
不同生境下泽兰实蝇的寄生状况及其影响因素分析
2016-06-23 03:13刘钊,周伟
安徽农业科学 2016年11期
关键词:生境
刘 钊,周 伟
(1.河西学院农业与生物技术学院,甘肃张掖 734000;2.西南林业大学林学院,云南昆明 650224)
不同生境下泽兰实蝇的寄生状况及其影响因素分析
刘 钊1,2,周 伟2
(1.河西学院农业与生物技术学院,甘肃张掖 734000;2.西南林业大学林学院,云南昆明 650224)
摘要[目的]研究不同生境下泽兰实蝇的寄生状况及其影响因素。[方法]通过对云南大中山地区紫茎泽兰入侵生境中泽兰实蝇的寄生状况抽样调查,分别从环境因素和紫茎泽兰生长状况两方面,分析了影响泽兰实蝇寄生的因素。[结果]不同生境泽兰实蝇的寄生状况有显著差异,其中林区公路和林区空旷地寄生率、虫瘿数和虫瘿内幼虫数最高,显著大于华山松林和落叶阔叶林。乔木层盖度与泽兰实蝇的寄生率、虫瘿数、虫瘿重呈显著负相关,而灌木层盖度与泽兰实蝇的寄生率呈显著正相关。泽兰实蝇寄生率与紫茎泽兰均高和开花率均呈极显著的正相关。[结论]开阔的生境有助于泽兰实蝇的寄生和扩散,林下的光照条件对泽兰实蝇的寄生亦有影响。泽兰实蝇偏好寄生林区公路边和林区空旷地植株较大、分枝较多的植株,泽兰实蝇对花期的植株可能更具有趋性。
关键词紫茎泽兰;泽兰实蝇;生境;寄生率;乔木层盖度;开花率
紫茎泽兰(Eupatoriumadenophorum)是一种世界性恶性杂草[1],也是我国首批公布的危害最严重的入侵种之一[2]。紫茎泽兰隶属菊科(Compositae)泽兰属(Adenophorum),为多年生丛生型半灌木状草本,原产于中美洲墨西哥、哥斯达黎加等地[3-4],现已广泛分布于澳大利亚、夏威夷、新西兰、菲律宾、缅甸、越南、泰国、印度和尼泊尔等 30 多个国家和地区[5]。紫茎泽兰于20世纪40年代从中缅边境传入我国云南地区[6]。目前在我国云南、四川、广西、贵州、西藏广泛入侵,也在重庆和湖北有零星报道,其中以云南省的分布最广,且以每年60 km的速度向东、向北入侵[1,6-8],已经对我国西南地区乃至华中、华南地区的生物多样性构成了巨大的威胁,并给西南地区农林牧业生产带来了巨大的损失[9]。
泽兰实蝇(Procecidocharesutilis)是紫茎泽兰重要的专性寄生性天敌,隶属双翅目实蝇科。我国从20世纪80年代开始研究泽兰实蝇的生物学特性、安全性测试及繁殖释放。1984年率先从西藏聂拉木县樟木区将泽兰实蝇引入云南部分地区,用于控制紫茎泽兰。此后,泽兰实蝇又被引入四川、贵州等地区,用于紫茎泽兰的生物防治[10-11]。大量研究表明,泽兰实蝇能够抑制紫茎泽兰的种子产量、萌发率、分指数、株高、光合作用和生物量分配等,甚至导致很高的死亡率,能够抑制紫茎泽兰的蔓延危害[12-17]。目前的研究报道大多限于研究泽兰实蝇寄生对紫茎泽兰生长的影响,而泽兰实蝇在不同生境类型中的寄生状况及其影响因素研究则鲜见报道。笔者以云南大中山紫茎泽兰入侵生境为采样地,研究了不同生境下泽兰实蝇的寄生状况,旨在为应用泽兰实蝇进行生物防治,并提高其防治效果提供理论指导。
1材料与方法
1.1研究区概况哀牢山国家级保护区南华片区系原云南省大中山保护区(简称大中山)(100°44′28″~100°57′42″ E,24°43′32″~25°01′10″ N),保护区总面积25 327 hm2,全年光照不足,气候垂直变化显著,从谷底到山顶高差1 750 m,上下温差在10 ℃以上,具有云南亚热带北部与亚热带南部过渡区特征和典型的山地气候特点。保护区主要有3个植被带,即海拔1 500 m以下为河谷稀树灌草丛及云南松林(Pinusyunnnanensis),海拔1 500~2 400 m 为半湿润阔叶林及云南松林,海拔2 400~2 714 m 为中山湿性常绿阔叶林,林区还有人工种植的华山松林(Pinusarmandii)[18]。
1.2数据收集2006年7~8月在南华片区进行为期32 d的野外调查与数据收集。在海拔2 200~2 500 m的范围内,紫茎泽兰主要分布于华山松林、林区公路边、落叶阔叶林和林区空旷地。在以上4种生境中,分别选取有紫茎泽兰分布的典型地段设置样地调查入侵情况[19],在每种生境各设 30 个样方,样方大小为 5 m×5 m[20],共120个。在 5 m×5 m大小的样方中测量海拔、坡度、坡向、乔木密度、乔木层盖度、灌木密度和灌木层盖度;在样方的中心及四角设立5个 1 m×1 m 的小样方,调查草本植被的多度和盖度[21]。在样方内统计的泽兰实蝇的寄生率和紫茎泽兰开花率,并采集10株优势紫茎泽兰全植株,共采集紫茎泽兰 1 200 株,统计并测定被寄生植株的虫瘿数、虫瘿重和虫瘿内幼虫数。
1.3数据处理采用Kolmogorov Smirnov Z-检验分析不同生境环境生态因子及泽兰实蝇寄生特征因子的正态性[22]。分析不同生境泽兰实蝇寄生状况和环境因子的差异性,当数据符合正态分布时,采用单因素方差分析和LSD多重比较法检验生境间差异性;否则,采用非参数Kruskal-WallisH检验和S-N-K多重比较法检验生境间差异性。采用相关性分析,探讨各环境因子对泽兰实蝇寄生状况的影响[22-23]。
所有数据处理均使用Excel 2003和SPSS 17.0统计软件。
2结果与分析
2.1不同生境泽兰实蝇的寄生状况比较单因素方差分析(F值)及非参数 Kruskal-WallisH检验(χ2值)结果表明,在不同生境中泽兰实蝇对紫茎泽兰的寄生率有显著差异(F=10.66,P<0.01),其中在林区公路的寄生率最高(P<0.01),其次是林区空旷地(P<0.01),在华山松林中的寄生率最低(P<0.01)。不同生境的泽兰实蝇虫瘿数有显著差异(χ2=2.83,P>0.05),林区公路和林区空旷地的虫瘿数较多(P<0.05),显著大于华山松林和落叶阔叶林(P<0.05)。不同生境泽兰实蝇虫瘿重有显著差异(χ2=25.51,P<0.01),林区公路边虫瘿重最大(P<0.01),林区空旷地虫瘿重较小。不同生境的泽兰实蝇幼虫数有显著差异(χ2=14.74,P<0.01),林区公路的幼虫数最多(P<0.05),其次是林区空旷地,华山松林和落叶阔叶林的幼虫数较少(P<0.05)(表1)。
2.2环境因子对泽兰实蝇寄生状况的影响Spearman相关性分析表明,泽兰实蝇对紫茎泽兰的寄生率与乔木层盖度呈极显著的负相关(r=-0.381,P<0.01),寄生率与灌木层盖度呈极显著的正相关(r=0.321,P<0.01)。寄生率(y)与乔木盖度和灌木盖度均符合一元线性回归模型(表2)。Spearman相关性分析表明,泽兰实蝇虫瘿数(r=-0.340,P<0.05)、虫瘿重(r=-0.336,P<0.05)、幼虫数(r=-0.413,P<0.01)与乔木层盖度均呈显著负相关。泽兰实蝇虫瘿数(r=0.425,P<0.01)、虫瘿重(r=0.267,P<0.05)、幼虫数(r=0.233,P<0.05)分别与灌木层盖度呈显著的正相关。泽兰实蝇寄生状况各项指标与乔木层盖度和灌木层盖度均符合一元线性回归模型(表2)。其中,乔木层盖度较低的林区公路边样方寄生率和虫瘿数较高;乔木盖度较高的华山松林和落叶阔叶林寄生率和虫瘿数较低(图1A,C)。林区公路边灌木盖度较高,寄生率和虫瘿数较高;灌木盖度较低的华山松林和落叶阔叶林,寄生率和虫瘿数较低(图1B,D)。
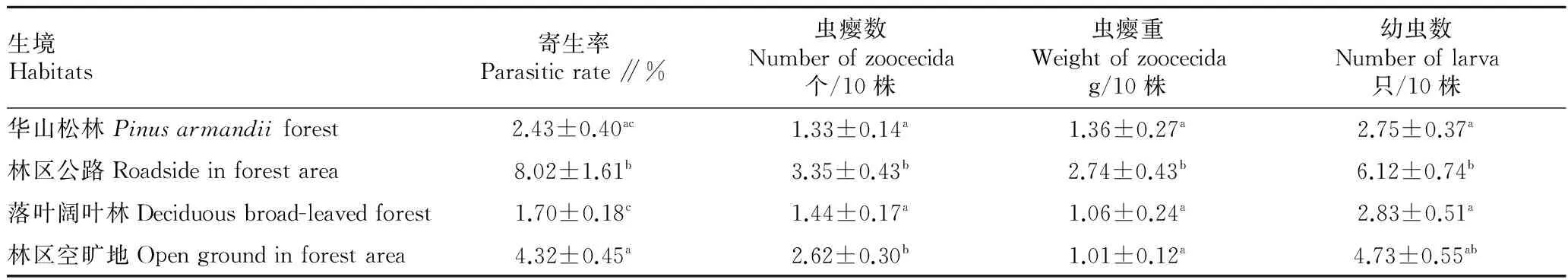
表1 不同生境紫茎泽兰的寄生情况比较
注:同列不同小写字母表示差异显著(P<0.05)。
Note:Different lowercases in the same column stand for significant differences(P<0.05).
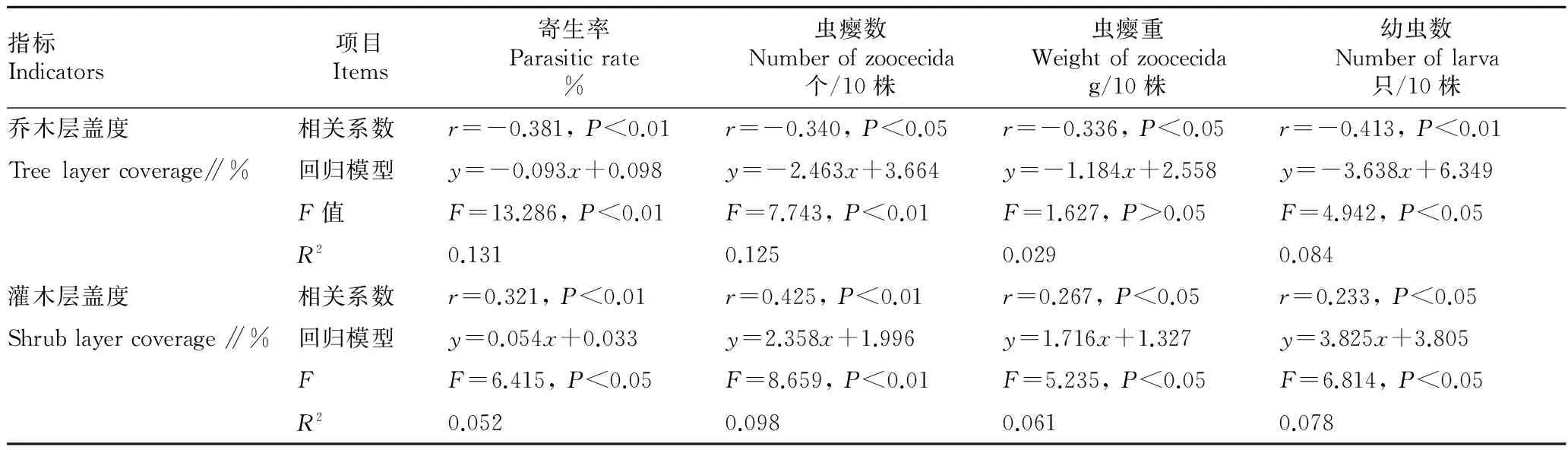
表2 泽兰实蝇寄生状况与环境因子的关系
2.3紫茎泽兰生长状况对寄生状况的影响从图2可以看出,泽兰实蝇寄生率与紫茎泽兰均高呈极显著的正相关(r=0.317,P<0.01),寄生率(y)与紫茎泽兰均高(x)符合一元线性回归模型y=0.001x-0.019(F=12.417,P<0.01)。泽兰实蝇寄生率和紫茎泽兰开花率呈极显著的正相关(r=0.405,P<0.01),寄生率(y)与开花率(x)符合一元线性回归模型y=0.104x+0.007(F=30.969,P<0.01)。泽兰实蝇虫瘿数与紫茎泽兰开花率呈极显著的正相关(r=0.318,P<0.01),虫瘿数(y)与紫茎泽兰开花率(x)符合一元线性回归模型y=2.310x+1.512(F=8.590,P<0.01)。林区公路边和空旷地中紫茎泽兰的高度和开花率较高,寄生率和虫瘿数也较高;华山松林中紫茎泽兰的均高和开花率较低,寄生率和虫瘿数则较低(图2A,B,C)。
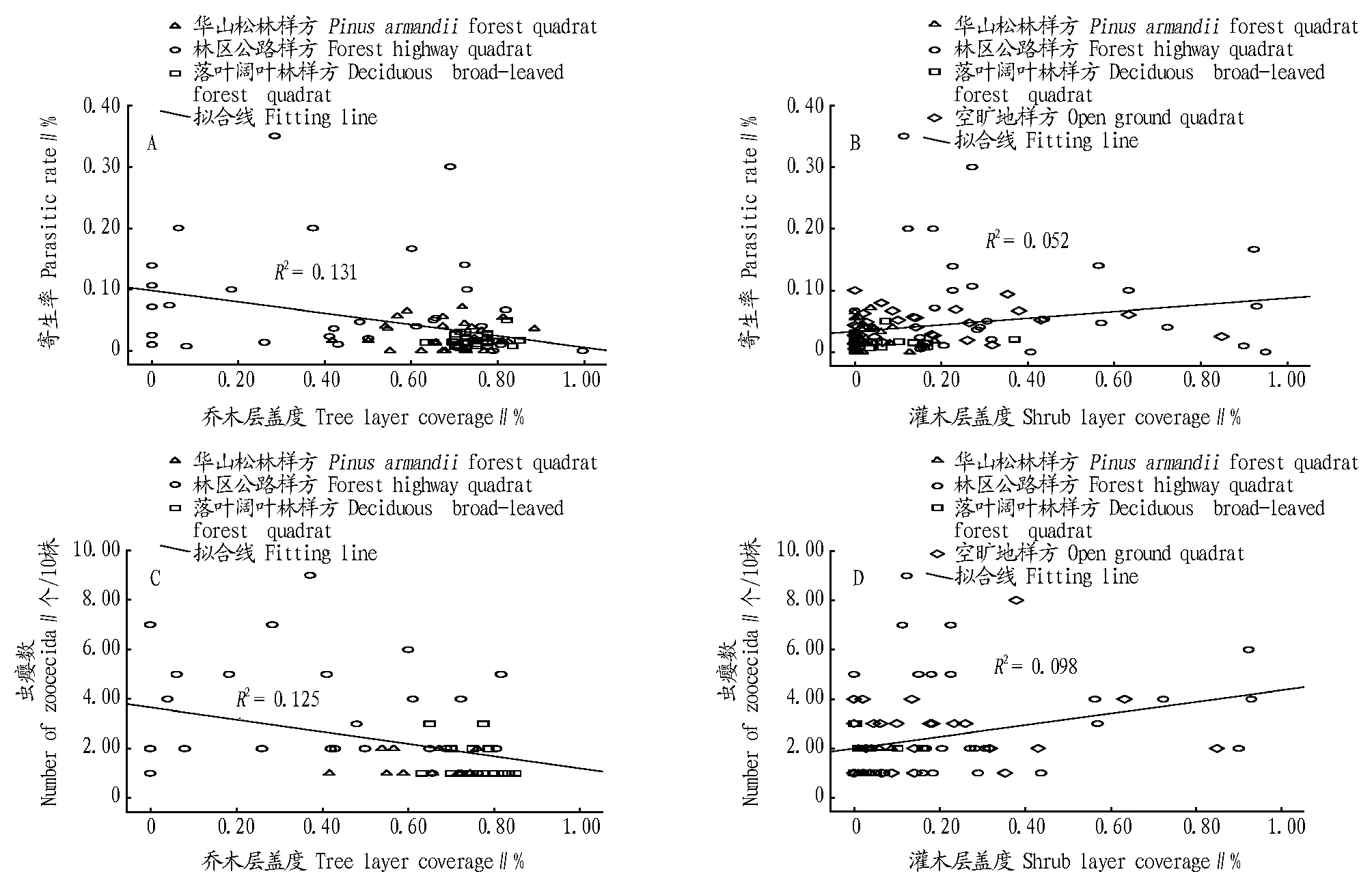
图1 泽兰实蝇寄生状况与乔木层盖度和灌木层盖度的关系Fig.1 The relationship between Procecidochares utilis parasitic status and tree layer coverage,shrub layer coverage
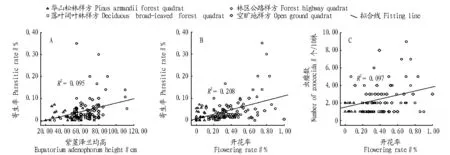
图2 泽兰实蝇寄生状况与紫茎泽兰均高和开花率的关系Fig.2 The relationship between Procecidochares utilis parasitic status and average height,flowering rate of Eupatorium adenophorum
2.4不同生境影响因子的比较由表3可知,单因素方差分析(F值)及非参数 Kruskal-WallisH检验(χ2值)结果表明,不同生境乔木层盖度有显著差异(F=124.30,P<0.01),落叶阔叶林和华山松林的乔木层盖度较大,显著大于林区公路(P<0.05)。不同生境灌木层盖度有显著差异(χ2=21.54,P<0.01),林区公路边的灌木层盖度最大,其次是林区空旷地,而华山松林灌木层盖度最小。不同生境紫茎泽兰均高存在显著差异(F=19.64,P<0.01),其中林区空旷地和林区公路边的紫茎泽兰均高较高,而华山松林样方最低(P<0.05)。不同生境紫茎泽兰开花率有显著差异(F=27.70,P<0.01),其中林区公路边和林区空旷地开花率最高,华山松林开花率最低。
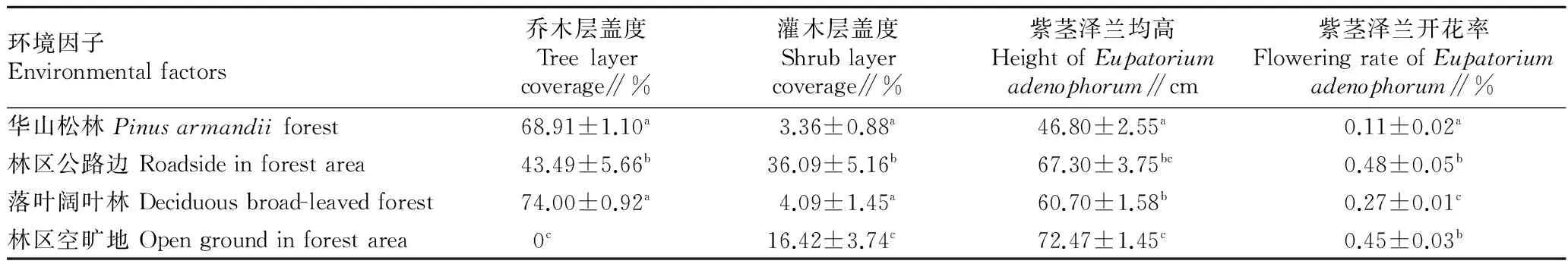
表3 紫茎泽兰不同生境影响因子的比较(n=30)
注:同列不同小写字母表示差异显著(P<0.05)。
Note:Different lowercases in the same column stand for significant differences(P<0.05).
3讨论与结论
3.1环境因素对泽兰实蝇寄生状况的影响不同的生境类型对泽兰实蝇的寄生状况有影响。以前对紫茎泽兰的调查研究多限于泽兰实蝇的寄生对植株生长发育及生物量分配的影响[15,24],比较不同生境类型中泽兰实蝇的寄生状况的研究较少,而泽兰实蝇在不同生境中的扩散速度和寄生强度均存在差异[17]。该试验结果表明泽兰实蝇在林区公路和林区空旷地的寄生率较高,虫瘿和幼虫的数量较大,这些生境特征有助于泽兰实蝇的寄生和扩散。紫茎泽兰的扩散速度远远大于泽兰实蝇的扩散速度,而环境因素对泽兰实蝇和紫茎泽兰的扩散都有一定的限制作用[7,16-17,25]。泽兰实蝇成虫虫体微小,易随风扩散[14],因此开阔的林区公路和空旷地有利于泽兰实蝇的扩散和寄生。林冠郁闭的落叶阔叶林中紫茎泽兰虽然植株高大,但寄生率不高,这也可能与林下的弱光照等物理条件有关。
3.2紫茎泽兰生长状况与泽兰实蝇寄生的关系紫茎泽兰的生长状况直接影响到泽兰实蝇的分布和寄生情况。泽兰实蝇是紫茎泽兰的专性天敌,有很强的搜索寄主的能力[14],说明紫茎泽兰的生长状况直接影响到泽兰实蝇的分布情况[17]。该试验结果表明不同生境中紫茎泽兰的构件生长和分枝格局差异很大,华山松林林冠郁闭,林下的紫茎泽兰植株较其他生境弱小,泽兰实蝇的寄生率较低;林区公路和林区空旷地紫茎泽兰植株高大,分枝较多,泽兰实蝇寄生率较高。紫茎泽兰的生长状况影响着泽兰实蝇的寄生。紫茎泽兰对寄主的选择力,是其发挥生物防控效果的关键[26]。研究表明,公路边和空旷地紫茎泽兰的开花率较高,泽兰实蝇的寄生率也较高,而在植株弱小的华山松林中寄生状况则相反。泽兰实蝇对花期的植株更具有趋性,因此花期的植株挥发物有待进一步研究,其中的某些有效成分能开发成提高泽兰实蝇寄生率的引诱物[26]。
参考文献
[1] 强胜.世界性恶性杂草:紫茎泽兰研究的历史及现状[J].武汉植物学研究,1998,16(4):366-372.
[2] 王俊峰,冯玉龙.人工群落中苗期紫茎泽兰的化感作用和对光环境的适应[J].植物生态学报,2006,26(6):1809-1817.
[3] AULD B A.The distribution ofEupatorium.adenophorumSpreng.on the far north coast of New South Wales[J].Journal and proceedings,royal society of New South Wales,1969,102:159-161.
[4] 刘伦辉,谢寿昌,张建华.紫茎泽兰在我国的分布、危害及防除途径的探讨[J].生态学报,1985,5(1):1-6.
[5] 王文琪,王进军,赵志模.紫茎泽兰种子种群动态及萌发特性[J].应用生态学报,2006,6(17):982-986.
[6] 赵国晶,马云萍.云南省紫茎泽兰的分布与危害的调查研究[J].杂草学报,1989,3(2):37-40.
[7] 刘伦辉,刘文耀,郑征,等.紫茎泽兰个体生物及生态学特性研究[J].生态学报,1989,9(1):66-70.
[8] 段惠.外来恶性杂草紫茎泽兰的入侵性若干方面及其控制方法的研究[D].南京:南京农业大学,2004.
[9] 卢志军,马克平.地形因素对外来入侵种紫茎泽兰的影响[J].植物生态学报,2004,28(6):761-767.
[10] 何大愚,刘伦辉,荆桂芬,等.泽兰实蝇的安全性试验[J].生物防治通报,1987,3(1):1-3.
[11] 张智英,魏艺,何大愚.泽兰实蝇生物学特性的初步研究[J].生物防治通报,1988,4(1):10-13.
[12] BESS H A,HARAMOTO F H.Biological control of Pamakani,Eupatoriumadenophorum,in Hawaii by a tephritid gall fly,Procecidocharesutilis.2.Population studies of the weed,the fly,and the parasites of the fly[J].Ecology,1959,40:244-249.
[13] BENNETT P H,VAN STADEN J.Gall formation in crofton weed,EupatoriumadenophorumSpreng(syn.Ageratinaadenophora)by theEupatoriumgall flyProcecidocharesutilisStone(Diptera:Trypetidae)[J].Australia journal of botany,1986,34(4):473-480.
[14] 陈旭东,何大愚.泽兰实蝇对紫茎泽兰的控制作用及其评价研究[J].杂草学报,1990,4(3):1-6.
[15] 刘文耀,刘伦辉,和爰军.泽兰实蝇对紫茎泽兰生长发育及生物量分配影响的研究[J].生态学报,1991,11(3):291-293.
[16] 王文琪,王进军,赵志模.紫茎泽兰种子种群动态及萌发特性[J].应用生态学报,2006,17(6):982-986.
[17] 李爱芳,高贤明,党伟光,等.泽兰实蝇寄生状况及其对紫茎泽兰生长与生殖的影响[J].植物生态学报,2006,30(3):496-503.
[18] 王裕康.南华大中山省级自然保护区综合科学考察报告[R].南华县林业局,2000.
[19] 卢志军,马克平.地形因素对外来入侵种紫茎泽兰的影响[J].植物生态学报,2004,28(6):761-767.
[20] 黄忠良,曹洪麟,梁晓东,等.不同生境和森林内薇甘菊的生存与危害状况[J].热带亚热带植物学报,2000,8(2):131-138.
[21] 于兴军,于丹,马克平.不同生境条件下紫茎泽兰化感作用的变化与入侵力关系的研究[D].植物生态学报,2004,28(6):773-780.
[22] QUINN G P,KEOUGH M J.Experimental design and data analysis for biologists[M].New York:Cambridge University Press,2002.奎恩,基奥.生物实验设计与数据分析[M].蒋志刚,李春旺,曾岩,译.北京:高等教育出版社,2003.
[23] FOWLER J,COHEN L,JARVIS P.Practical statistic for field biology[M].West Sussex:Open University Press,1998.
[24] 郑征,唐继武.泽兰实蝇对紫茎泽兰生长及光合作用影响的研究[J].杂草学报,1989,3(2):21-24.
[25] 孟秀祥,冯金朝,周宜君,等.四川西南紫茎泽兰 (Eupatoriumadenophorum)入侵生境因子分析[J].中央民族大学学报(自然科学版),2003,12(4):293-300.
[26] 马沙,吴国星,谭清,等.泽兰实蝇雌成虫的寄主选择行为[J].生物安全学报,2015(1):26-31.
Analysis of the Parasitic Status ofProcecidocharesutiliin Different Habitats and the Impact Factors
LIU Zhao1,2, ZHOU Wei2
(1. College of Agriculture and Biotechnology, Hexi University, Zhangye, Gansu 734000; 2. Department of Forestry, Southwest Forestry University, Kunming, Yunnan 650224)
Abstract[Objective] The aim was to study the parasitic status of Procecidochares utili in different habitats and the impact factors. [Method] The factors that influenced the parasitic status of Procecidochares utilis were analyzed from two sides, environmental factors and growth conditions of Eupatorium adenophorum, based on a sampling survey in different habitats which Procecidochares utilis had invaded in Dazhongshan Mountain in Yunnan Province. [Result] The results showed that there were significant differences of the parasitic status in different habitats. The parasitic rate of Procecidochares utilis, number of zoocecida and the number of larva in zoocecida at roadside and open ground in forest areas were significantly higher than those in Pinus armandii forests and deciduous broad-leaved forests. It was found that the tree layer coverage was positively correlated with parasitic rate, number of zoocecida, weight of zoocecida, respectively, and the shrub layer coverage was positively correlated with parasitic rate. The results also showed that parasitic rate was positively correlated with average height of Eupatorium adenophorum and flowering rate. [Conclusion] Procecidochares utilis will better disperse and parasite in open microhabitats and the light conditions under the tree canopy will affect the parasitism of Procecidochares utilis. Procecidochares utilis prefers to lay eggs first on taller plants with more branches and tend to choose Eupatorium adenophorum in the blooming period.
Key wordsEupatorium adenophorum; Procecidochares utilis; Habitats; Parasitic rate; Tree layer coverage; Flowering rate
基金项目国家重点基础研究发展计划项目(2002CB111407);云南省软科学重大项目(2004RK07)。
作者简介刘钊(1981- ),男,山东菏泽人,副教授,博士,从事野生动植物保护与利用。
收稿日期2016-03-21
中图分类号S 476.3
文献标识码A
文章编号0517-6611(2016)11-006-04
猜你喜欢
落叶果树(2021年6期)2021-02-12
生态学报(2018年12期)2018-08-10
现代园艺(2018年1期)2018-03-15
华东师范大学学报(自然科学版)(2017年1期)2017-02-27
体育科技(2016年2期)2016-02-28
海洋与湖沼(2016年2期)2016-01-15
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
东北林业大学学报(2012年5期)2012-09-18
