
喀斯特重度石漠化地区苔藓植物优势种的种间关联性研究
2016-05-17 11:24:40殷声锋王智慧张朝晖
生态科学 2016年5期
殷声锋, 王智慧, 张朝晖,*
喀斯特重度石漠化地区苔藓植物优势种的种间关联性研究
殷声锋1, 王智慧2, 张朝晖1,*
1. 贵州师范大学, 贵州省山地环境信息系统与生态环境保护重点实验室, 贵州 贵阳 550001
2. 贵州师范大学, 生命科学学院, 贵州 贵阳 550001
苔藓植物在喀斯特生态结构中占据着极其重要的地位, 在维系生态系统生态多样性方面发挥着重要的生态功能。以贵州典型喀斯特地区—贵阳市花溪区为研究对象, 对苔藓植物的物种分布特征进行了调查, 并运用X2检验, Pearson相关系数检验, Spearman秩相关系数检验和主成分分析(PCA)进行分析, 研究结果表明: 本地区共有苔藓植物5科11属30种; 通过对14种优势苔藓植物的关联性分析, 发现苔藓植物物种间总体上呈显著的负联结性, 说明该苔藓群落正处于演替的初级阶段; 正负关联比(小于1)说明苔藓植物生态位分离; 根据PCA分析结果, 划分2个生态种组, 分别是分布于背阴岩壁和分布于向阳岩面的苔藓植物。
喀斯特重度石漠化; 苔藓植物; 种间关联; PCA
1 前言
种间关联(Interspecific association)是指异种种群之间的在空间分布上的相互关联性, 是植物种群间相互联系和相互影响的综合反映, 通常是由于群落生境的差异影响了物种的分布而引起的[1–3]。群落的种间关联性, 不仅能够有效地反映各物种在群落中的分布情况及各物种对环境因子的适应程度, 而且能反映物种在特定环境因子作用下的种间相互关系[4]。种间关联的研究是物种群落结构稳定程度与群落演替的重要基础[5]。国内对于种间关联的研究主要集中在森林木本和草本上的研究[6–10]。
苔藓植物作为喀斯特重度石漠化地区的主要物种[11], 对其的研究主要集中于苔藓植物区系、生态和生物喀斯特沉积等[12–14]。在其种间关联方面的研究尚未见报道。本文研究了喀斯特重度石漠化地区的优势苔藓植物关联性和相关性, 了解喀斯特地区苔藓植物物种组成结构和植物演替进程, 为石漠化地区植被恢复与重建提供理论依据。
2 研究区域概况
采样地点位于贵州省贵阳市花溪区青岩镇龙井村东部, 东经106°39′22.3″, 北纬26°20′29.2″, 海拔(1106±4) m。该区域为典型的喀斯特溶沟地貌, 具明显的脊背, 石壁和沟底。区域内为裸露的碳酸岩, 仅在沟底有少量土壤, 平均裸露岩石面积达90%, 为重度石漠化区。研究区域内, 苔藓植物生长于岩面或岩壁上, 除此之外, 偶见其他高等植物。
该地区气候类型为亚热带季风湿润气候, 年均温度为14.9 ℃, 年平均降雨量1100—1200 mm, 无霜期270天, 日照时数1354小时[15]。
3 材料与方法
3.1野外样本采集
研究区域内按生态学系统取样的样地设置方法随机选取五条溶沟, 每条溶沟分别在脊背、石壁和底部各设置10个10 cm×10 cm的样方进行采样, 即每条溶沟采集30份样本。区域内共采集150份样本,带回实验室进行标本鉴定。
3.2标本鉴定
将采集的标本置于阴凉处晾干后, 装入标本袋中。标本鉴定采用经典的苔藓植物形态学鉴定方法,借助HWG-1双筒解剖镜和SMARTe-320一体化数码显微镜观察并绘图或拍照。重点参照《中国苔藓志》(第一、二、四, 七, 八卷)[16–19]及《中国生物物种名录》(第一卷)[20]。
3.3数据分析方法
3.3.1 重要值计算采用重要值表示苔藓植物物种的优势度
重要值=(相对频度+相对盖度)/2
3.3.2 总体关联性检验
总体关联性是指一个群落所有种类的关系, 通过计算物种间联结指数(VR)来测定所研究种类的总体关联性。
种间关联性分析 测定在一定的置信水平上检验两个种间是否存在关联, 一般用X2检验。为种间关联性的定性指标。
Pearson相关分析和Spearman秩相关分析 使用盖度数据计算种间相关系数来衡量两个种间的相关程度。为种间关联性的定量指标。
以上指标计算公式, 参见张金屯所编《数量生态学》一书[21]。
3.3.3 数据运算
所有数据运算与统计在Microsoft Excel、R语言和Canodraw for Windows 5.0中完成。
4 结果与分析
4.1苔藓植物物种多样性
根据野外调查结果, 在该采样地点内共记录苔藓植物5科11属30种, 其中优势科(≥3属)为丛藓科(Pottiaceae)(4属)和灰藓科(Hypnaceae)(4属), 优势属(≥5种)为真藓属(Bryum)(7种)、湿地藓属(Hyophila)(6种)和扭口藓属(Barbula)(5种)。根据种的重要值(表1), 该喀斯特地区内呈现出少数几个物种所主导的格局: 美灰藓(Eurohypnum leptothallum)、北地扭口藓(Barbula fallax)和宽叶真藓(Bryum funkii)三种苔藓植物重要值之和为65.96%。这说明这三种垫状丛集生长的矮小型苔藓植物较为适应干旱、缺土的喀斯特石漠环境, 为喀斯特石漠化地区的常见苔藓植物种类。这主要与这三种苔藓植物具有较强的耐旱能力, 以及在能在少土石面聚集丛生有关。

表1 喀斯特重度石漠化地区苔藓植物的重要值Tab. 1 Names and importance values of karst severe rocky desertification area
4.2优势苔藓植物种间总体联结性
选取重要值≥0.5%的14种苔藓植物优势种进行种间总体联结性计算, 计算VR值(VR=0.72<1),说明这14种苔藓植物种间表现出负联结。对以上计算结果, 引入统计量W检测VR偏离1的显著性。W= 106.01<X20.95,148=121, 说明14个优势种之间在总体上表现出显著的负联结。这反应出喀斯特重度石漠化地区苔藓植物群落正处于非常不稳定的初级群落阶段, 易受环境影响, 环境的微小变化便可导致苔藓植物群落组成发生变化。这可能是由于苔藓植物作为拓荒植物首先入侵喀斯特重度石漠化地区, 很少有其他高等植物生长, 物种单一, 群落简单。随着时间的推移, 由于苔藓植物的长期生长改善了喀斯特石漠化地区干旱、少土的环境, 让其他原来不适应此环境的高等植物也有了立足之地, 物种趋于多样化, 群落也逐渐趋于稳定, 最终改善喀斯特环境。
4.3X2检验
图1为X2检验半矩阵图, 14个优势种组成的91个种对中, 正关联35个种对, 占总种对数的38.46%,其中显著、极显著正关联5对, 占总种对数5.50%;负关联56个种对, 占总种对数的61.54%, 其中显著、极显著负关联7对, 占总种对数7.69%; 正负关联比为0.62。由此可见, 负关联种对数明显比正关联总对数多, 表明14种苔藓植物种群的生态位分离,各类苔藓植物的生态适应性各不相同, 种对间两两同时出现的比率不高, 种群处于发展演替中, 不够稳定, 容易受到外界因素干扰而发生波动, 这与所研究的喀斯特重度石漠化地区正处于演替的最初级阶段吻合。种对间极显著和显著关联只有12对, 占总种对数的13.19%, 比例并不高, 这说明物种间相互依赖程度不高, 虽然呈现一定的联结性, 但仍表现出相对独立的分布格局。
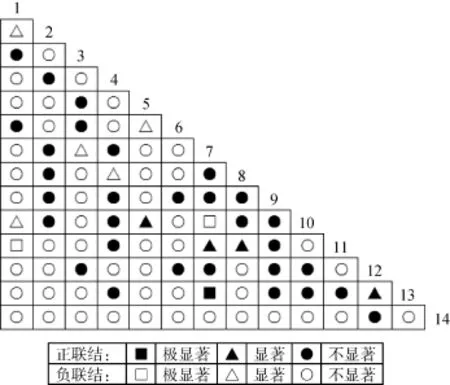
图1 14个优势种对间X2检验的半矩阵图Fig. 1 The semi-matrix of X2–text between 14 dominant species
4.4Pearson相关系数和 Spearman秩相关系数
种对的Pearson相关系数和 Spearman秩相关系数是反映两个物种线性关系的重要指标[21], 其处理的是定量数据, 反映两个物种同时出现的可能程度。
图2为14个种对间的Pearson相关系数和 Spearman秩相关系数的半矩阵图, 3种检验比较结果见表2。
Pearson相关系数检验中正相关的种对数为24,占总种对数的26.37%; 负关联种对数为67, 占总种对数的73.63%, 正负关联比为0.36。Spearman秩相关系数检验中正相关的种对数为36, 占总种对数的39.56%; 负关联种对数为55, 占总种对数的60.44%,正负关联比为0.65。负关联种对数明显高于正关联种对数, 这说明绝大多数物种间的关系并不紧密,种间联结较为松散, 彼此间的生长过程中影响较小,种间独立性较强, 受外界干扰时易发生波动。Pearson相关系数检验和Spearman秩相关系数检验的结果与X2检验的结果相符合
Spearman秩相关系数检验中, 显著、极显著关联占总种对数的17.59%, 高出X2检验(13.19%)和Pearson相关系数检验(8.79%), 这说明Spearman秩相关系数检验比X2检验和Pearson相关系数检验更加敏感, 这与王琳和张金屯[4]的研究相似。
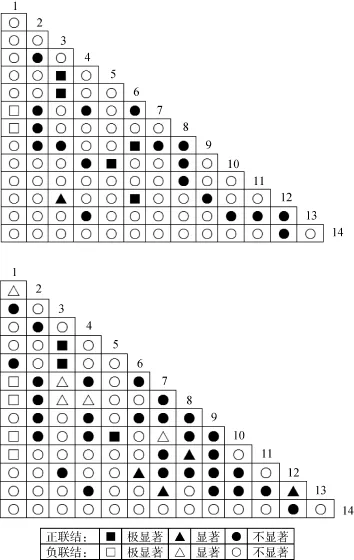
图2 14个优势种对间Pearson相关系数和Spearman秩相关系数半矩阵图Fig. 2 The semi-matrix of Spearman’s rank correlation coefficients and Pearson’s correlation coefficients between 14 dominant species
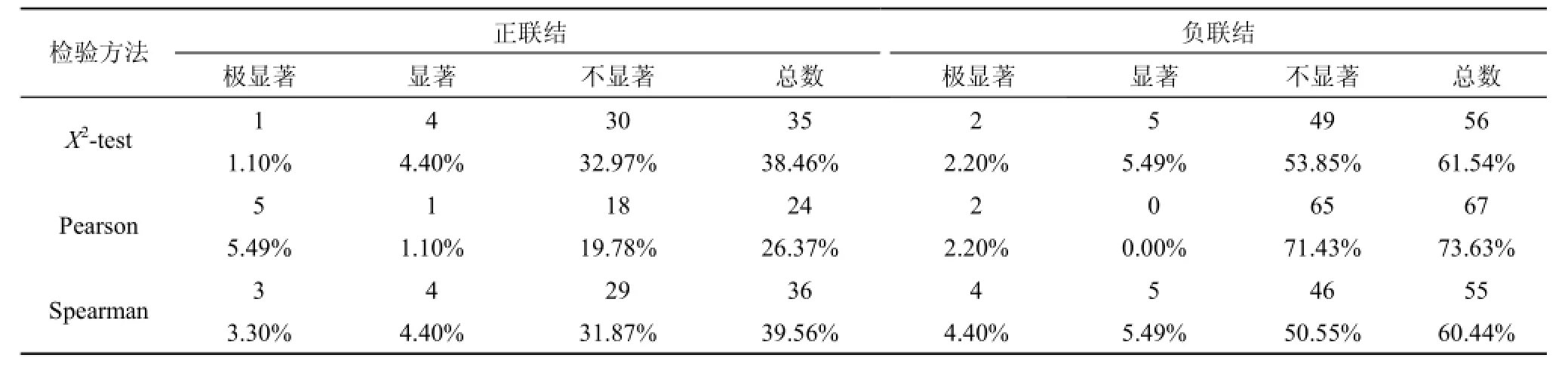
表2 X2检验, Pearson相关系数检验, Spearman秩相关系数检验对比Tab. 2 Comparison of X2–text,Pearson,s correlation coefficients and Spearman,s rank correlation coefficients
由图2可知, 在Pearson相关系数和 Spearman秩相关系数的半矩阵图中, 可以得出大体一致的结论: 宽叶真藓与黑扭口藓、宽叶真藓与卷叶湿地藓、黑扭口藓与芽胞湿地藓三个种对间均为极显著正相关, 这说明两物种间有相近的生物学特性, 以及相似的生态位, 表现出一定的生态位重叠。据观察, 这些苔藓植物均喜好生长在背阴面的岩石上。郭泉水的研究表明[22], 2个物种呈正相关, 则表明当一个种的多度在一个样方中增加时, 则另一个种的多度在同一个样方中也相应地增加; 2个物种间呈负相关,则表明一个种的多度在一个样方中增加, 而另一个种的多度在同一个样方中减少。美灰藓和毛口藓、阔叶毛口藓、芽胞湿地藓和丛生真藓均表现出极显著负相关, 而且美灰藓与其它苔藓植物也基本表现出负关联, 这说明美灰藓与其它苔藓植物的生态学特征不一致, 美灰藓为交织型苔藓植物, 一般为单种纯群落生长, 不与其它苔藓植物混合生长。
4.5生态种组划分
生态种组的划分可以反映物种的生物学特性差异, 并且在一定程度上可以反映物种对不同微环境的适应。图3为14种优势苔藓植物的主成分分析(PCA)二维排序图, 图中箭头长度表示物种多度(重要值)的大小, 两物种射线之间夹角代表其物种间相关性, 夹角的余弦值在数值上等于两物种间的相关系数, 夹角越小相关性越大。PCA结果显示, 第一和第二排序轴的特征值为40.65%和15.40%, 为主成分轴。
群落中生态位及生活习性相同的可以划为一个生态种组[23], 根据以上关于优势苔藓植物种间关联性和相关性研究结果, 以及对种群数据所作PCA排序分析, 可以将14种优势苔藓植物划分为2个生态种组。
第1生态组包括宽叶真藓、黑扭口藓、卷叶湿地藓、芽胞湿地藓和小石藓原变种共7个种, 该组苔藓植物多分布在背阴岩石表面上, 喜好生长在背阴岩壁或遮荫的岩面上, 喜阴。
第2生态组包括北地扭口藓、银叶真藓、毛口藓、阔叶毛口藓、钙生灰藓和丛生真藓共6个种。该组苔藓植物多分布于干燥、向阳且土壤稀薄的岩面上, 喜阳, 耐干旱。

图3 14种优势苔藓植物PCA二维排序图Fig. 3 Two dimensional graph of PCA ordination of 14 dominant bryophytes species
美灰藓和羽枝青藓均为交织型苔藓植物, 多为纯群落, 不喜与其他种植物混生, 一般为岩面薄土生长, 故与其他苔藓植物相关性不高。
5 讨论
5.1X2检验, Pearson相关系数检验和Spearman秩相关系数检验
X2检验, Pearson相关系数检验和Spearman秩相关系数检验的结果具有一致性, 但也存在一定的差异性。X2检验只能作出定性判断, Pearson相关系数检验和Spearman秩相关系数检验是基于重要值、盖度或频率等定量数据的检验方法, 属于定量检测方法[24]。Pearson相关系数检验要求物种数据服从正态分布, 但大多数物种难以满足此条件, 这极大限制了相关系数检验的应用范围[4]。而Spearman秩相关系数检验属于非参数检验, 它并不要求知道物种服从何种分布, 因此应用更为灵活和方便[4]。在本研究中, Spearman秩相关系数检验的显著、极显著关联占总种对数的17.59%, 明显高出X2检验(13.19%)和Pearson相关系数检验(8.79%), 体现出Spearman秩相关系数检验的敏感性, 灵活性。
5.2种间关联与群落演替
理论上讲, 正负关联比越高, 群落结构越趋于稳定, 多物种越趋于稳定, 多物种可以稳定共存,群落达到与环境条件相适应的稳定阶段[25]。有研究表明, 群落优势种种群种对间正负关联比>1, 体现出群落处于演替阶段的后期[26]; 反之, 这说明群落处于演替阶段的前期或中期[27]。本研究中, 三种检验方法对喀斯特严重石漠化地区优势苔藓植物种间关联的检验结果均表现出正负关联比<1的情况, 这说明该区域苔藓植物群落处于演替的前期或中期。事实上, 该区域为重度石漠化区域, 未见其他高等植物, 仅有苔藓植物及少量草本植物生长, 正处于演替的初级阶段, 群落并不稳定, 种间独立性强,外界的干扰极可能造成环境巨大的波动, 对植物群落造成毁灭性损害。
6 结论
在喀斯特重度石漠化地区共记录苔藓植物5科11属30种, 呈现出美灰藓、北地扭口藓和宽叶真藓三种苔藓植物所主导的格局(重要值之和为65.96%)。这说明这三种垫状丛集生长的矮小型苔藓植物较为适应干旱、缺土的喀斯特石漠环境, 为喀斯特石漠化地区的常见苔藓植物种类。
选取14种优势苔藓植物作种间总体联结性、结果表明喀斯特重度石漠化地区14种优势苔藓植物间在总体上表现出显著的负联结, 表现出苔藓植物群落正处于非常不稳定的初级群落阶段。X2检验, Pearson相关系数检验和Spearman秩相关系数检验分析, 正负关联比分别为0.62、0.36和0.65, 均<1,这均表明苔藓植物生态为发生分离, 各类苔藓植物的生态适应性各不相同。显著、极显著关联占总种对数的比例分别为13.19%、8.79%和17.59%, 均不高, 这表明种间联结较为松散, 彼此间的生长过程中影响较小, 种间独立性较强, 受外界干扰时易发生波动。
宽叶真藓与黑扭口藓、宽叶真藓与卷叶湿地藓、黑扭口藓与芽胞湿地藓三个种对间均为极显著正相关, 美灰藓和毛口藓、阔叶毛口藓、芽胞湿地藓和丛生真藓均表现出极显著负相关,
对14种苔藓植物做主成分分析(PCA), 可将苔藓植物分为2个生态组, 1组为喜阴的宽叶真藓、黑扭口藓、卷叶湿地藓、芽胞湿地藓和小石藓原变种;另1组为喜阳的北地扭口藓、银叶真藓、毛口藓、阔叶毛口藓、钙生灰藓和丛生真藓。
[1] 张璐, 苏志尧, 陈北光. 中国特有植物—华南五针松群落的种间关系[J]. 生态学报, 2006, 26(4): 1063–1072.
[2] 梁士楚. 云贵鹅耳枥群落演替中乔木树种间协变的研究[J]. 广西植物, 1995, 15(4): 335–339.
[3] GREIG-SMIT H P. Quantitative plant ecology [M]. Oxford: Black well Science Publications, 1983: 105–112.
[4] 王琳, 张金屯. 历山山地草甸优势种的种间关联和相关分析[J]. 西北植物学报, 2004, 24(8): 1435–1440.
[5] 刘喆, 岳明. 太白山独叶草及其伴生种的种间关联研究[J]. 武汉植物学研究, 2007, 25(5): 445–450.
[6] 郭忠玲, 马元丹, 郑金萍, 等. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究[J].应用生态学报, 2004, 15(11): 2013–2018.
[7] 谢湉, 巨天珍, 师贺雄, 等. 甘肃省小陇山濒危珍稀植物白皮松群落种间关联[J]. 生态学杂志, 2010, 29(3): 448–453.
[8] 赵彩莉, 张峰, 庞春花, 等. 反枝苋群落优势种的种间关联性分析[J]. 植物研究, 2013, 33(4): 454–460.
[9] 胡文强, 黄世能, 李家湘, 等. 南岭石坑崆山顶矮林乔木优势种群的种间关联性[J]. 生态学杂志, 2013, 32(10): 2665–2671.
[10] 张桂萍, 张峰, 茹文明. 山西绵山植被木本植物优势种群种间关联[J]. 生态学杂志, 2006, 25(3): 295–298.
[11] 李军峰, 王智慧, 张朝晖. 喀斯特石漠化山区苔藓多样性及水土保持研究[J]. 环境科学研究, 2013, 26(7): 759–764.
[12] 张朝晖, 王智慧, 祝安. 黄果树喀斯持洞穴群苔藓植物岩溶的初步研究[J]. 中国岩溶, 1996, 15(3): 224–232.
[13] ZHANG Zhaohui. Bryoflora and some speleothems of karst caves in Guizhou S. W. Chin[C]//Switzerland, Proceedinds of 12thInternational Congress of Speleology, Vo1. 3; Biospeology, 1997: 297–300.
[14] 张朝晖, 陈家宽. 黔中瀑布水生苔藓植物区系及其生物喀斯特沉积生态类型研究[J]. 中国岩溶, 2007, 26(2): 170–177.
[15] 崴嵬, 刘方, 向仰州. 贵阳市花溪麦坪煤矿废弃地植被调查与分析[J]. 贵州大学学报(自然科学版), 2009, 26(2): 132–135.
[16] 高谦. 中国苔藓志 [M]. 北京: 科学出版社, 1996: 1–277.
[17] 黎兴江. 中国苔藓志. 第四卷[M]. 北京: 科学出版社, 2006: 1–238.
[18] 胡人亮, 王幼芳. 中国苔藓志. 第七卷[M]. 北京: 科学出版社, 2005: 1–243.
[19] 吴鹏程, 贾渝. 中国苔藓志. 第八卷[M]. 北京: 科学出版社, 2011: 1–493.
[20] 贾渝, 何思. 中国生物物种名录. 第一卷[M]. 北京: 科学出版社, 2013: 1–417.
[21] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004: 98–109.
[22] 郭泉水, 包奋强, 王祥福, 等. 2008. 三尖杉所属群落优势乔木树种种间关系[J]. 林业科学研究, 21(5): 662–668.
[23] 张先平, 王孟本, 张伟锋, 等. 庞泉沟国家自然保护区森林群落木本植物种间关系的分析[J]. 植物研究, 2007, 20(3): 351–355.
[24] 简敏菲, 刘琪, 朱笃. 九连山常绿阔叶林乔木优势种群的种间关联性分析植物[J]. 生态学报, 2009, 33(4): 672–680.
[25] 周先叶, 王伯荪, 李鸣光, 等. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J]. 植物生态学报, 2000, 24(3): 332–339.
[26] 金则新. 浙江天台山常绿阔叶林优势种群结构及种间联结性研究[J]. 广西植物, 2002, 22(3): 203–208.
[27] 胡贝娟, 张钦弟, 张摇玲, 等. 山西太岳山连翘群落优势种种间关系[J]. 生态学杂志, 2013, 32(4): 845–851.
Interspecific association among dominant species of bryophytes in karst severe rocky desertification area
YIN Shengfeng1, WANG Zhihui2, ZHANG Zhaohui1*
1.Key Laboratory for Information System of Mountainous Areas and Protection of Ecological Environment of Guizhou Provincial,Guizhou Normal University,Guiyang, Guizhou550001,China
2.School of Life Sciences,Guizhou Normal University,Guiyang, Guizhou550001,China
Bryophytes occupy a very important position in the karst ecological construction. It plays an important ecological function in maintaining the ecological biodiversity. We selected karst areas in Huaxi District, Guiyang, Guizhou Province for the study and investigated the distribution of bryophytes.X2-test, Pearson’s correlation coefficients, Spearman’s rank correlation coefficients and PCA were applied in the analysis. The results showed that 30 species belonged to 11 genus and 5 families. From the correlation analysis of 14 dominant bryophytes, a significantly negative correlation between the species of bryophytes was found, which indicated the bryophyte communities were in the primary stage of succession; the ratio of the positive and negative correlation (less than 1) suggested that the bryophytes niches were separated; the PCA results showed that two ecological groups were divided, which were bryophytes located in shady rocks and apricus rock surface.
karst severe rocky desertification; bryophytes; interspecific association; PCA
10.14108/j.cnki.1008-8873.2016.05.013
Q948.1
A
1008-8873(2016)05-090-06
殷声锋, 王智慧, 张朝晖. 喀斯特重度石漠化地区苔藓植物优势种的种间关联性研究[J]. 生态科学, 2016, 35(5): 90-95.
YIN Shengfeng, WANG Zhihui, ZHANG Zhaohui. Interspecific association among dominant species of bryophytes in karst severe rocky desertification area[J]. Ecological Science, 2016, 35(5): 90-95.
2015-06-19;
2015-10-11
国家自然科学基金项目(31160042); 贵州省国际科技合作项目(黔科合外G字2013-7016号)
殷声锋(1988—), 男, 福建南平人, 在读硕士, 主要从事环境生态学研究, E-mail: yinshengfeng042@163.com
*通信作者: 张朝晖, 男, 博士, 教授, 主要从事环境生态学研究, E-mail: academiclife@126.com
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
长江科学院院报(2023年8期)2023-08-29 07:05:22
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
内蒙古林业调查设计(2021年2期)2021-06-29 02:38:12
阅读(低年级)(2020年8期)2020-11-06 06:23:19
乡村地理(2019年2期)2019-11-16 08:49:32
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
中国水土保持(2018年11期)2018-11-06 11:52:38
学术论坛(2016年5期)2016-05-17 05:44:46
杂草学报(2015年2期)2016-01-04 14:57:55
