
瓯江口树排沙湿地不同生境大型底栖动物群落多样性研究*
2016-01-15 06:47胡成业水玉跃张春草覃胡林冀萌萌水柏年
海洋与湖沼 2016年2期
胡成业 水玉跃 田 阔 张春草 覃胡林 冀萌萌 水柏年①
(1. 浙江海洋大学水产学院 舟山 316000; 2. 舟山市海洋与渔业局 舟山 316000)
河口湿地是河流生态系统和海洋生态系统之间的生态交错带, 具有独特的结构与功能(童春富,2004)。频繁的人类活动对河口湿地生态系统的干扰较大, 河口湿地是一个生态敏感区。大型底栖动物是湿地生态系统的重要生态类群, 分解有机物, 转化营养物质, 污染物代谢和加速自净等, 在物质循环和能量流动中起着承上启下的作用(胡知渊等, 2009)。因此, 有关河口湿地大型底栖动物的研究倍受学者的关注(葛宝明等, 2005; 周晓等, 2006; 余骥等, 2013)。近年来, 瓯江口实施大规模的围填海工程, 造成湿地面积锐减。这不仅使湿地生物失去了栖息地, 直接影响到了沿岸渔业生产, 还导致沿岸生态环境恶化, 海岸灾害增多(张晓龙等, 2005)。为改善河口生态环境,龙湾区海洋与渔业局于2012年开始在瓯江口的树排沙湿地大片种植秋茄(Kandelia candel)、桐花树(Aegiceras corniculata)等红树植物, 红树林人工种植后, 需要对新生态系统进行有效的监控和管理, 其中就包括对新的红树林生态系统内的大型底栖动物进行详细的研究(陈光程等, 2008)。某些大型底栖动物具有较强的迁移能力, 在研究大型底栖动物群落时,仅考虑单一生境或只将某一生境与另一生境进行比较, 则无法充分认识造成生境中大型底栖动物在空间和时间上差异的原因, 也无法了解某个生境的变化会对整个区域生境内的底栖动物的影响(Morrisey et al, 2003; 陈光程, 2009)。鉴于此, 本研究选取了红树林、互花米草和光滩3种不同的生境, 通过物种多样性指数和 G-F多样性指数研究不同分类阶元水平上的多样性, 进而探讨种植红树植物对改善大型底栖动物多样性的效果, 以期为红树林湿地恢复提供相关基础资料。
1 材料与方法
1.1 研究区域
树排沙湿地位于瓯江口南支, 是携带入海的泥沙淤积而成, 水深 0—3m, 低潮时大片裸露成沙洲,是典型的河口沙洲湿地。为改善河口生态环境, 龙湾区海洋与渔业局于2012年逐渐引进秋茄、桐花树等红树植物, 因而造就了红树林、互花米草和光滩3种生境。目前, 是我国最北端大规模人工种植红树林的区域, 也是湿地水鸟的栖息地和迁徙地。
1.2 样品的采集、处理和分析
于 2014 年 10 月(秋季)、2014 年 12 月(冬季)、2015年3月(春季)2015年6月(夏季)对瓯江口树排沙湿地红树林、互花米草和光滩3种生境布设9条断面进行大型底栖动物定量采集(图1)。每条断面设10个样方,每个样方的面积为 25cm×25cm, 取样深度为 30cm。所取的沉积物用 0.5mm 孔径的分样筛进行淘洗, 获得的大型底栖动物用无水酒精固定后, 带回实验室进行分类鉴定、计数和称重, 采样与测定方法遵循国家标准《海洋调查规范》(GB/T12763.6-2007)。

图1 研究区域及采样点Fig.1 The study area and sampling sites
1.3 数据处理
1.3.1 物种多样性指数

式(1)、(2)和(3)中, S为总种数; N为所有种类的总尾数; Pi为第i种的尾数占总尾数的比例。
1.3.2 G-F多样性指数

式(4)、(5)和(6)中, qj为群落中j属大型底栖动物种数与总种数的比值, p为群落中总属数, m为群落中总科数, DFk为k科中的物种多样性, n为k科中的属数, pi为群落中k科i属中的种数占k科总种数的比值。
利用Excel 2007计算大型底栖动物栖息密度、生物量和G-F多样性指数, Primer 5.0计算物种多样性指数, SPSS 19.0进行单因素方差分析。
2 结果
2.1 种类组成及生活型
4个季度在3种生境中共鉴定出大型底栖动物48种, 隶属于5纲15目31科40属, 其中以甲壳类、腹足类和多毛类为主, 分别物种数的 36.73%、28.57%和18.37%。不同生境大型底栖动物的物种分布不同,其中在红树林生境物种数最多, 达 38种, 其次是光滩和互花米草生境, 分别为28种和25种。单因素方差分析表明, 物种数季节间的差异不显著(F=1.088,P=0.497), 物种数生境间的差异显著(F=7.268,P=0.013)。根据大型底栖动物的栖息空间将其划分为底上运动迟缓型、游泳型、潜穴型和穴居型(范航清等, 2000), 其中红树林生境底上运动迟缓型、游泳型、潜穴型和穴居型的种类依次有11种、5种、12种和10种; 互花米草生境底上运动迟缓型、游泳型、潜穴型和穴居型的种类依次有8种、5种、5种和7种; 光滩生境底上运动迟缓型、游泳型、潜穴型和穴居型的种类依次有7种、3种、10种和8种(表1)。
2.2 栖息密度和生物量
红树林生境中大型底栖动物的年平均栖息密度最高, 达84.60 ind./m2, 互花米草生境次之, 为54.43
ind./m2, 而光滩生境略低于互花米草生境, 为 53.70 ind./m2。四个季度均为红树林生境栖息密度高于互花米草生境和光滩生境, 其中秋季红树林生境的栖息密度最高(123.07 ind./m2), 夏季互花米草生境最低(20.67 ind./m2)。光滩生境栖息密度季节间的变化幅度最小, 而互花米草生境的变化幅度最大(图 2)。单因素方分析表明, 大型底栖动物栖息密度季节和生境间的差异均不显著(季节:F=1.986,P=0.195; 生境:F=1.625,P=0.250)。

表1 不同生境大型底栖动物种类组成及其生活型Tab.1 Species composition and life forms of macrobenthos in different habitats

图2 不同生境大型底栖动物栖息密度Fig.2 The inhabitant density of macrobenthos in different habitats
红树林生境中大型底栖动物的年平均生物量最高, 达63.98 g/m2, 互花米草生境次之, 为35.12 g/m2,而光滩生境略低于互花米草生境, 为34.59 g/m2。四个季度均为红树林生境生物量高于互花米草生境和光滩生境, 其中夏季红树林生境的生物量最高(105.07 g/m2), 冬季光滩生境最低(12.32 g/m2)。互花米草生境生物量季节间的变化幅度最小, 而红树林生境的变化幅度最大(图3)。单因素方差分析表明, 大型底栖动物生物量季节和生境间的差异均不显著(季节;F=3.540,P=0.068; 生境:F=1.922,P=0.202)。
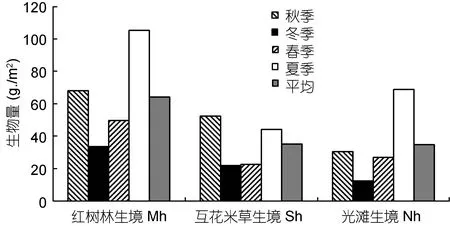
图3 不同生境大型底栖动物生物量Fig.3 The biomass of macrobenthos in different habitats

表2 不同生境大型底栖动物物种多样性指数Tab.2 The species diversity index of macrobenthos in different habitats
2.3 物种多样性
对大型底栖动物进行物种多样性分析结果表明(表 2), 不同生境间大型底栖动物 Shannon-Wiener多样性指数(H′)、Pielou 均匀度指数(J′)和 Margalef物种丰富度指数(D)变化幅度较大, 其中H′、J′和D均为红树林生境最高。从不同季节来看,H′最高值是春季红树林生境(2.21), 而最低值是冬季光滩生境(1.21);J′最高值是夏季光滩生境(0.77), 而最低值是冬季光滩生境(0.46);D最高值是秋季红树林生境(3.52), 而最低值是夏季光滩生境(1.90)。单因素方差分析表明,H′在不同季节间差异不显著(F=0.884,P=0.489), 在不同生境间差异显著(F=5.135,P=0.033);J′在不同季节和生境间差异均不显著(季节:F=2.936,P=0.099;生境:F=1.299,P=0.319);D在不同季节间差异不显著(F=0.327,P=0.806), 在不同生境间差异显著(F=5.152,P=0.032)。
2.4 G-F多样性指数
从全年来看,G指数红树林生境最高, 达3.46, 而互花米草生境最低, 仅2.98;F指数同样是红树林生境最高,达3.58, 而互花米草生境最低, 仅3.30;G-F指数互花米草生境最高, 达0.10, 而红树林生境最低, 仅0.03。从不同季节来看,G指数秋季红树林生境最高, 冬季互花米草生境和夏季光滩生境最低, 均为2.35;F指数春季互花米草生境和光滩生境最高, 均为 2.89, 夏季光滩生境最低,仅0.69;G-F指数春季光滩生境最高, 达0.15, 秋季红树林生境和互花米草生境最低, 均为–0.36(表3)。

表3 不同生境大型底栖动物G-F多样性指数Tab.3 The G-F diversity index of macrobenthos in different habitats
3 讨论
3.1 不同生境大型底栖动物栖息特征
大型底栖动物群落分布与温度、盐度、底质和植被等因子密切相关(寿鹿等, 2007), 由于研究区域为长度约3 km、宽度约0.5 km的河口沙洲湿地, 不同生境间的温度、盐度和底质等因子较相似, 因此植被类型是影响不同生境大型底栖动物种类分布的重要因子。单因素方差分析表明, 不同生境中大型底栖动物物种数差异显著。这与魏德重等(2012)针对乐清湾茅埏岛红树林生境、互花米草生境和光滩生境大型底栖动物物种数差异显著的研究结果相一致。此外, 红树林生境大型底栖动物的物种数最高, 以腹足类和甲壳类为主, 光滩生境次之, 而互花米草生境最少。与互花米草和光滩生境不同, 红树林复杂的生境为大型底栖动物尤其是腹足类和甲壳类提供了适宜的栖息环境。在野外调查时发现互花米草生境底质发臭,根系发达以及植株间隙较小等不良条件制约了多毛类、蟹类和双壳类等潜穴型动物的栖息。光滩生境多毛类物种数高于红树林生境和互花米草生境, 与此同时林俊辉等(2014)研究发现光滩生境多毛类的密度和生物量显著高于红树林生境。因此, 光滩生境可能更有利于多毛类栖息。
3.2 生境对物种多样性的影响
与邻近相同潮区其他生境相比, 红树林生境有助于提高林内大型底栖动物的物种多样性。在四个季节中, 红树林生境大型底栖动物群落的 Shannon-Wiener多样性指数、Pielou均匀度指数和 Margalef物种丰富度指数均高于互花米草生境和光滩生境。目前, 有关红树林对大型底栖动物物种多样性的影响原因主要有以下四点: (1) 红树林复杂的生境为大型底栖动物提供良好的栖息、繁育和庇护场所(Roach et al, 2000; 叶勇等, 2006); (2) 与其它栖息地相比红树林具有较高的生产力, 其凋落物为林下大型底栖动物主要是蟹类、腹足类和邻近水域生态系统食物链开端的消费者提供食物来源(Robertson, 1986; 陈雅萍等, 2013); (3) 红树林会改变林内土壤的性质, 如盐度、酸度和营养元素含量, 而这些底质因素是影响大型底栖动物组成和分布的重要因子(陈光程等, 2013);(4) 红树林对潮间带不利环境的改善也有利于大型底栖动物的分布(Dye, 1983; 陈光程等, 2013)。
对红树林生境、互花米草生境和光滩生境中大型底栖动物Shannon-Wiener多样性指数、Pielou均匀度指数和 Margalef物种丰富度指数分别进行生境和季节间单因素方差分析。结果表明, 不同生境大型底栖动物群落的 Shannon-Wiener指数和 Margalef物种丰富度指数差异显著。此外, 赵彩云等(2014)对广西北海西村港不同生境大型底栖动物群落研究发现Shannon-Wiener多样性指数和Simpson指数在红树林生境和互花米草生境间差异显著。由此可见, 生境间的差异是造成大型底栖动物物种多样性差异的主要原因之一。
3.3 属级和科级多样性
Shannon-Wiener多样性指数、Pielou均匀度指数和 Margalef物种丰富度指数能较好的反映群落的物种多样性, 被广泛应用于大型底栖动物的群落研究中。从分类阶元角度来看, 上述指数仅考虑了物种在种这一分类阶元上的多样性, 尚未涉及到属和科级分类阶元上的多样性。物种分类学关系往往是不均匀的, 即大型底栖动物的多样性在不同分类阶元上是不一致的, 仅研究物种多样性并不能较好的反映群落多样性。鉴于此, 蒋志刚等(1999)最早提出了 G-F多样性指数来反映物种在属级和科级分类阶元上的多样性。因此, 结合物种多样性指数和G-F多样性指数更能全面客观的反映大型底栖动物群落多样性。
本研究得出, 光滩生境大型底栖动物全年属级和科级的多样性指数均高于互花米草生境, 这与物种多样性指数相反。由此可见, 某一生境物种多样性高并不意味属级和科级的多样性就会高。因此, 在衡量某一生物群落的物种多样性时, G-F指数可以弥补Shannon-Weiner指数的不足(张淑萍等, 2002)。通常DG-F值一般情况是不会出现负数的, 但李海宏等(2015)研究茅埏岛不同生境大型底栖动物时发现DG-F值出现负数。在本研究中, 不同生境的大型底栖动物群落各个季节的DG-F值也出现负数。究其原因, 这主要与大型底栖动物种数较少, 且在科级分类阶元上的分布较分散, 造成属级和科级分类阶元上只出现一种大型底栖动物, 进而造成DG>DF有关。此外, 造成DG-F值为负数可能与 G-F多样性指数本身有关。史 赟 荣等(2010)研究发现, G-F多样性指数的关键函数Y = –X lnX不是单调函数, 是造成DG-F值为负数的原因之一。
叶 勇, 翁 劲, 卢昌义等, 2006. 红树林生物多样性恢复.生态学报, 26(4): 1243—1250
史 赟 荣, 李永振, 艾 红等, 2010. 西沙群岛珊瑚礁海域鱼类分类学多样性. 水产学报, 34(11): 1753—1761
寿 鹿, 高爱根, 曾江宁等, 2007. 底质环境对浙江衢山岛潮间带大型底栖动物分布的影响. 动物学杂志, 42(3):79—83
李海宏, 江 旷, 鲍毅新等, 2015. 茅埏岛不同生境大型底栖动物生物多样性. 生态学杂志, 34(3):765—772
余 骥, 马长安, 吕巍巍等, 2013. 崇明东滩潮间带大型底栖动物的空间分布与历史演变. 海洋与湖沼, 44(4): 1078—1085
张晓龙, 李培英, 李 萍等, 2005. 中国滨海湿地研究现状与展望. 海洋科学进展, 23(1): 87—95
张淑萍, 张正旺, 徐基良等, 2002. 天津地区水鸟区系组成及多样性分析. 生物多样性, 10(3): 280—285
陈光程, 2009. 九龙江口秋茄红树植被与主要大型底栖动物某些生态关系的研究. 厦门: 厦门大学博士学位论文, 5
陈光程, 叶 勇, 卢昌义, 2008. 九龙江口秋茄红树林恢复对大型底栖动物群落的影响. 厦门大学学报(自然科学版),47(2): 260—264
陈光程, 余 丹, 叶 勇等, 2013. 红树林植被对大型底栖动物群落的影响. 生态学报, 33(2): 327—336
陈雅萍, 叶 勇, 2013. 红树林凋落物生产及其归宿. 生态学杂志, 32(1): 204—209
范航清, 何斌源, 韦受庆, 2000. 海岸红树林地沙丘移动对林内大型底栖动物的影响. 生态学报, 20(5): 722—727
林俊辉, 郑凤武, 何雪宝等, 2014. 福建沿岸红树林湿地多毛类生态分布. 生态学报, 34(17): 4910—4919
周 晓, 王天厚, 葛振鸣等, 2006. 长江口九段沙湿地不同生境中大型底栖动物群落结构特征分析. 生物多样性, 14(2):165—171
赵彩云, 柳晓燕, 白加德等, 2014. 广西北海西村港互花米草对红树林湿地大型底栖动物群落的影响. 生物多样性,22(5): 630—639
胡知渊, 鲍毅新, 程宏毅等, 2009. 中国自然湿地底栖动物生态学研究进展. 生态学杂志, 28(5): 959—968
葛宝明, 鲍毅新, 郑 祥, 2005. 灵昆岛围垦滩涂潮沟大型底栖动物群落生态学研究. 生态学报, 25(3): 446—453
蒋志刚, 纪力强, 1999. 鸟兽物种多样性测度的G-F指数方法.生物多样性, 7(3): 220—225
童春富, 2004. 河口湿地生态系统结构、功能与服务——以长江口为例. 上海: 华东师范大学博士学位论文, 1
魏德重, 项长友, 鲍毅新等, 2012. 红树林种植对大型底栖动物群落结构及功能群的影响. 浙江师范大学学报(自然科学版), 35(2): 195—202
Dye A H, 1983. Composition and seasonal fluctuations of meiofauna in a Southern African mangrove estuary. Marine Biology, 73(2): 165—170
Morrisey D J, Skilleter G A, Ellis J I et al, 2003. Differences in benthic fauna and sediment among mangrove (Avicennia marina var. australasica) stands of different ages in New Zealand. Estuarine, Coastal and Shelf Science, 56(3—4):581—592
Roach A C, Lim R P, 2000. Variation in the population dynamics of the intertidal pulmonate gastropod Salinator solida Martens (Gastropoda: Amphibolidae) at Towra Point, NSW, Australia. Wetlands Ecology and Management,8(1): 53—69
Robertson A I, 1986. Leaf-burying crabs: Their influence on energy flow and export from mixed mangrove forests(Rhizophora spp.) in northeastern Australia. Journal of Experimental Marine Biology and Ecology, 102(2—3):237—248
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
儿童故事画报·自然探秘(2022年3期)2022-04-27
海洋通报(2021年5期)2021-12-21
幼儿园(2021年4期)2021-07-28
落叶果树(2021年6期)2021-02-12
能源与环境(2020年4期)2020-09-02
歌海(2018年4期)2018-05-14
体育科技(2016年2期)2016-02-28
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
