
日本乙型脑炎病毒对猪外周血单核细胞炎性因子mRNA转录时相影响的研究
2016-06-07 09:27何潇湘景亚星靳晓慧兰培英张利卫魏战勇
河南农业大学学报 2016年1期
何潇湘,景亚星,靳晓慧,兰培英,张利卫,周 雍,魏战勇,
猪日本乙型脑炎(Porcine Japanese Encephalitis B)是由日本乙型脑炎病毒(Japanese Encephalitis Virus,JEV)引起的,主要依靠吸血的雌蚊为传播媒介而进行传播的一种严重侵害中枢神经系统的急性人兽共患的传染病。JEV的危害较大,可感染很多动物和人,对猪的危害最为严重[1-2]。该病毒可引起母猪发生毒血症、子宫内膜炎;怀孕母猪发生流产、死胎、畸形胎、木乃伊胎等繁殖障碍[3];公猪发生单侧性睾丸炎,严重影响着养猪业的发展[4-5]。有关JEV感染引起猪的相关疾病的发病机制和免疫病理学研究尚不深入,这严重制约了新型疫苗、有效药物的研发和疾病防控。猪外周血单核细胞(Porcine Peripheral Blood Mononuclear Cell,PBMC)是机体重要的免疫细胞,能分泌白介素、干扰素、集落刺激因子等细胞因子,在免疫应答过程中,细胞因子对细胞间的相互作用、细胞的生长和分化具有重要的作用,研究细胞因子的转录对研究并揭示猪炎症反应和免疫应答的激发与调控有着重要作用[6]。罗线连等[7]研究发现,JEV 感染猪PBMC 后可引起 MHC-Ⅰ、Mx1、2’,5’寡腺苷酸合成酶(2-5OAS)、转录因子IRF3等抗病毒相关因子增加,为研究细胞抗病毒机制奠定基础。但对于JEV促炎性反应,特别是对于促炎因子的研究较少。本试验利用相对荧光定量PCR方法,分析不同时间点,JEV感染PBMC后的促炎性细胞因子(IL-8、IL-12、IL-17、IL-18)mRNA 的表达,为探讨JEV免疫致病机制提供依据。
1 材料与方法
1.1 病毒和细胞
猪日本乙脑病毒标准毒株购自中国兽药监察所,经河南省动物性食品安全重点实验室传代保存,病毒在PK-15细胞上传代10代,测定病毒滴度为104.5TCID50·mL-1。选择 60 日龄,体重为 50 kg左右的长白猪(由河南省正阳种猪场提供),经PCR技术或ELISA检测猪的常见传染病,如猪伪狂犬病毒(PRV)、猪细小病毒(PPV)、猪繁殖与呼吸综合症病毒(PRRSV)、猪圆环病毒2型(PCV-2)等均为阴性。无菌采集猪抗凝的外周血,参照文献[3]的方法分离获得猪PMBC,并将其培养于含7%FBS的RPMI-1640培养基中,5%CO237℃培养。
1.2 主要仪器和试剂
MX3005 Real-Time PCR仪器(吉泰科技公司生产);紫外凝胶成像系统(美国SIM公司生产);台式高速离心机(Sigma公司生产);Revert AidTMFirst Strand cDNA Synthesis Kit I(MBI Fermentas公司);胎牛血清培养液(Hyclone公司);E.Z.N.A Total RNA KitⅠ试剂(Omega公司);SYBR Premix Ex Taq试剂(TaKaRa公司);RPMI-1640培养液(Gibco公司);其他试剂均为分析纯。
1.3 病毒感染
取2 mL的PMBC细胞(细胞浓度为105个·mL-1)置于6孔板每孔中;采取同步接种方法接种JEV病毒(感染复数MOI为1),每个处理设2个重复孔,轻轻摇匀,放入37℃ 5%的CO2恒温培养箱中培养。
1.4 细胞样品收集
将 JEV 感染后 1,3,12,24,48,72 h 的单核细胞用2 000 r·min-1离心收集 PMBC,并用 PBS重悬、洗涤2次,并离心收集细胞沉淀,然后直接提取RNA或者保存于-20℃备用。
1.5 RNA的提取和反转录
JEV的RNA和细胞总RNA提取参照E.Z.N.A Total RNA Kit I说明书进行,具体方法和步骤参照文献[8-9]进行,提取的总RNA为模板分别进行JEV反转录和炎性细胞因子的反转录,反转录合成的cDNA作为RT-PCR的模板。
1.6 引物和相对定量PCR
引物的设计,参照GenBank公布的JEV和各炎性因子的基因序列,使用Primer Premier 5.0软件进行引物设计,设计后使用 GenBank的 Blast Primer对引物进行筛选,RT-PCR引物及其反应条件见表1。荧光定量PCR反应体系为20 μL,其中SYBR Premix Ex Taq 10 μL,上游、下游引物各 0.4 μL(浓度为 15 μmol·L-1),cDNA 400 ng,用灭菌双蒸水添加至20 μL;反应条件为:95℃ 1 min;95℃ 10 s。表1中所指示退火温度15 s,72℃ 30 s,40个循环;循环结束后升温至95℃ 30 s,再降至55℃,开始以0.2℃·s-1递增至95℃,采集荧光信号得出扩增产物的溶解曲线,并于20℃结束反应。
1.7 数据分析
Ct值在荧光定量PCR中,主要反映模板中目标基因含量,通过与β-action为看家基因比较,以采用2-△△Ct法[10]进行分析,能准确反映出目标基因的相对含量。试验中所得数据应用SPSS软件进行统计学分析,p<0.05时表示差异显著。
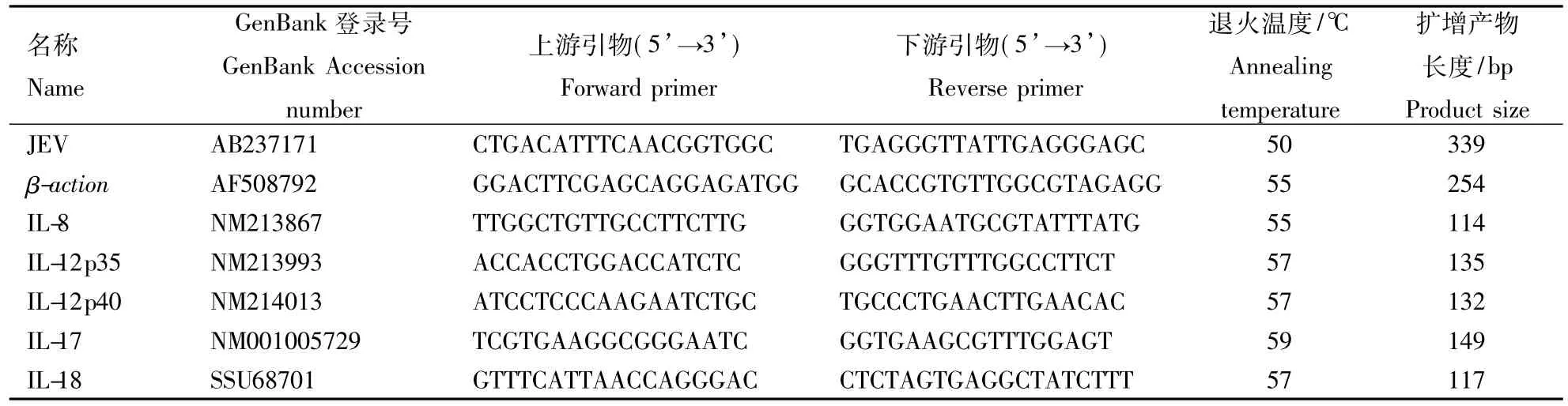
表1 Real-time PCR引物及其反应条件Table 1 Primers and conditions used for Real-time PCR assays
2 结果与分析
2.1 病毒RNA水平的测定
PBMC 在感染 JEV 后,分别在 1、3、12、24、48、72 h收集细胞,离心收集细胞沉淀,并提取RNA,之后反转录为cDNA,以用于RT-PCR。与看家基因β-action进行比较分析后,将病毒感染1 h时病毒RNA含量定义为1倍,经计算得到不同时间病毒RNA在细胞的增殖倍数,结果见图1。

图1 感染后不同时间PBMCs中JEV RNA相对含量的变化Fig.1 Changes of relative content of JEV DNA in PBMCs after infection
由图1可知,JEV感染PMBC后,1 h就可以检测到病毒RNA,随着感染时间延长,病毒RNA含量逐渐上升,在感染后24 h病毒含量大量增加,在感染后72 h达到最大值,约为1 h时的10倍左右。
2.2 炎性细胞因子转录时相
猪 PBMC 感染 JEV 后,在 1、3、12、24、48、72 h时候分别收集细胞,并按试剂盒步骤提取RNA,反转录合成c DNA,经荧光定量PCR,与看家基因βaction进行比较分析,并以JEV 0 h mRNA含量定义为1倍,计算出不同时间段细胞因子mRNA在细胞内的增殖倍数。
IL-8 感染 1、3、24、72 h 的表达量分别是对照组的1.7、10、3.2、2.8 倍,与对照组相比,均差异极显著(P <0.01),12,48 h表达量降低;IL-12p40 感染1、3、12、24 h 后分别为对照组的 1.6、5.7、5.3和1.9倍,与对照组相比,均差异极显著(p<0.01)接着在48 h表达量降低,72 h甚至不表达;IL-12p35感染1 h开始表达,3 h表达降低,12、24 h表达量都增加,48 h表达量达到峰值,约73倍左右,与对照组相比,差异极显著(p<0.01),72 h表达量降低;IL-17感染1、3、12 h后与对照组相比,表达量增加的幅度不明显,差异不显著(P>0.05),24、72 h 甚至不表达,48 h 表达量大量增殖;IL-18感染1 h开始表达,3 h大量表达,12、24、48 h表达量增高趋势,在72 h量达到峰值,约为7.8倍,且与对照组相比,差异极显著(P <0.01)结果见图2。
3 讨论
JEV可以在PK15、Vero和BHK21等细胞中增殖,且能引起较明显的细胞病变。本研究发现JEV在感染PBMC后1 h就能检测到该病毒,感染后3 h开始增殖,72 h达到峰值。这有可能JEV可以感染PBMC,但却不是JEV增殖的理想细胞,因而前期病毒增殖较为缓慢,病毒在后期大量增殖,引起细胞死亡,可能是PBMC作为免疫细胞而发挥抗病毒作用所致[7]。
促炎性细胞因子是指机体受到细菌或者病毒的感染时,细胞内诱导生成的一系列对炎症反应有促进作用的细胞因子,例如白介素、干扰素和肿瘤坏死因子等。而本文所研究的IL-8,IL-12,IL-17和IL-18都是促炎性细胞因子,它们的作用常常交替重叠[22]。
白细胞介素8(IL-8)是一种主要来源于单核巨噬细胞形成的细胞趋化因子。1986年KOWNATZKI首先发现单核细胞可以分泌产生IL-8,该因子在激活嗜中性粒细胞反应中发挥很大作用,对炎症反应和免疫过程有重要的调节作用。有报道CXC家族(α亚家族)趋化因子主要在疾病的急性反应期介导炎性细胞起作用[11],IL-8通过自分泌和旁分泌的作用刺激内膜细胞和其他的腺体细胞,而这些细胞又分泌包括IL-8在内的多种细胞因子,此时若有其他炎症因子的刺激,分泌作用又进一步地加强,从而形成恶性循环,加快疾病的发展。据孙琳[12]的研究推测,IL-8参与疾病的致病过程,可能存在与其他趋化因子参与疾病的慢性化。
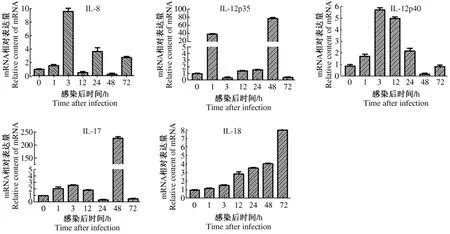
图2 细胞因子的转录时相Fig.2 The transcriptional profiles of cytokines
本研究发现,JEV感染淋巴细胞1 h后IL-8的表达量开始表达,但不太显著;在3 h表达量达到峰值,随后在12 h表达量下降,在24 h后IL-8的表达量有增高趋势,随后在48 h表达量又紧跟着出现下滑趋势;接着在72 h再次出现少量增高。据此可以推测PBMC在受到JEV感染后,前期细胞活性强,病毒没有大量增殖,随着细胞内外环境的改变,IL-8表达量出现忽高忽低的表现,说明细胞和病毒产生对抗反应;如果病毒毒力强,细胞因子IL-8 mRNA就能快速大量表达,以便于参与抗病毒反应,发挥其生物学效应。
白细胞介素12(IL-12)是由T淋巴细胞、B淋巴细胞、巨噬细胞刺激产生的细胞因子。IL-12含有亚基IL-12p35和亚基IL-12p40,这2个亚基通过二硫键的作用有机结合在一起,p35只决定IL-12的种属特异性,p40却在IL-12与受体结合时起关键作用[13]。单独的两个亚基都不能与IL-12受体结合,只有当2个亚基有机结合在一起后才可以与IL-12受体充分结合,这样才能发挥IL-12生物学效应[14]。IL-12是细胞毒性T淋巴细胞(CTL)的始动因子,它能促进Th1淋巴细胞的增殖和分化,在机体组织的抗病毒感染过程中有强大的功能。IL-12和IL-18共同作用可以诱导刺激B细胞产生大量的IFN-γ的mRNA和蛋白,而在抗肿瘤的免疫应答过程中发挥重要作用。所以人们常利用其抗肿瘤的这种生理作用,把IL-12因子应用于慢性感染疾病的治疗,有时也可用于治疗因免疫系统发育不全或遭受损害引起的免疫陷性疾病[15]。尹晓玲等[16]研究发现,IL-12能够促进结肠癌细胞的凋亡,它低水平表达/不表达与结肠癌进展的相关性,证实IL-12对直接限制结肠癌恶性表型的潜在益处。王明琼[17]发现,对乙脑重型和普通型患儿的IL-12水平无显著性差异,但均明显高于轻型,IL-12可作为乙脑实质损伤程度的重要指标和治疗新靶点,这为乙脑的研究拓展了新领域。
通过研究发现,在JEV感染PBMC 1h后IL-12p35和IL-12p40在mRNA水平的表达量均明显增加,随着时间的推移,表达量逐渐增加,IL-12p35在48 h时表达量达到峰值,约为空白细胞的73倍左右,IL-12p40在3 h时表达量达到峰值,为空白细胞的5.7倍之多,随后呈低水平表达,72 h后二者表达量都出下降趋势。根据实验结果推测:IL-12是细胞免疫应答反应的起始因子,IL-12 mRNA的表达和蛋白质的合成可能比其他细胞因子要早,这为病毒感染早期诊断提供指标。在感染72 h后二者表达量都表现出现下降趋势,原因可能是IL-4和IL-10等其他细胞因子在免疫系统的调控下,对IL-12 mRNA的表达产生了抑制作用。
白细胞介素17(IL-17)普遍存在于单核细胞中,是一种多功能的炎症前细胞因子,具有155个氨基酸。它能够参与组织细胞炎症发生发展、转归反应过程;也与免疫应答、免疫排斥等多种生物学活性有关[18]。在炎症的反应中,IL-17与中性粒细胞的生成快慢、分散和聚集有密切关系。IL-17在疾病的发展过程中起着关键的作用,在类风湿关节炎这个疾病中,IL-17是判断其疫病程度的重要指标之一[19]。
试验结果发现,当JEV感染PBMC后1,3和12 h表达量增加缓慢,在24 h时,IL-17的表达量明显下降,在后期48 h才大量表达。IL-17在感染72 h的时候表达量又呈现剧烈下滑趋势,甚至可以说是不表达。这是由于免疫系统高度调控IL-17的表达,在细胞的自身免疫应答阶段中,一般很难检测到IL-17 mRNA的基因表达,又加上JEV感染细胞的过程中大量促炎细胞因子参与抗病毒反应,能大大降低IL-17 mRNA的表达量。
白细胞介素18(IL-18)因其可以诱导γ干扰素(IFN-γ)的产生,因此又叫做 IFN-γ诱导因子[20]。它是一种由活化的单核细胞和巨噬细胞系统的细胞分泌产生的细胞因子,在机体组织和细胞中普遍表达。研究发现,IL-18的结构类似IL-1,但功能却与IL-12相似,它与其他因子产生一定的作用[21]。IL-18能诱导多种细胞因子的产生,如TNF-α,而它又能刺激IL-6的产生,IL-18也能直接促进中性粒细胞产生IL-6。IL-18是促进Th1反应为主的细胞因子,在抵抗病毒感染、免疫调节和抗肿瘤上有很重要的作用。IL-18因子能刺激NK细胞产生抗肿瘤能力,IL-18还可以趋化中性粒细胞,中性粒细胞对病原体的吞噬、杀死、降解作用往往与它也有关。本研究表明,JEV感染外周血细胞后3 h后IL-18表达量开始增多,在感染后72 h,IL-18表达量达到高峰。因此,IL-18参与病毒的杀死过程,在炎症反应整个阶段中都起到很重要作用。
本试验利用荧光定量PCR,测定和分析了JEV感染猪PBMC后病毒及促炎细胞因子的mRNA转录水平的变化情况。结果显示,JEV感染PBMC后随着时间的延长而含量增高,在72 h达到高峰。而IL-8在感染初期的时候含量增高,随后降低,在72 h时候又再次升高;IL-12p35在48 h达到高峰,IL-12p40在感染后3 h就达到高峰;IL-17在48 h达到高峰之后,随后开始下降;IL-18从开始便逐渐上升,在感染后期达到稳定。由此推测,JEV感染促进了PBMC的发生炎症,促进细胞释放病毒,影响其他促炎性细胞因子的释放,继而影响特异性免疫功能的激活与正常发挥,为 JEV病程的持续发展创造了前提条件。
[1] SOLOMON T,NI H,BEASLEY D W,et al.Origin and evolution of Japanese encephalitis virus in Southeast Asia[J].J Virol,2003,77(5):3091 -3098.
[2] GHOSH D,BASU A.Japanese encephalitis a pathological and clinical perspective[J].PLOS Negl Trop Dis,2009,3(9):e437.
[3] NGA P T,CARMEN P,CUONG V D,et al.Shift in Japanese encephalitis virus(JEV)genotype circulating in Northern Vietnarn:implications for frequent introductions of JEV from Southeast Asia to East Asia[J].J Gen Virol,2004,85(6):1625 -1631.
[4] 汤德元,郭万柱.日本脑炎病毒及其疫苗的研究[J].中国兽医学报,2005,25(2):217-221.
[5] 张杰.猪乙型脑炎病的诊断与治疗[J].猪场兽医,2014,305(9):88.
[6] TATIANA G,VALERIO L,LIUDMILA S,et al.αvβ3-integrin is a major sensor and activator of innate immunity to herpes simplex virus-1[J].Proc Natl Acad Sci USA,2012,109(48):19792-19797.
[7] 罗线连,王瑞宁,李金磊,等.日本乙型脑炎病毒对猪的外周血单核细胞抗病毒因子mRNA表达的影响[J].中国人兽共患病学报,2012,28(2):116-119.
[8] 萨姆布鲁克 J,拉塞尔 D W .分子克隆实验指南[M].黄培堂,译.北京:科学出版社,2001:463-471.
[9] 李厚伟,魏战勇,尹海燕,等.猪细小病毒感染PK-15细胞抗病毒相关因子转录变化的分析[J].畜牧兽医学报,2011,42(1):48-55.
[10]郭东辉,张莉娟,李金磊,等.PCV2与PPV共感染猪外周血单核细胞对其细胞凋亡相关因子表达水平的影响[J].畜牧兽医学报,2013,44(1):71-77.
[11] ULUKUS M,ULUKUS E C,TAVMERGEN E N,et al.Expression of interleukin-8 and monocyte chemotactic protein 1 in women with endometriosis[J].Fertil Steril,2009,91(3):687-693.
[12]孙琳,王建.白细胞介素-8在子宫内膜异位症中的作用[J].现代预防医学,2013,40(9):1643-1637.
[13]张立,王迎新,杨婷,等.白介素12与常见自身免疫性疾病的研究进展[J].中华疾病控制杂志,2014,18(11):1099-1103.
[14] ROMERO J F,IBRAHIM G H,RENGGLI J ,et al.IL-12p40-independent induction of protective immunity upon multiple Plasmodium berghei irradiated sporozoite immunizations[J].Parasite Immunology,2007,29(11):541-548.
[15]陈鸿珊,张兴权.抗病毒药物及其研究方法[M].北京:化学工业出版社,2006:253-268.
[16]尹晓玲,陈应果,张建红,等.人IL-12对结肠癌肝细胞生物学特性的影响及其机制[J].第三军医大学学报,2013,35(12),1215-1218.
[17]王明琼.流行性乙型脑炎患儿ET-1和IL-12检测的临床意义[J].中国现代医学,2010,48(2):40-41.
[18] KOLIS J K,LINDEN A.Interleukin-17 family members and inflammation[J].Immunity,2004,21(4):467 -476.
[19]戴小波,孙万邦.IL-17免疫调节作用的研究进展[J].检验医学与临床,2011,18,(6):732-735.
[20] HUNG J,MCQUILLAN B M,CHAPMAN C M,et al.Elevated interleukin-18 levels associated with the metabolic syndrome independent of obesity and insulin resistance[J].Immunology,2005,96(2):247 -251.
[21] TROSEID M,SELJEFLOT I,HJERKINN E M,et al.Interleukin-18 is a strong predictor of cardiovascular events in elderly men with the metabolic syndrome:synergistic effect of inflammation and hyperglycemia[J].Diabetes Care,2009,32(3):486-492.
[22] 刘大彪.2型糖尿病患者血清 IL-6、IL-8、IL-10、IL-18检测的临床意义[J].放射免疫学杂志,2008,2(1):44-45.
猜你喜欢
红蜻蜓·低年级(2022年5期)2022-05-11
幼儿园(2021年16期)2021-12-06
数学物理学报(2021年4期)2021-08-30
现代临床医学(2021年4期)2021-07-31
中等数学(2020年1期)2020-08-24
红领巾·探索(2020年5期)2020-05-19
家教世界·创新阅读(2020年12期)2020-01-11
文化创新比较研究(2020年8期)2020-01-02
特别健康(2018年3期)2018-07-04
中西医结合心脑血管病杂志(2016年20期)2016-03-01
