
植物镁素生理功能及镁素营养诊断和施用
2016-05-30 11:58:40林敏霞张晓东邱美欢张洪溢王丹
热带农业科学 2016年3期
关键词:生理功能
林敏霞 张晓东 邱美欢 张洪溢 王丹
摘 要 随着土壤缺镁状况日益突出,缺镁对植物的正常生理功能和生长发育的影响越来越明显,因此镁肥的施用越来越受到重视。综述了植物镁素的生理功能,主要包括光合生理功能、酶活化功能、植物镁素对植物活性氧代谢和基因表达的影响,以及植物镁素营养诊断与镁肥的施用,为植物镁素营养的深入研究提供理论依据和参考。
关键词 镁素 ;生理功能 ;营养诊断 ;镁肥
分类号 S15 Doi:10.12008/j.issn.1009-2196.2016.03.009
Abstract With the deficiency of Magnesium (Mg) more and more obviously, and it affects the normal physiological function and growth of plants. Thus, people have paid more attention for application of Mg-containing fertilizers. The physiological function, nutrition diagnosis and application of Mg in plants were reviewed in this article. In addition, the physiological function includes photosynthesis physiological function, activating function of enzyme, the impact of oxygen metabolism and gene expression in plant. It will provide bases and references for the further study of Mg in the future.
Keywords magnesium ; physiological function ; nutrition diagnosis ; magnesium-containing fertilizers
镁素是植物所必需的中量元素之一,对植物生长发育起至关重要的作用。镁是植物细胞中重要的二价阳离子,是叶绿素的中心离子,还是很多酶的活化剂,参与能量代谢,对提高作物产量和森林蓄积量等具有重要作用[1-2]。但是,随着工业和农业产业的快速发展,很多地区酸沉降现象加剧,镁在酸性土壤中溶解流失加重[3],因此,缺镁现象普遍存在于植物中,中国许多地方出现植物缺镁失绿的状况,仅中国土壤缺镁面积就占了全国总耕地面积的6%[4-5],影响了农业生产的多个领域。因此,本文综述了镁素的各种生理功能,主要体现在光合生理功能、酶活化功能、其对植物活性氧代谢和基因表达的影响等方面,并对植物镁素营养失衡诊断及镁肥的施用进行综述,以期为更好地利用镁元素提高作物的产量和品质提供参考。
1 镁素生理生化功能
1.1 镁素的光合生理功能
镁素最重要的生理功能就是其作为叶绿素的中心离子所起的作用[6],镁可以占叶绿素分子量的2.7%,参与叶绿素(chl)、色素的构成,大约10%的镁结合在chl a和chl b中[7],对维持叶绿体结构起到非常重要的作用。但是植物缺镁时叶片叶绿素含量降低的真正原因,目前还存在争议。许多学者认为,由于镁是叶绿体正常发育所必需的元素,并且需求量很大,因此,缺镁时镁的不足是导致叶片失绿的原因;而有部分学者认为,缺镁导致蛋白合成受阻才是缺镁植物失绿的原因[8];Marschner等[9]和Cakmak等[10]研究结果发现,缺镁胁迫下的活性氧(ROS)受伤害才是叶绿素含量降低和叶片失绿的主要原因。研究证明,Mg2+可以影响植物光合膜的垛叠形成基粒,调节PSⅠ和PSⅡ 2个光系统之间的激发能分配、PSⅡ活性和原初光能的转化效率、光合电子传递速率、叶绿素荧光和光合碳代谢等过程[4]。凌丽俐等[11]研究表明,缺镁导致纽荷尔脐橙老叶叶绿素含量明显降低,但对新叶无影响。缺镁胁迫和过量镁胁迫均导致纽荷尔脐橙叶片的光化学效率(Fv/Fm)以及相对电子传递速率(rETR)降低,但是,缺镁胁迫的影响作用大于过量镁胁迫。李泽等[12]研究证实,低镁(Mg2+<2 mmol/L)胁迫显著降低了油桐叶片中叶绿素含量、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、最大PSⅡ的潜在活性等;而高镁(Mg2+>4 mmol/L)同样也降低了叶绿素含量、Pn、Gs等,但是镁浓度升高对光合生理指标的影响要小于缺镁胁迫的作用。
1.2 酶活化功能
镁是植物酶的重要组成部分,是植物体内多种酶的活化剂,几乎所有的磷酸化酶、磷酸激酶、二磷酸核酮糖羧化酶都需要镁的激活或者活化,若镁缺乏会抑制CO2的同化,从而影响光合作用[13-15]。Mg2+可以与ATP或ADP的焦磷酸盐结构和酶分子间呈桥式结合,从而促进磷酸化作用,其作用的机制是改变ATP酶的构象,使得埋藏在酶内部的结合态核苷酸更易暴露出来,更容易接近底物。镁对磷酸激酶和磷酸转移酶的活化是专性的[4-16]。
1.3 镁素对植物活性氧代谢的影响
叶绿体类囊体膜PSI的还原侧是植物活性氧(ROS)产生的重要部位[17]。植物一旦缺镁会降低卡尔文(Calvin)循环的效率,CO2同化受到限制,光合同化力NADPH/NADP+会累积,供给暗反应的能量随之减少,也就减少了NADPH的利用率,导致光合作用电子传递系统过度饱和,从而激发产生过ROS,过ROS传递到分子氧O2后会产生超氧阴离子O2-1等[18-19],由此衍生的羟自由基(OH-)和单线氧(1O2)等其他ROS均具有很强的活性,它们可以对植物的光合作用等许多生物功能造成伤害[20]。
但是,植物本身具有抗氧化能力,可通过产生一系列的抗氧化酶,如超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)和过氧化氢酶(CAT)等,以及小分子抗氧化物质,如还原型谷胱甘肽(GSH)、抗坏血酸(ASC)和维生素E等,来消除过量的ROS[7]。陈伟立等[21]研究发现,缺镁处理砂糖橘后植株叶片的叶绿素含量显著降低,并且POD和CAT活性显著提高,SOD和APX活性变化不明显,该结果说明,缺镁引起砂糖橘叶片中活性氧的积累,而POD和CAT可能参与了活性氧的清除,这与薄荷、玉米、龙眼等在缺镁处理后出现的植物体内抗氧化酶活性增强、小分子抗氧化剂含量明显增加的结果相同[22-23]。朱帅等[24]于低温下用30%镁处理黄瓜幼苗,结果发现幼苗植株中丙二醛(MDA)含量增加,但SOD、POD、CAT、APX和GR等活性降低。申燕等[25]研究发现,缺镁使“不知火”橘橙和椪柑叶片SOD活性显著降低,但二者的POD活性、MDA含量及“不知火”橘橙的CAT活性均显著提高,此结果说明“不知火”橘橙和椪柑虽然对缺镁胁迫均有一定的抵御和适应能力,但随着缺镁处理时间的延长,体内ROS的累积超过其保护酶的清除能力,其抗氧化防御系统已不能抵御质膜所受到的ROS的攻击,丧失对膜系统的保护,最终导致细胞伤害的发生和伤害程度的差异[26]。
1.4 镁素对植物基因表达的影响
镁在植物基因组的稳定中起到重要的作用[27]。当植物缺镁时,体内的核酸含量就会下降,某些正常情况下表达的基因不表达,而原来不表达的基因开始表达[4]。例如,拟南芥缺镁时,编码1-氨基环丙烷-1羧酸酶家族同工酶的4个基因(ACS11、ACS2、ACS7、ACS8)被诱导表达;在拟南芥的根和叶片中,ACS11基因表达上调,而ACS2、ACS7和ACS8基因只在叶片中表达上调。持续缺镁7 d后,缺镁处理组乙烯的释放量是对照组的2倍,同时研究发现,运输镁的通透酶也未表达,只有MRS2-9基因表达水平显著下调[28]。
2 植物镁素营养的诊断
植物缺镁是一个普遍存在的问题。镁是植物叶绿素的重要成分,因此,缺镁最明显的表现就是叶片失绿。而镁的移动性较强,所以,失绿首先表现在老叶,缺镁情况严重时会导致整片叶片发黄干枯[29-30]。李金玲等[31]采用砂培的方法,研究缺镁对一年生何首乌的影响,结果表明,缺镁胁迫导致何首乌叶片颜色失绿,叶绿素含量下降,地上部生长较差,膨大的块根减少。Tewari等[20]研究发现,缺镁症状首先表现在桑树的老叶边缘,继而向主脉蔓延,出现黄色斑点,随后斑点不断变大,颜色加深,最终形成坏死斑点,从而导致叶片凋落。植物缺镁除了出现叶片失绿等现象外,还表现在枝干细长柔弱、根数量减少、开花缓慢、花色不鲜亮、根茎比下降等方面[30]。彭昊阳[29]利用蛋白质组学研究了缺镁胁迫对雪柑的影响,结果表明,雪柑叶片中有89个蛋白发生了改变,包含58个上调蛋白,31个下调蛋白;根系中有32个蛋白发生了改变,包含20个上调蛋白,12个下调蛋白,表明缺镁胁迫对雪柑叶片和根蛋白的影响不同。此外,缺镁胁迫还导致Rubisco、Rubisco活化酶、类光合电子传递蛋白、FNR等多种参与光合作用的酶水平下降。靳晓琳[32]采用cDNA-AFLP 技术筛选并分析芦柑中与缺镁胁迫相关的基因。结果发现,芦柑叶对缺镁胁迫的响应主要涉及光合作用、碳水化合物代谢、细胞组织合成、逆境响应、氧化还原反应、蛋白质代谢、信号转导、转录翻译和转运等过程。
3 镁肥种类及施用
3.1 镁肥种类
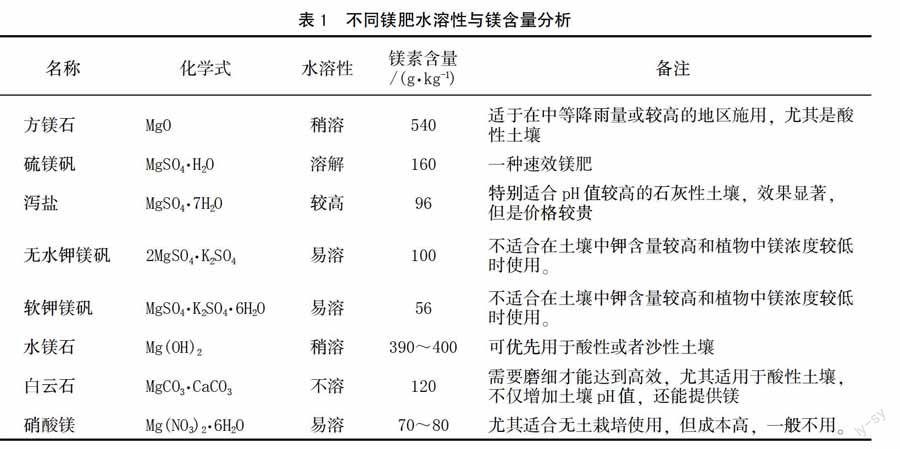
凡是含有镁素的化合物原则上都可以作为镁肥。目前一般使用的都是无机物,根据其溶解性不同可以分为水溶性、枸溶性和难溶性[33]。根据物质组成形式还可以将镁肥分为镁的氧化物、硫酸盐、碳酸盐、硝酸盐、磷酸盐、硅酸盐和氯化物等,具体分析见表1[30,34]。
3.2 镁肥施用
镁肥可作为基肥或者追肥。水溶性的镁肥一般可以表施,而溶解性小的可与土壤混施。一般来说,镁肥必须与其他肥料配施效果才好。中国南方地区为酸性土壤,镁素易淋失,尤其需要施加镁肥。早在20世纪60年代,谢建昌等[35]在南方12块红壤稻田和6块红壤旱地上进行了水稻、大豆镁肥肥效试验。结果发现,施用镁肥后,水稻平均增产10%左右,大豆平均增产23.5%。到70年代,海南大面积的橡胶由于缺镁而发生“鱼骨状”黄化病,此后镁肥开始被使用。80年代,南方地区陆续有关于多种作物出现缺镁症状的报道,如剑麻出现的茎枯病[36],水稻出现的火烧黄叶病[37]等。据文献报道,在南方酸性土壤上施用镁肥效果显著,如海南保亭县畅好农场连续施用镁肥15 a后,缺镁导致的橡胶黄化病发病率由原来的50.7%下降至16.0%,产量净增7%,而未施镁素的对照区,黄化病则越来越严重,发病率达90.0%以上[30]。周晓琳等[38]研究了叶面施镁肥对结球甘蓝产量与品质的影响。结果发现,施加MgSO4可以增大甘蓝球的横、纵径以及提高产量,并且增产可达5.97%~32.09%。王丹等[1]研究发现,镁可以显著促进冬季生长迟缓期的艾纳香生长,显著提高了艾纳香叶、茎和根生物量。其中10、15 g/L七水硫酸镁处理下的艾纳香叶片和茎生物量分别是对照组的10.08和5.28、10.91和5.50倍。高华军等[39]对不同施镁方式和施镁时间进行了研究。结果表明,施镁肥时,基施+追施可提高烤烟生长中期土壤中的镁素含量,增加烤后烟叶镁素含量(较对照组增加了12.68%),并明显促进植物生长发育、改善植物农艺性状。随着土壤缺镁面积的不断增加和人们对作物品质改善的要求,通过施用镁肥来提高作物产量和品质的方式也将越来越受到人们的重视,相关的研究报道也会越来越多。
参考文献
[1] 王 丹,范佐旺,庞玉新,等. 外源镁对冬季迟缓期的艾纳香生物量和有效成分含量的影响[J]. 中国实验方剂学杂志,2015,21(4):75-79.
[2] Beale S I. Enzymes of chlorophyll biosynthesisi[J]. Photosynthesis research, 1999(60): 43-73.
[3] Hermans C, Bourgis F, Faucher M, et al. Magnesium deficiency in sugar beet alters sugar partitioning and phloem loading in young mature leaves[J]. Planta, 2005, 220(4): 541-549.
[4] 熊英杰,陈少凤,李恩香,等. 植物缺镁研究进展及展望[J]. 安徽农业科学,2010,38(15):7 754-7 757.
[5] 靳晓琳,马翠兰,陈立松. 植物缺镁研究进展[J]. 亚热带农业研究,2012,8(2):118-122.
[6] 李 娟. 植物钾、钙、镁素营养的研究进展[J]. 福建稻麦科技,2007,25(1):39-42,30.
[7] 郑重禄. 缺镁胁迫对柑橘的影响研究综述(2)——缺镁对柑橘光合作用及相关生理生化的影响[J]. 浙江柑橘,2015,32(3):2-8.
[8] Marschener H. Mineral nutrition of higher plants [M]. London: Academic Press Inc, 1986, 235-243.
[9] Marschner H, Cakmak I. Hight light intensity enhances chlorosis and necrosis in leaves of zinc, potassium,and magnesium deficient bean(Phaseolus vulgaris)plants[J]. Journal of Plant Physiology, 1989, 134(3): 308-315.
[10] Cakmak I, Hengeler C, Marschenser H. Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphrus, potassium and magnesium deficiency[J]. Journal of Experimental Botany, 1994, 45(9): 1 245-1 250.
[11] 凌丽俐,朱春钊,彭良志,等. 镁胁迫对纽荷尔脐橙叶绿素荧光日变化特性的影响[J]. 西南大学学报(自然科学版),2014,36(6):67-73.
[12] 李 泽,谭晓风,卢 锟,等. 供镁水平对油桐幼苗生长及光合特性的影响[J]. 生态学杂志,2015,34(9):2 440-2 447.
[13] 尹永强,何明雄,韦峥宇,等. 烟草镁素营养研究进展[J]. 广西农业科学,2009,40(1):60-66.
[14] 杨军芳,周晓芬,冯 伟. 土壤与植物镁素研究进展概述[J]. 河北农业科学,2008,12(3):91-93,96.
[15] Cakmak I, Yazici A M. Magnesium: A forgotten element in crop production[J]. Better Crops, 2010, 94(2): 23-25.
[16] 李 延,刘星辉,庄卫民. 植物Mg素营养生理的研究进展[J]. 福建农业大学学报,2000,29(1):74-80.
[17] 余叔文,汤章城. 植物生理与分子生物学(第二版)[M]. 北京:科学出版社,1998:307.
[18] Winston G W. Physiochemical basis for free radical formation in cells: production and defenses [M]//Alschor R G, Cumming J R. Stress responses in plants: adaptation and acclimation mechanisms. New York: Wiley-liss, 1990: 57-86.
[19] Marschner H. Mineral nutrition of higher plants(2nd Ed)POD [M]. London, UK: Academic Press, 1995: 313-320.
[20] Tewari R K, Kumar P, Sharma P N. Magnesium deficiency induced oxidative stress and antioxidant responses in mulberry plants[J]. Scientia Horticulturae, 2006, 108: 7-14.
[21] 陈伟立,谢小林,李 娟,等. 缺镁胁迫对‘砂糖橘植株矿质养分及抗氧化酶的影响[J]. 热带农业科学,2015,35(6):5-10.
[22] Candan N, Tarhan L. Relationship among chlorophyll-carotenoid content, antioxidant enzyme activities and lipid peroxidation levels by Mg2+ deficiency in the Mentha pulegium leaves[J]. Plant Physiology Biochemistry, 2003, 41: 335-40.
[23] Tew Arirk, Kumar P, Tew Arin, et al. Macronutrient deficiencies and differential antioxidant responses-influence on the activity and expression of superoxide dismutase in maize [J]. Plant Science, 2004, 166: 187-694.
[24] 朱 帅,吴帼秀,蔡 欢,等. 低镁胁迫对低温下黄瓜幼苗光合特性和抗氧化系统的影响[J]. 应用生态学报,2015,26(5):1 351-1 358.
[25] 申 燕,肖家欣,杨 慧,等. 镁缺乏对“不知火”和“椪柑”生理指标、镁分布及叶片超微结构的影响[J]. 浙江大学学报(农业与生命科学版),2011,37(4):446-452.
[26] 郑重禄. 缺镁胁迫对柑橘的影响研究综述(1)-缺镁对柑橘生长发育的影响[J]. 浙江柑橘,2015,32(2):7-10.
[27] 姚嘉宜,刘利英,宋土生. 镁在基因组稳定中的作用[J]. 国外医学医学地理分册,2003,24(3):115-118.
[28] Hermans C, Vuylsteke M, Coppen F, et al. Systems analysis of the responses to long-term magnesium deficiency and restoration in Arabidopsis thaliana[J]. New Phytologist, 2010, 187(1): 132-144.
[29] 彭昊阳. 缺镁胁迫下雪柑根叶蛋白质组学研究[D]. 福州:福建农林大学,2013.
[30] 汪 洪,褚天铎. 植物镁素营养诊断及镁肥施用[J]. 土壤肥料,2000(4):4-8.
[31] 李金玲,熊寅森,赵 致,等. 钙镁元素缺乏对何首乌生长发育的影响[J]. 贵州农业科学,2012,40(11):68-70.
[32] 靳晓琳. 缺镁胁迫下柑橘幼苗cDNA-AFLP分析[D]. 福州:福建农林大学,2013.
[33] 李明晶,孙华云,刘亦凡. 镁在植物生长中的作用及镁肥[J]. 硫磷设计与粉体工程,2009(4):36-39,52.
[34] 郭如新. 镁肥肥源及其应用[J]. 磷肥与复肥,2011,26(1):50-52.
[35] 谢建昌,陈际型,朱月珍,等. 红壤区几种主要土壤的镁素供应状况及镁肥肥效的初步研究[J]. 土壤学报,1963,1(3):294-305.
[36] 华南热作研究院橡胶栽培研究所营养诊断课题组. 我国橡胶树的缺镁症及其防治的研究[J]. 热带作物研究,1985(2):1-10.
[37] 李伏生. 红壤地区镁肥对作物的效应[J]. 土壤与环境,2000,9(1):53-55.
[38] 周晓琳,李洪杰,赵同凯,等. 叶面喷施钙镁肥对结球甘蓝养分积累及产量的影响[J]. 山东农业科学,2015,47(9):98-100.
[39] 高华军,林北森,杨小梅,等. 镁肥施用方法和时期对烤烟镁含量及产量、品质的影响[J]. 河南农业科学,2013,42(10):36-40.
猜你喜欢
国外畜牧学·猪与禽(2022年1期)2022-04-23 21:22:06
昆明医科大学学报(2020年12期)2021-01-26 00:44:14
猪业科学(2018年4期)2018-05-19 02:04:56
广东饲料(2016年4期)2016-12-01 03:43:17
广东饲料(2016年8期)2016-02-27 11:10:02
湖北农业科学(2015年15期)2015-09-09 21:36:59
宠物世界·狗迷(2015年7期)2015-05-30 09:54:42
中国粮油学报(2015年5期)2015-02-06 01:47:19
南方农业·下旬(2014年5期)2014-08-30 20:18:26
河南科技(2014年7期)2014-02-27 14:11:10
