juvenile sturgeon.
Key words:Sturgeon; Electrosensory; Avoiding behavior; Threshold; Ampullae
中图分类号:Q424
文献标识码:A
文章编号:1000-3207(2016)03-0501-06
doi:10.7541/2016.67
收稿日期:2016-01-11;
修订日期:2016-03-21
基金项目:国家自然科学基金(41406150); 上海高校青年教师培养资助计划[Supported by the National Natural Science Foundation of China(41406150); Special Research Fund for Supporting Young Teachers in Shanghai Education Institutions of China]
作者简介:杨全威(1990—),男,河南商丘人; 硕士; 主要研究方向为鱼类感觉神经与行为。E-mail:yangquanwei126@163.com
通信作者:宋佳坤,教授,E-mail:jksong@shou.edu.cn; 张旭光,E-mail:zhang_xuguang@163.com
猜你喜欢
渔业致富指南(2021年20期)2021-11-29
古今农业(2021年2期)2021-08-14
制造技术与机床(2019年9期)2019-09-10
西南交通大学学报(2018年6期)2018-12-18
河北遥感(2017年2期)2017-08-07
大自然探索(2017年1期)2017-02-14
衡阳师范学院学报(2016年3期)2016-07-10
电测与仪表(2015年13期)2015-04-09
陕西学前师范学院学报(2014年5期)2014-07-31
海外星云 (2001年27期)2001-05-22
