
铅处理对菜用大黄种子萌发和幼苗生长的影响
2016-06-14 15:49姜立娜邵珠田宋子文蔡祖国
江苏农业科学 2016年4期
姜立娜+邵珠田+宋子文+蔡祖国


摘要:以菜用大黄为试验材料,研究不同浓度的铅溶液(0、50、150、300、600、1 200 mg/L)对菜用大黄种子萌发和幼苗生长的影响。结果表明,Pb2+浓度为0~300 mg/L时对菜用大黄种子发芽指标影响不大,随着Pb2+浓度的升高出现一定的抑制作用,表明菜用大黄种子萌发对铅有一定耐性。幼苗形态指标在Pb2+浓度为0~50 mg/L时最高,当浓度大于50 mg/L时各项指标均表现为下降趋势;幼苗可溶性蛋白含量、过氧化氢酶(CAT)活性随着Pb2+浓度的升高呈现波动变化的趋势,丙二醛(MDA)含量整体呈现升高的趋势。
关键词:铅;菜用大黄;种子萌发;幼苗生长
中图分类号: S649.01
文献标志码: A
文章编号:1002-1302(2016)04-0223-03
近年来,随着中国工业的快速发展,环境中重金属污染加重,特别是铅(Pb)含量明显增加。铅不是植物生长发育的必需元素,但是植物可以通过根系吸收土壤中的铅。研究发现,高浓度铅会改变植物细胞膜透性,影响植物光合作用、呼吸作用和多种代谢过程,进而对植物造成伤害,并且会进一步通过食物链进入人体,危及人类健康[1-3]。
菜用大黄(Rheum rhaponticum L.)为蓼科大黄属多年生草本植物,以叶柄为食用器官,其产量高、便于管理,1次定植可连续收获4~6年[4-6]。由于其营养价值高、保健功能强、风味独特被列入奥运蔬菜,开发前景广阔。目前,对菜用大黄的研究主要集中在组培苗不定芽增殖能力、生根特性和气孔特性、核型分析等方面[7-9],关于重金属铅对菜用大黄种子及幼苗的影响还未见报道。本试验研究了铅处理对菜用大黄种子萌发和幼苗生长的影响,以此衡量菜用大黄对重金属铅的耐受能力,以期为预防菜用大黄早期铅伤害以及筛选重金属铅的耐性植物提供理论依据。
1 材料与方法
1.1 试验材料
菜用大黄种子由河南科技学院园艺园林学院菜用大黄引种课题组提供。经测定试验所用菜用大黄种子净度为9381%,千粒质量为13.5 g。
1.2 试验方法
将菜用大黄种子置于50~55 ℃的温水中消毒15 min,其间不断搅拌,将温度降至25 ℃左右。用蒸馏水冲洗 5~6 次,吸水纸吸去种子表面的水分。分别用不同浓度(0、50、150、300、600、1 200 mg/L)的硝酸铅溶液(以铅的浓度计算)进行浸种处理,每个处理用种60粒,试验进行3 次重复。浸种时间为24 h,种子浸泡后分别放在垫有2层滤纸并用各种浓度处理液浸湿的各个培养皿中,每个培养皿30粒。将培养皿置于(22±1) ℃恒温人工气候箱中,待种子长出2张子叶后,打开光照将强度设定为4 000 lx,每天添加处理液以保持种子湿润状态。于3 d后计算发芽势,7 d后计算发芽率,14 d后测定形态指标,同时测定可溶性蛋白含量、过氧化氢酶(CAT)活性、丙二醛(MDA)含量等生理指标。
1.3 测定指标与方法
1.3.1 发芽指标[10]
发芽势=3d后发芽种子数/供试种子总数×100%;
发芽率=7d后发芽种子数/供试种子总数×100%。
1.3.2 幼苗形态指标 在试验14 d后,测定幼苗鲜质量、根鲜质量、根长、株高。
1.3.3 幼苗生理指标 可溶性蛋白含量的测定采用考马斯亮蓝G250法[10];CAT活性的测定采用紫外吸收法[11];丙二醛(MDA)含量测定采用硫代巴比妥酸法[12]。
2 结果与分析
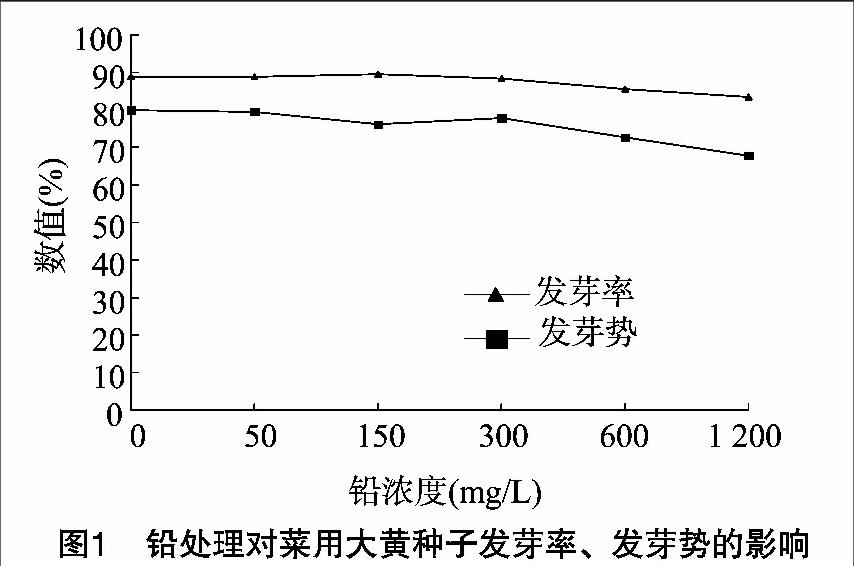
2.1 铅处理对种子发芽势和发芽率的影响
从图1可以看出,随着Pb2+浓度的增加,菜用大黄种子的发芽率、发芽势整体呈现下降的趋势。Pb2+浓度低于 300 mg/L 时,对菜用大黄种子的发芽率、发芽势影响不大;当Pb2+浓度高于于300 mg/L时,种子发芽率、发芽势与对照相比均表现为下降;Pb2+浓度为1 200 mg/L时,种子的发芽率达到最低值,比对照低5%左右,种子的发芽势比对照低13%,但是仍能达到67.78%。结果表明,Pb2+浓度较低时对菜用大黄种子的发芽率、发芽势影响不大;当Pb2+浓度高于 300 mg/L 时对发芽势表现出了一定的抑制作用。
2.2 铅处理对菜用大黄幼苗形态指标的影响
从表1可以看出,菜用大黄的根长、株高在Pb2+浓度为50 mg/L时,与对照相比差异不显著;随着Pb2+浓度的增加,当Pb2+大于50 mg/L时,根长、株高开始下降,与对照差异显著,表明高浓度Pb2+处理对菜用大黄的根长、株高具有明显抑制作用。幼苗鲜质量、根鲜质量随着Pb2+浓度的增加呈现先增加后减少的趋势,当Pb2+浓度大于150 mg/L时,与对照差异明显。当Pb2+浓度为1 200 mg/L时幼苗几乎停止生长,无法进行相应生理指标的测定。结果表明,低浓度的Pb2+处理对菜用大黄的鲜质量有一定的促进作用,高浓度则出现抑制;对根长、株高的影响则是表现为抑制作用,浓度越高,抑制作用越显著。
2.3 铅处理对菜用大黄幼苗生理指标的影响
2.3.1 铅处理对幼苗可溶性蛋白含量的影响 从图2可以看出,菜用大黄幼苗可溶性蛋白的含量随着Pb2+浓度的升高,整体呈现下降的趋势,在150 mg/L时出现反弹,比相邻浓度高,但仍比对照组含量低。当Pb2+浓度超过150 mg/L后,可溶性蛋白含量逐渐降低,表现出铅处理对菜用大黄幼苗的抑制作用。
2.3.2 铅处理对幼苗CAT活性的影响 从图3可以看出,不同浓度的Pb2+处理后,菜用大黄幼苗的CAT活性呈现先升高后降低的趋势,在600 mg/L时出现反弹。表现为低浓度Pb2+范围(<50 mg/L) 内随着Pb2+浓度的增加,CAT的活性增强,当浓度为50 mg/L时活性高于CK并达到最高值;在高浓度Pb2+(150~600 mg/L)作用下,CAT活性下降且明显低于对照,在浓度为300 mg/L时达到最低值。结果表明,低浓度的Pb2+对CAT活性具有激活作用,随着Pb2+浓度的逐渐增大,又转为抑制作用,并对CAT酶系统产生伤害。
2.3.3 铅处理对幼苗MDA含量的影响 从图4可以看出,随着Pb2+浓度的增加,MDA含量整体呈现增加的趋势。当Pb2+浓度在50~150 mg/L时,MDA含量与CK相比变化不大;当Pb2+浓度为300、600 mg/L时,MDA含量与对照相比有明显的升高,分别为对照的185.30%、192.80%。
3 结论与讨论
试验结果表明,随着Pb2+浓度的提高,菜用大黄种子的发芽率、 发芽势、 发芽指数和活力指数均逐渐降低,与Pb胁迫条件下牛蒡、狭叶香蒲、黄瓜种子萌发特性的研究结论[13-15]一致。本研究中,在Pb2+处理下,菜用大黄种子的发芽率、发芽势均低于对照,但其发芽率仍然达到88.33%,与对照88.89%差异不显著。对菜用大黄幼苗生理指标的测定结果表明,在较低浓度(<50 mg/L) Pb2+处理下,幼苗的株高、鲜质量高于对照,在高浓度下开始下降;而根长、根鲜质量在一定范围内随着Pb2+浓度的提高而下降,表明幼苗根对重金属Pb的反应比芽更为敏感,本结论与戴敏等相关研究结果[13,16]类似。尽管高浓度Pb对菜用大黄种子萌发和幼苗生长有一定的抑制效应,但Pb2+浓度在1 200 mg/L时,菜用大黄种子仍具有较高的萌发率,仅发芽势有所降低,萌发速率减缓,前期幼苗仍可以生长,但生长非常缓慢,后期开始出现死亡,表明菜用大黄种子萌发和幼苗生长都对Pb均具有一定耐性。
Pb是一种对植物有积累性危害的重金属,过量的Pb进入植物体内,会破坏生物膜的结构,导致叶绿体、线粒体及细胞核等重要器官的损伤,从而影响各种代谢过程,最终导致植物生长减缓甚至死亡[1]。 可溶性蛋白含量的变化反映了植物生长的状态和新陈代谢的水平。本研究中不同浓度的Pb2+处理后,菜用大黄幼苗的可溶性蛋白含量均低于对照,且浓度越高,含量越低,表明铅处理影响了菜用大黄幼苗的生理代谢。不同浓度的Pb2+处理对菜用大黄幼苗的CAT活性的影响不同,低浓度Pb2+对菜用大黄幼苗的CAT活性有激活作用,高浓度的Pb2+对菜用大黄幼苗的CAT活性有抑制作用,表明菜用大黄幼苗对Pb2+浓度有一定的适应性,当Pb2+浓度超过菜用大黄种子的耐受极限时将出现抑制作用,与铅胁迫对甜瓜种子胚芽酶系统影响的结论[17]一致。
植物在逆境或衰老条件下会发生膜脂过氧化,MDA是一种重要的膜脂过氧化产物,膜脂过氧化程度可以通过植物组织内MDA的含量来反映。本试验中Pb2+浓度在0~150 mg/L时,菜用大黄幼苗中MDA含量变化不大,表明较低浓度Pb胁迫对植株伤害并不明显;但是随着Pb2+浓度的进一步增加Pb2+浓度﹥300 mg/L时),MDA含量则急剧增加,表明高浓度的铅处理造成菜用大黄膜脂过氧化加剧,膜伤害程度增加。
综上所述,菜用大黄的种子萌发和幼苗生长对铅有一定的耐受性,但随着处理浓度的升高,幼苗生长受到明显抑制。本研究结果为后期进行菜用大黄耐铅品种选育以及菜用大黄耐铅机制研究提供了相关理论依据。
参考文献:
[1]江行玉,赵可夫. 植物重金属伤害及其抗性机理[J]. 应用与环境生物学报,2001,7(1):92-99.
[2]李 斌,李志辉,吴际友,等. 铅胁迫对4种行道树种幼苗叶片丙二醛含量的影响[J]. 湖南林业科技,2010,37(2):8-11.
[3]虎 瑞,苏 雪,晏民生,等. 重金属Pb(Ⅱ)对3种藜科植物种子萌发的影响[J]. 植物研究,2009,29(3):362-367.
[4]Zhao Y P,Grout B,Crisp P. Unexcepted susceptibility of breeding lines of European rhubarb(Rheum rhaponticum L.) to leaf and petiole spot disease[J]. Acta Hort,2004,637:139-144.
[5]王健梅. 奥运蔬菜系列——叶菜类[J]. 中国农业信息,2006(2):8.
[6]卢 莉,赵一鹏. 菜用大黄的研究进展[J]. 广东农业科学,2008(2):19-21,27.
[7]蔡祖国,周 岩,卢 莉,等. 不同激素对菜用大黄茎尖离体再生的影响研究[J]. 广东农业科学,2010,37(3):132-134,144.
[8]任文娟,郭小菲,姜立娜,等. 菜用大黄染色体制片优化及核型分析[J]. 华北农学报,2013,28(5):128-132.
[9]赵一鹏,Grout B W,周 岩. 欧洲大黄茎尖组织培养与快速繁殖[J]. 河南职业技术师范学院学报,2004,32(3):24-25,28.
[10]张志良,瞿伟菁,李小方. 植物生理学实验指导[M]. 4版.北京:高等教育出版社,2009:58-60,159-160.
[11]李合生,孙 辉,赵世杰,等. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:134-137.
[12]高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006:210-211.
[13]戴 敏,孙 坤,苏 雪,等. 铅胁迫对牛蒡种子萌发及幼苗生长的影响[J]. 北方园艺,2014(4):132-136.
[14]徐金波,徐迎春,赵 慧,等. Pb 胁迫条件下狭叶香蒲种子的萌发特性及其幼苗的生理响应[J]. 植物资源与环境学报,2014,23(3):74-80.
[15]陈新红,叶玉秀,庞闰瑾. 镉、铅对黄瓜种子发芽及幼苗生长的影响[J]. 中国蔬菜,2009(8):18-22.
[16]符卓旺,彭 娟,朱 洁,等. 铅对紫花苜蓿种子萌发与幼苗生长的影响[J]. 中国农学通报,2011,27(3):303-306.
[17]赵惠新,覃建兵,祝长青,等. 铅胁迫对甜瓜种子胚芽酶系统及MDA含量的影响[J]. 种子,2008,27(12):82-84.滕红梅,昝亚玲,贾 鹏,等. 不同产地黄芪药材中毛蕊异黄酮葡萄糖苷含量的比较[J]. 江苏农业科学,2016,44(4):226-228.
猜你喜欢
现代农业科技(2016年20期)2016-12-20
现代园艺(2016年17期)2016-10-17
