
竹节草种质资源抗寒性初步评价研究
2016-05-30 10:48廖丽蒋仁娇刘建秀张欣怡陈菊梅白昌军王志勇徐毓皎
热带作物学报 2016年2期
廖丽 蒋仁娇 刘建秀 张欣怡 陈菊梅 白昌军 王志勇 徐毓皎
摘 要 以85份竹节草种质资源为材料,利用2014年和2015年观测的叶片电导率、田间枯黄率和形态数据为指标进行抗寒性分析。结果表明:不同竹节草种质资源间抗寒性达极显著相关(p<0.01),在一定程度范围内,植株密度越大,叶片宽度越大,坪用质量越高,则电导率越小,抗寒性越强。相关性分析结果表明,竹节草的叶片电导率和田间枯黄率与外部形态指标间存在极显著相关(p<0.01),相关系数最高达到0.867。竹节草抗寒性与植株密度、叶宽、坪用质量呈显著相关(p<0.05)。本研究为进一步开展竹节草优异种质选育提供理论依据。
关键词 竹节草;种质资源;抗寒性;相关性
中图分类号 S543.9 文献标识码 A
Evaluation of Chrysopogon aciculatus Accessions for Cold Tolerance
LIAO Li1, JIANG Renjiao1, LIU Jianxiu2, ZHANG Xinyi1,
CHEN Jumei1, BAI Changjun3, WANG Zhiyong1*, XU Yujiao1
1 Key Laboratory of Protection and Developmental Utilization of Tropical Crop Germplasm Resources,
Ministry of Education/ College of Agronomy, Hainan University, Haikou,Hainan 570228,China
2 Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing, Jiangsu 210014,China
3 Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, Danzhou, Hainan 571737,China
Abstract The cold resistance of Eighty-five Chrysopogon aciculatus(Retz.)Trin accessions were evaluated based on conductivity, leaf yellow rate and morphological characteristics. The results showed different C. aciculatus accessions were of extremely significant level(p<0.01). Within a certain degree, larger plant density led to wider leaf, and greater flat with higher quality resulted in smaller conductivity and stronger cold hardiness. There was extremely related(p<0.01) among conductivity, leaf yellow rate and morphological characteristics and the highest correlation index was 0.867. This study would provide a theoretical basis for the further development of excellent C. aciculatus accessions.
Key words Chrysopogon aciculatus;Germplasm resource;Cold resistance;Correlation analysis
doi 10.3969/j.issn.1000-2561.2016.02.004
竹节草[Chrysopogon aciculatus(Retz.)Trin]分布于世界热带和亚热带地区,在台湾、广东、广西、陕西及云南等省分布[1]。竹节草属于禾本科(Gramineae)金须茅(Chysopogon Trin.)属多年生草本植物,具根茎和匍匐茎。秆高20~50 cm,叶鞘无毛,叶多聚集于匍匐茎和秆的基部,秆生者稀疏或短于节间。叶片条形,顶端钝,长2~5 cm,宽3~6 mm。圆锥花序带紫色,长5~9 cm;分枝细弱;小穗数枚生于顶端。金须茅属植物在世界上约有20种,在中国有3个种,分别是刺金须茅(C. echinulatus)、金须茅(C. orientalis)和竹节草[2]。竹节草是金须茅属中唯一适合作草坪草的草种,由于植株低矮,弹性好,易繁殖,蔓延快,优良的水土保持植物,且成坪性好,是用于庭园、运动场和固土护坡的优良暖季型草坪草之一。但冬季低温,竹节草叶色变成黄褐色,降低了其观赏价值和应用前景[3]。
目前国内外学者对竹节草研究甚少[2]。在国内,廖丽等[2]对一些竹节草的种质资源进行搜集整理,并在田间对其进行过一些生态适应性进行相关评价。刘世忠等[4]用不同水肥处理对竹节草等3种草坪草延长冬季绿期的效果进行研究。刘建秀等[5]对竹节草在中国的分布范围进行调查。由于暖季型草坪草的抗寒性较弱,冬季都出现一定的枯黄休眠期,导致其观赏价值和使用价值降低[6]。但对竹节草种质资源的抗寒性研究尚未开展。本研究利用两年的观测指标(电导率和枯黄率)对85份竹节草的种质资源抗寒性进行初步评定,以明确竹节草抗寒性差异,进而鉴定竹节草种质资源抗寒性,对指导抗寒竹节草的育种及栽培具有重大意义,为选育优质品系奠定基础。
1 材料与方法
1.1 材料
1.1.1 试验基地概况 实验基地位于海南大学海甸校区农学院实验基地,地理坐标为北纬20°03′,东经110°19′,海拔22.9 m,属于热带季风气候,太阳辐射强,光热充足。实验前该地肥力中等,有灌溉条件。
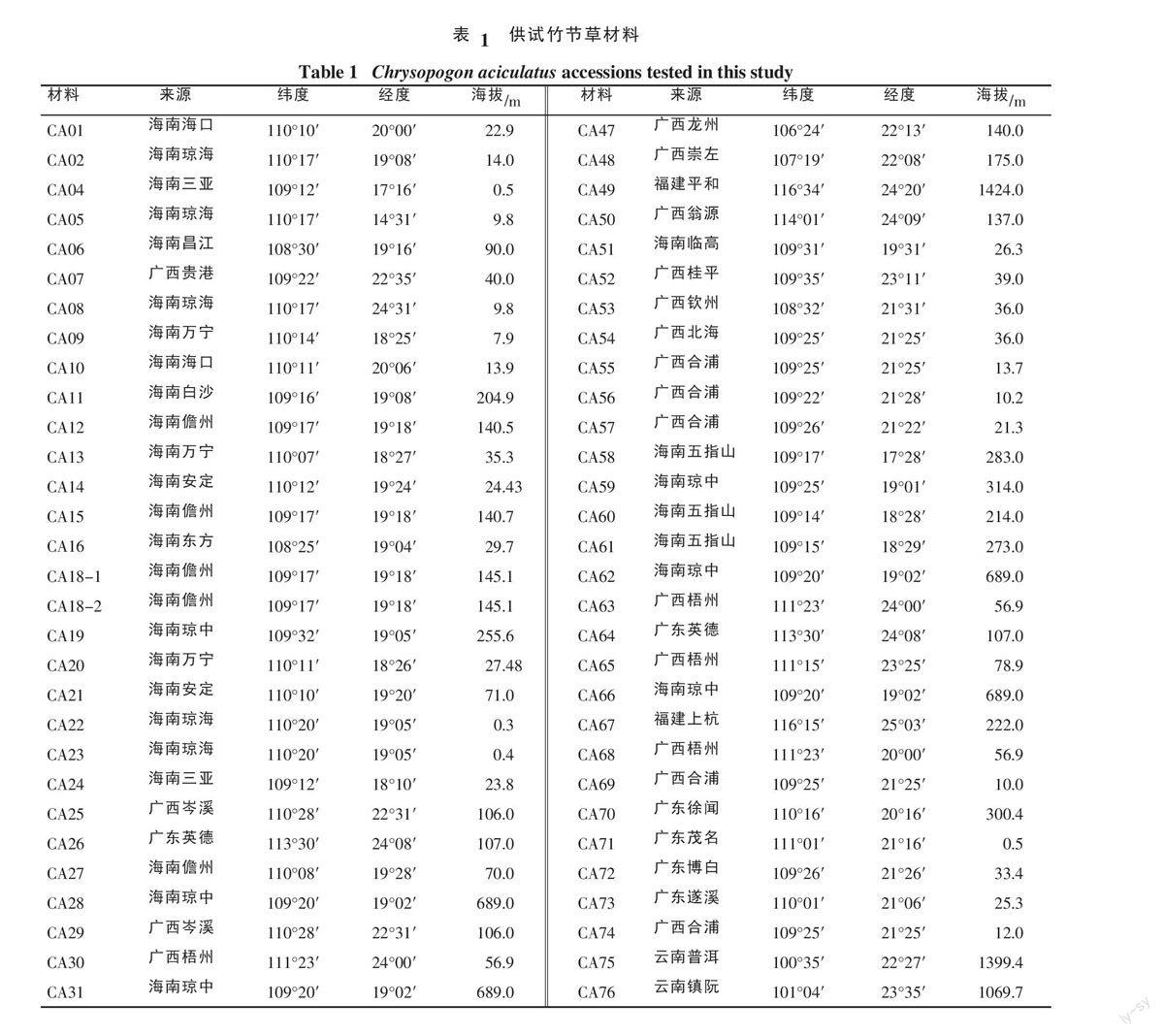
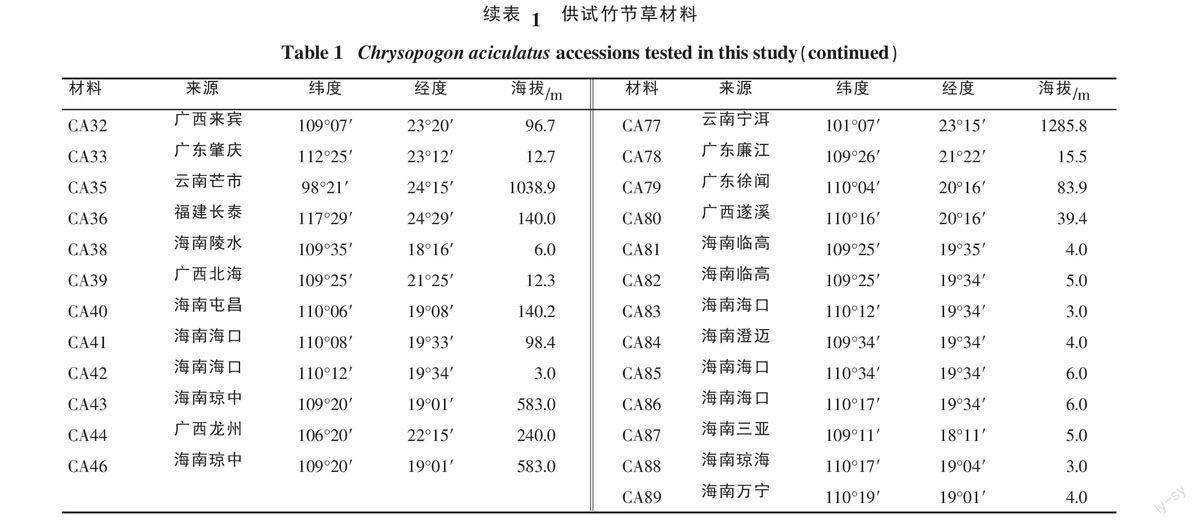
1.1.2 试验材料 85份竹节草种植资源主要来源云南、广东、广西、福建和海南5个省如表1所示。
1.2 方法
1.2.1 指标的测定和计算 (1)生殖枝高度。分别测量每块地的竹节草的生殖枝高度(从地面到花穗顶部的自然高度),重复10次。
(2)叶长和叶宽。匍匐茎的倒数第四叶片的长和宽,用米尺测量,重复10次。
(3)密度。草坪密度是指单位面积上草坪植物个体或枝条的数量。选择目测法,是以目测估计单位面积内的草坪植物的数量,并人为地划分一些密度等级,以此来对草坪密度进行分级或打分。本实验采用十分制,其中1表示极差,5表示中等,10表示优。
(4)坪用质量。根据草坪的刚性,弹性、光滑度、青绿度等的指标综合考虑。采用十分制定分[7]。
(5)花序分枝数。随机选取小区内的10枝花序,计算每个花序的分枝数,求平均值。
(6)电导率测定。以健康的竹节草为样品,采集时需要注意采集同一位置的叶片,在现场选择完好的叶片装入保鲜袋中,带回实验室做叶片的电导率测定。竹节草带回实验室中,并在12 h内,用去离子水清洗采集来的样品,剪成0.5~1.0 cm,每份称取1 g,每份3个平行样,并将它们分别放入试管中,贴好标签。在常温下,分别在试管中加入30 mL去离子水,用保鲜膜封好试管,静置24 h。24 h之后开始测第一次电导率(R1)。将测完电导率的样品放入沸水恒温水浴锅中,水煮30 min,煮好后冷却,待测它的电导率(R2)。
叶片相对电导率=R1/R2[8]。
(7)叶片枯黄率。叶片枯黄率采用目测打分法记录各材料叶片枯黄率(采用百分制,5%以下表示竹节草没有黄叶出现;50%表示竹节草有一半枯黄;95%以上表示基本没有绿叶而死亡)[9]。
1.2.2 数据处理 运用EXCEL 2000和SPSS16.0 软件对数据进行分析和处理。
2 结果与分析
2.1 竹节草种质资源的相对电导率与叶片枯黄率、形态指标的相关性
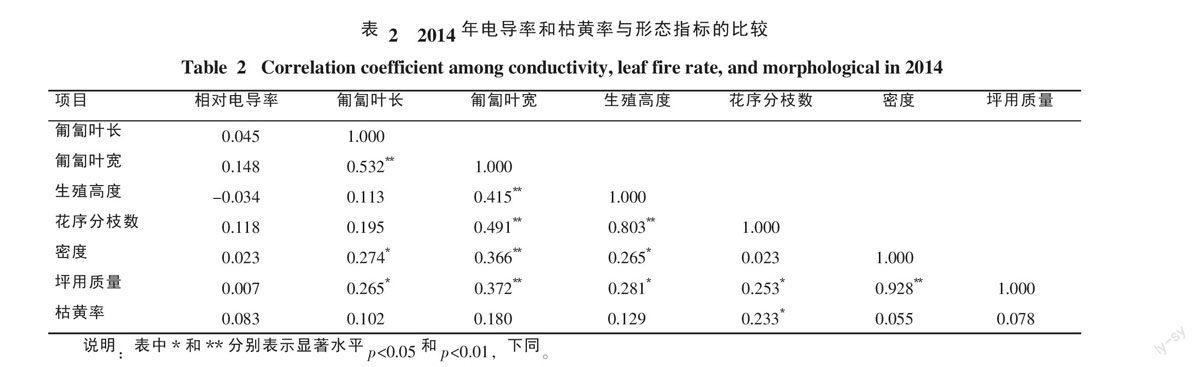
从表2可知枯黄率与花序分枝数存在显著正相关(p<0.05),相关系数为0.233。从表3可知相对电导率与花序分枝数存在显著正相关(p<0.05),相关系数为0.239;枯黄率与密度存在极显著正相关(p<0.05),相关系数为0.298;枯黄率与坪用质量存在显著正相关(p<0.05),相关系数为0.265。从表4可见,相对电导率与坪用质量和密度存在极显著负相关(p<0.01),相关系数为-0.388和-0.347,与叶宽存在显著正相关(p<0.05),相关系数为0.249;坪用质量与密度存在极显著正相关(p<0.01),相关系数为0.867;生殖枝高度与花序分枝数存在极显著正相关(p<0.01),相关系数为0.475,与匍匐叶宽存在显著正相关(p<0.05),相关系数为0.257;花序分枝数与叶宽存在极显著正相关(p<0.01),相关系数为0.368,与密度存在极显著负相关(p<0.01),相关系数为-0.313;匍匐叶宽与匍匐叶长存在极显著正相关(p<0.01),相关系数为0.464。竹节草抗寒性越强,植株密度越大,匍匐叶片宽度越大,坪用质量越高。
2.1.1 竹节草相对电导率和叶片枯黄率的相关性 从表5可见,2014年和2015年的相对电导率和叶片枯黄率相关性不显著。
2.2 竹节草种质资源间枯黄率和相对电导率之间的差异分析
85份竹节草种质资源的相对电导率和相对枯黄率的多重比较结果表明如表6,不同地域竹节草间抗寒性达到显著(p<0.05)或极显著水平(p<0.01)。相对电导率1的变异范围是12.67%~43.33%,变异系数是36.80%;叶片枯黄率1的变异范围是3.9%~57.1%,变异系数是50.16%;相对电导率2的变异范围是10.0%~85.0%,变异系数是47.22%;叶片枯黄率2的变异范围是6.67%~95.67%,变异系数是43.39%;相对电导率的变异范围是12.40%~81.10%,变异系数是43.195%;叶片枯黄率的变异范围是11.20%~46.00%,变异系数是38.23%。变异范围表明不同的种质间的差异大小,对品种抗寒性选育中有一定的参考价值。
3 讨论与结论
植物的相对电导率的变化能反应出植株抗寒性的大小,可作为抗寒性筛选的指标。本试验主要是对85份自然低温条件下,竹节草种质资源的抗寒性进行研究。通过对竹节草外部性状与相对电导率的相关性分析,相对电导率与坪用质量和密度存在极显著负相关(p<0.01),与叶宽存在显著正相关(p<0.05);坪用质量与密度存在极显著正相关;生殖枝高度与花序分枝数存在极显著正相关(p<0.01),与叶宽存在显著正相关(p<0.05);花序分枝数与匍匐叶宽存在极显著正相关(p<0.01),与密度存在极显著负相关(p<0.01);匍匐叶宽与匍匐叶长存在极显著正相关(p<0.01)。这说明在一定程度范围内,植株密度越大,叶片宽度越大,坪用质量越高,则电导率越小,抗寒性越。
但从表2可知枯黄率与花序分枝数存在显著正相关(p<0.05),相关系数为0.233。而从表3可知相对电导率与花序分枝数存在显著正相关(p<0.05),相关系数为0.239;枯黄率与密度存在极显著正相关(p<0.01),相关系数为0.298,与坪用质量存在显著正相关(p<0.05),相关系为0.265。然而,本研究中竹节草相对电导率与枯黄率相关性不显著(p<0.05)。
本研究以2015年(相对电导率1和叶片枯黄率1)、2014年(相对电导率2和叶片枯黄率2)以及2014年与+2015年(相对电导率3和叶片枯黄率3)相对电导率和叶片枯黄率对竹节草抗寒性进行研究,其中,2015年的相对电导率1的变异范围是12.67%~43.33%,叶片枯黄率1的变异范围是3.9%~57.1%;2014年相对电导率2的变异范围是10.0%~85.0%,叶片枯黄率2的变异范围是6.67%~95.67%;2014年与2015年的相对电导率3的变异范围是12.40%~81.10%,叶片枯黄率3的变异范围是11.20%~46.00%,这6个指标的变异范围均达极显著(p<0.01)水平。本研究表明叶片枯黄率对叶片相对电导率存在影响,进而使竹节草种质资源的抗寒性表现出丰富的差异。王晓玲等[10]已证明在植物抗寒性中,电导率越大抗寒性越低。
85份竹节草种质分别从广东,广西,福建,云南等地收集,竹节草的抗寒性的差异原因可能是地域差异造成植物对外界生境适应行差异。总之,我国华南地区蕴藏丰富的竹节草种质资源,本研究结果将对今后深入开展竹节草抗寒性机理和优异抗逆育种提供参考。
参考文献
[1] 郑玉忠, 徐嘉宾, 杨中艺,等. 中国竹节草野生种质资源调查及生物学特性研究[J]. 草业学报, 2005, 14(3): 117-122..
[2] 廖 丽, 王志勇, 白昌军,等. 竹节草种质资源形态多样性研究究[J]. 热带作物学报, 2011, 32(11): 2 042-2 047.
[3] 王鹏良, 徐 洋, 吕智鹏, 等. 假俭草杂种F1抗寒性遗传分析[J]. 草业学报, 2011, 20(2): 290-294.
[4] 刘世忠, 夏汉平, 蔡锡安, 等. 不同水肥处理对三种草坪草延迟冬绿期效果比较[J]. 中国草地, 2002, 24(4): 25-30.
[5] 刘建秀, 刘永东, 贺善安, 等. 中国暖季型草坪草物种多样性及其地理分布特点[J]. 草地学报, 1998, 6(1): 45-52.
[6] 刘 琳, 毛 凯, 干友民,等. 暖地型草坪草抗寒性的研究概况[J]. 草原与草坪, 2004(1): 8-14.
[7] 刘建秀. 草坪坪用价值综合评价体系的探讨I.评价体系的建立[J]. 中国草地, 1998(1): 44-47.
[8] 林 艳, 郭伟珍, 徐振华,等. 大叶女贞抗寒性及冬季叶片电导率变化的研究[J]. 天津农业科学, 2012, 18(5): 145-149.
[9] 廖 丽, 张 静, 吴东德,等. 竹节草种质资源耐盐性初步评价[J]. 热带作物学报, 2014, 35(10): 1 905-1 911.
[10] 王晓玲, 胡亚岚, 毛丽衡,等. 不同枣品种抗寒性的比较[J]. 北方园艺, 2012, 19: 1-4.
[11] Subba R C, Raju N S N, Subba R M V. Pollination and seed set in tropical wetland grasses[J]. Nordic Journal of Botany, 2010, 28(3): 354-365.
猜你喜欢
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
吉林农业·下半月(2016年11期)2017-01-09
科技视界(2016年21期)2016-10-17
中国实用医药(2016年24期)2016-10-17
中国实用医药(2016年24期)2016-10-17
中国实用医药(2016年24期)2016-10-17
新疆农垦科技(2016年2期)2016-08-21
中国果菜(2016年9期)2016-03-01
