
柴达木盆地梭梭AQP基因克隆及结构预测
2016-06-16 01:58:09马玉花冶贵生马秀芳李玉英龙晓晨马娟冶桂莲
甘肃农业大学学报 2016年2期
马玉花,冶贵生,马秀芳,李玉英,龙晓晨,马娟,冶桂莲
(青海大学农牧学院,青海 西宁 810016)
柴达木盆地梭梭AQP基因克隆及结构预测
马玉花,冶贵生,马秀芳,李玉英,龙晓晨,马娟,冶桂莲
(青海大学农牧学院,青海 西宁810016)
摘要:【目的】 克隆得到柴达木盆地梭梭AQP基因并预测其结构.【方法】 以柴达木盆地梭梭同化枝为材料,RT-PCR扩增其AQP基因并进行序列测定.【结果】 所得柴达木盆地梭梭AQP基因的碱基数为481个,编码160个氨基酸.AQP亲水性、二级结构、跨膜螺旋区、三级结构预测显示柴达木盆地梭梭AQP二级结构主要为α螺旋和无规则卷曲.AQP有1个 N-糖基化位点,2个蛋白激酶C磷酸化位点,4个酪蛋白激酶Ⅱ磷酸化位点,2个N-端豆蔻酰化位点, 1个微体细胞C端靶信号和1个白细胞介素10家族信号.此外AQP蛋白的亲水性较弱,具有3个跨膜螺旋区.柴达木盆地梭梭AQP三维结构预测结果显示其主要由α螺旋和无规则卷曲互相盘绕而成,含有少量β折叠.【结论】 研究结果为下一步柴达木盆地梭梭AQP基因的表达特性及功能研究提供了依据.
关键词:梭梭;AQP基因;扩增;序列分析
梭梭(HaloxylonAammodendron)为藜科(Chenopodiaceae)梭梭属(HaloxylonBunge)超旱生盐生小乔木或大灌木,具有抗旱、耐盐碱等生态适应特征,广泛分布于亚洲和非洲荒漠地区,是荒漠地区主要的建群种和优良固沙植物.梭梭不仅是优良的薪炭材料,也是荒漠草场主要的饲料树种之一[1],梭梭还是肉苁蓉的寄主,因而对梭梭的研究和开发利用具有重要的生态、经济和社会价值.作为超旱生耐盐植物[2],关于梭梭抗逆研究主要集中在抗旱方面[3-5],耐盐机理方面的研究少有报道,而且主要集中在盐处理对梭梭种子萌发、幼苗生长、单一生理生化指标影响等研究[1,6-7],对梭梭抗旱耐盐相关基因的研究报道较少[8-12].
水通道蛋白(AQPs)是水选择性通透的膜内在蛋白,种子萌发、气孔运动、细胞伸长、生殖生长、韧皮部装卸和植物应激等生命过程都与水通道蛋白密切相关.目前已在多种植物中发现了AQPs,研究表明植物通过调节AQPs的活性协同微调水分的跨膜运输,进而维持高渗条件下的水分平衡以抵御各种逆境胁迫.目前有关梭梭AQP的研究尚未见报道.本文通过对柴达木盆地梭梭AQP基因进行克隆、序列分析和蛋白结构预测,研究结果可为下一步梭梭AQP的功能研究奠定基础.
1材料与方法
供试梭梭的种子采自青海省柴达木盆地,实验室育苗后取同化枝进行总RNA的提取.RNA提取试剂盒为北京艾德来生物技术有限公司产品,ExTaq DNA聚合酶、反转录酶、DL2000 DNA Marker购自宝生物工程(大连)有限公司.
参照试剂盒说明提取梭梭总RNA.根据已发表的AQP基因序列设计扩增引物,由大连宝生物工程有限公司合成,预期目的片段长度为481 bp.引物序列为:AQP-F:5′-CTYGTYTACTGCACHGCY-3′、AQP-R:5′-CCVACCCARAADATCCAN-3′.按试剂盒说明反转录后进行PCR扩增,扩增程序为:94 ℃ 1 min,61 ℃ 1 min,72 ℃ 1 min,35个循环,72 ℃ 10 min.PCR产物送交生物公司进行序列测定,应用DNASTAR软件进行序列分析,predictprotein预测蛋白的二级结构和亲水性及分析蛋白修饰位点,用TMHMM进行蛋白跨膜螺旋区分析,应用I-Tasser在线服务器预测蛋白的三级结构.
2结果与分析
2.1梭梭总RNA提取
试剂盒提取获得的梭梭总RNA经琼脂糖凝胶电泳得到梭梭RNA电泳图(图1),可见所得RNA条带清晰,28S为18S的2倍,可以满足后续试验的要求.
2.2梭梭AQP基因PCR扩增结果
将AQP基因的PCR扩增产物进行1%琼脂糖凝胶电泳,结果表明AQP基因所在泳道出现目的条带,大小与预期的481 bp长度相符(图2).

图1 柴达木盆地梭梭总RNA电泳图Fig.1 Total RNA of Haoxylon ammodendron

图2 AQP基因PCR产物电泳图Fig.2 Amplified results of AQP gene
2.3AQP基因核苷酸序列和氨基酸序列分析
2.3.1核苷酸序列分析AQP基因核苷酸序列经DNASTAR分析(表1),结果表明AQP基因核苷酸序列长度为481 bp,T的含量较高,达到29.52%,基因序列中A+T的含量高于G+C的含量.
2.3.2氨基酸序列分析推导的AQP基因氨基酸序列经DNASTAR软件分析后表明(表2),氨基酸序列长度为160 aa,分子质量为17 026.84 U.各类氨基酸中碱性和酸性氨基酸的含量分别为7.5%和3.12%,疏水性和亲水性氨基酸的含量分别为45%和21.25%.

表1 AQP基因核苷酸序列分析

表2 AQP氨基酸序列分析
2.3.3柴达木盆地梭梭AQP基因与参考基因核苷酸序列和氨基酸序列比较结果将获得的柴达木盆地梭梭AQP基因与Genebank中登录的拟南芥(Arabidopsisthaliana,NM100044.4)、甜菜(Betavulgarissubsp.vulgaris,NM001303070.1)、二穗短柄草(Brachypodiumdistachyon,XM003563129.2)、甘草(Glycyrrhizauralensis,AY781788.1)、荷花(Nelumbonucifera,XM010257025.1)、烟草(Nicotianatabacum,BK008511.1)、桃(Prunuspersica,AF367460.1)、大果栎(Quercusmacrocarpa,FJ495157.1)、Quercusmacrocarpa(FJ495159.1)、番茄(Solanumlycopersicum,AB845607.1)、龙葵(Solanumnigrum,GU575314.1)、独脚金(Strigaasiatica,DQ445128.1)等植物进行核苷酸与氨基酸序列比较,结果表明(图3-4)所得梭梭AQP基因序列与非梭梭属植物AQP基因序列间存在着丰富的变异,其核苷酸序列的同源性在71.7%~85%,氨基酸序列同源性在75%~91.9%,可见柴达木盆地梭梭AQP基因与其他科属植物的AQP基因存在这丰富的变异.

图3 AQP基因核苷酸序列同源性Fig.3 The homology of AQP gene

图4 AQP基因氨基酸序列同源百分比Fig.4 The percentage homology of AQP
2.4AQP蛋白结构预测
Predictprotein预测二级结构结果显示梭梭AQP蛋白二级结构主要为α螺旋占42.5%,β折叠占8.75%,无规则卷曲占48.75%,可见α螺旋和无规则卷曲是梭梭AQP蛋白二级结构的主要构成元件.用TMHMM预测梭梭AQP跨膜区,结果显示AQP含三个跨膜螺旋的结构(图5).

图5 梭梭AQP蛋白跨膜结构域Fig.5 Transmembrane structure prediction of AQP protein
Predictprotein对AQP进行功能位点预测,结果表明,AQP具有1个 N-糖基化位点,即135-138位的NKSK,其糖基化的连接点为天冬酰胺;2个蛋白激酶C磷酸化位点,分别为119-121位的TLR,174-176位的TTK;4个酪蛋白激酶Ⅱ磷酸化位点,分别为25-28位的TLSD,70-73位的SLLD,112-115位的SLGE,160-163位的SEFD; 2个N-端豆蔻酰化位点,分别为16-21位的GVAASR,154-159位的GVYKAM;1个微体细胞C端靶信号,175-177位的TKM;白细胞介素10家族信号,76-96位的KGYLGCQALSEMIQFYLEEVM.
Kyte-Doolittle方法预测结果显示(图6),AQP的亲水性较弱,分布区域较少,疏水性很强,具有多个连续疏水区.在蛋白质二级结构的基础上,采用I-Tasser在线软件对柴达木盆地梭梭AQP进行三维结构预测(图7).预测结果显示:AQP三维结构主要由α螺旋、无规则卷曲和少量β折叠盘绕而成.模型的C-score达到了1.41,说明所建模型可信度较高.
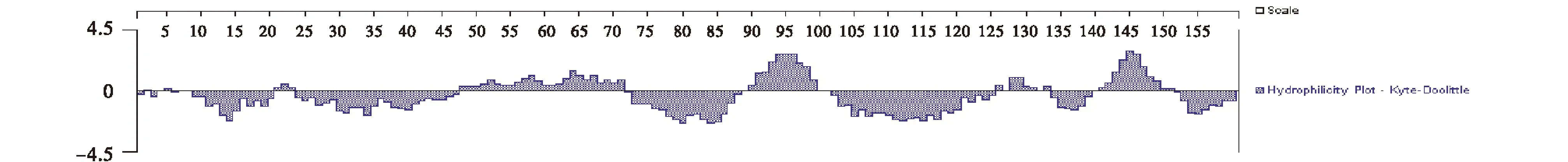
图6 梭梭AQP亲水性分析Fig.6 Hydrophilicity plot of AQP protein

图7 梭梭AQP三维结构的预测Fig.7 The predicted 3D structure of AQP protein
3讨论与结论
在水通道蛋白被发现之前,水分的跨膜运输被认为是通过简单扩散实现的[13].1988年Denker等第一次从人类红细胞膜上纯化分离水通道蛋白[14],1991年Preston等完成了第一个水通道蛋白的克隆、氨基酸序列与蛋白结构的研究,推测其可能是与水分运输相关的蛋白[15],1992年进一步对其功能进行验证表明其在水分运输中发挥作用着重要的功能[16],自此证实水分的跨膜运输是通过水通道蛋白实现的.
目前已证实水通道蛋白广泛存在于动物、植物、酵母、细菌及各种有机体中.植物水通道蛋白在植物应对低温、干旱、盐碱等逆境胁迫时通过调控水分运输从而维持胞内渗透压而保证了植物在逆境胁迫下的生命活动[17-20].由此可见植物能够通过调节AQP的活性,从而使植物在逆境胁迫下保持细胞内水分,在植物适应极端环境中AQP起着重要的作用.
本文通过柴达木盆地梭梭AQP基因的扩增,并对AQP基因的序列进行分析,对其蛋白结构进行预测,旨在为柴达木盆地梭梭AQP的功能研究奠定基础,并为梭梭抗旱、耐盐机制的阐明奠定基础,为转基因培育抗逆优质植物品种积累基因资源.所得柴达木盆地梭梭AQP基因长度481 bp,编码160个氨基酸.另外,将获得的柴达木盆地梭梭AQP基因序列与NCBI登录的其他植物进行同源性比对分析,表明梭梭AQP基因核苷酸序列与不同科属植物AQP基因的同源性在71.7%~85%,氨基酸序列同源性在75%~91.9%,可见不同植物AQP基因存在着核苷酸序列和氨基酸序列上丰富的变异,在保证蛋白必须结构及相应功能位点的稳定基础上又有非功能位点丰富的变异性,是不同植物适应环境的表现.
在蛋白质二级结构中,由于α螺旋含有氢键不易形变,且其一侧由亲水性氨基酸组成,而另一侧由疏水性氨基酸组成,因此使蛋白质具有两亲的性质,即α-螺旋的亲水侧形成空桶状结构进行水分的跨膜运输,而疏水的一侧则通过疏水键与磷脂双分子层作用而将蛋白固定在膜上,从而构成了稳定的跨膜结构域以便进行水分的跨膜运输.本研究中梭梭AQP具有大量的α螺旋,为梭梭AQP二级结构的主要构成元件,符合AQP作为水分转运蛋白的功能特点.此外AQP含有大量的无规则卷曲,而它们往往是构成酶活性部位和其他蛋白质特异的功能部位,在蛋白功能执行中起着重要的作用.此外,柴达木盆地梭梭AQP的疏水性较强,而亲水性较弱,具有多个连续疏水区,疏水区可形成跨膜结构域,与膜上的磷脂双分子层紧密结合,从而形成跨膜通道进行水分转运,跨膜区预测显示AQP具有3个跨膜螺旋的结构,为跨膜蛋白,符合跨膜蛋白疏水性较强的特征.
参考文献
[1]Huang Z Y,Zhang X S,Zheng G H,et al.Influence of light,temperature,salinity and storage on seed germination ofHaloxylonammodendron[J].Journal of Arid Environments,2003,55(3):453-464
[2]赵可夫,周三,范海.中国盐生植物种类补遗.植物学通报,2002,19(5):611-613
[3]史胜青,张守攻,肖文发,等.梭梭小G蛋白基因HaRAN1克隆与表达分析.西北植物学报,2009,29(10):1939-1945
[4]Shi S Q,Shi Z,Qi LW,et al.Molecular responses and expression analysis of genes in a xerophytic desert shrubHaloxylonammodendron(Chenopodiaceae) to environmental stresses[J].African Journal of Biotechnology,2009,8(12):2667-2676
[5]Jiang X C,Guo X H,Pan X L,et al.Construction and differential screening of a cDNA library specific to osmotic stress ofHaloxylonammodendronseedlings[J].Journal of Biochemistry and Molecular Biology,2004,37:527-532
[6]陈鹏,潘晓玲.干旱和NaCl胁迫下梭梭幼苗中甜菜碱含量和甜菜碱醛脱氢酶活性的变化.植物生理学通讯,2001,37(6):520-522
[7]郭春秀,李德禄,刘虎俊,等.不同种源梭梭苗木耐盐性盆栽试验[J].甘肃农业大学学报,2008,43(6):110-112
[8]石磊,甘晓燕,陈虞超,等.梭梭甜菜碱醛脱氢酶基因克隆及序列分析[J].西北植物学报,2010,30(2):223-228
[9]甘晓燕,石磊,周晓燕,等梭梭过氧还蛋白基因(PrxQ)克隆与序列分析[J].西北农业学报,2012,21(6):53-57
[10]周春娥,耿飒,齐力旺.梭梭胆碱单氧化物酶基因(CMO)的cDNA克隆[J].安徽农业科学,2007,35(9):2558-2559
[11]杨丽丽,吴明朝,甘晓燕,等.梭梭VHA-B基因克隆及序列分析[J].西北农业学报,2013,22(10):68-74
[12]甘晓燕,石磊,陈虞超,等.梭梭Na+/H+逆向转运蛋白基因克隆及分析[J].西北植物学报,2012,32(2):225-230
[13]Agre P,Bonhivers M,Borgnia M-J.The aquaporins,blueprints for cellular plumbing systems[J].The Journal of Biological Chemistry,1998,273(24):14659-14662
[14]Denker B M,Smith B L,Kuhajda F P,et al.Identification,purification,and partial characterization of a novel Mr28,000 integral membrane-protein from erythrocytes and renal tubules[J].Journal of Biological Chemistry,1988,263(30):15634-16542
[15]Preston G M,Agre P.Isolation of the cDNA for erythrocyte integral membrane protein of 28 kilodaltons:member of an ancient channel family[J].Proceeding of the NationalAcademy of Sciences of the United States of America,1991,88(24):11110-11114
[16]Preston G M,Carroll T P,Guggino W B,et al.Appearance of water channels inXenopusoocytesexpressing red cell CHIP28 protein[J].Science,1992,256(5055):385-387
[17]Srivastava A K,Penna S,Nguyen D V,et al.Multifaceted roles of aquaporins as molecular conduits in plant responses to abiotic stresses[J].CriticalReviews in Biotechnology,2014,28:1-10
[18]Li G,Santoni V,Maurel C.Plant aquaporins:roles in plant physiology[J].Biochimica et Biophysica Acta,2014,1840(5):1574-1582
[19]Huang C,Zhou S,Hu W,et al.The wheat aquaporin gene TaAQP7 confers tolerance to cold stress in transgenic tobacco[J].Journal of Biosciences,2014,69 (3-4):142-148
[21]Xu Y,Hu W,Liu J,et al.A banana aquaporin gene,MaPIP1;1,is involved in tolerance to drought and salt stresses[J].BMC Plant Biology,2014,14(1):59
(责任编辑李辛)
Gene clone and protein prediction ofAQPgene inHaloxylonammodendronin Qaidam basin
MA Yu-hua,YE Gui-sheng,MA Xiu-fang,LI Yu-ying,LONG Xiao-chen,MA Juan,YE Gui-lian
(College of Agriculture and Animal Husbandry,Qinghai University,Xining 810016,China)
Abstract:【Objective】 To clone the AQP gene and protein prediction of Haloxylon ammodendron .【Method】 The fragment of AQP gene of H.ammodendron in Qaidam basin was amplified by the specific primers and sequenced,then the nucleotide sequence and amino acid sequence of AQP gene was identified and summarized in this paper.【Result】 The sequence analysis indicated that the total length of AQP gene was 481 bp,which coding 160 AA.Predictions of hydrophilicity,secondary structure,transbilayer helix,tertiary structure showed that α-helix and random coil were the main secondary structure of AQP,AQP had 1 N-glycosylation sites,2 protein kinase C-phosphorylation site,4 casein kinase II phosphorylation sites,2 N-myristoylation sitess,1 microbodies C-terminal targeting signal,1 interleukin-10 family signature.AQP had a weaker hydrophilicity,and it had 3 transmembrane structures.The tertiary structure prediction showed the main tertiary structure of AQP include α-helix and random coil,and a small amount of β fold.【Conclusion】 This result will lay foundation for the further study of the structure of AQP gene and it's function of H.ammodendron.
Key words:Haloxylon ammodendron;AQP gene;amplification;sequence analysis
基金项目:青海省科技厅自然科学基金项目(2012-Z-938Q);“123高层次人才培养工程”项目.
收稿日期:2015-03-06;修回日期:2015-06-23
中图分类号:Q 78
文献标志码:A
文章编号:1003-4315(2016)02-0093-05
第一作者:马玉花(1978-),女,副教授,博士,主要从事森林培育理论与技术、植物资源开发利用方面的研究.E-mail:sophere8@163.com
猜你喜欢
哈哈画报(2022年4期)2022-04-19 11:11:54
科学大众(2020年17期)2020-10-27 02:48:48
农民致富之友(2020年12期)2020-05-11 05:55:03
中学生天地(B版)(2019年4期)2019-05-08 03:22:54
江苏农业科学(2017年1期)2017-02-27 10:30:11
山东农业科学(2016年11期)2016-12-17 21:00:42
江苏农业科学(2016年4期)2016-06-14 02:05:28
江苏农业科学(2015年9期)2015-10-20 17:02:59
江苏农业科学(2015年8期)2015-09-10 21:03:17
绿色中国(2015年6期)2015-07-03 01:13:10
