
半干旱风沙区野生山里红种群特征研究
2016-05-11 01:44栗生枝
防护林科技 2016年4期
关键词:种群
栗生枝
(辽宁省林业调查规划院,辽宁 沈阳 110122)
半干旱风沙区野生山里红种群特征研究
栗生枝
(辽宁省林业调查规划院,辽宁 沈阳 110122)
摘要辽宁省彰武地区属半干旱风沙区,生态环境恶劣,野生山里红是该地区的主要树种之一,对当地生态环境的影响有着举足轻重的作用。因此,对野生山里红进行合理的保护及开发利用十分必要。通过野外调查和室内分析相结合的方法,对辽宁彰武地区野生山里红种群分布格局以及群落特征进行了研究。结果表明:(1)辽宁彰武地区野生山里红种群空间分布格局为聚集分布,且聚集度高;(2)辽宁彰武地区野生山里红群落物种丰富度指数、多样性指数以及均匀度指数均偏低,该地区物种类型较为单一,且分布不均匀,多样性小;(3)辽宁彰武地区野生山里红群落中,杨树重要值最大,为绝对优势树种,野生山里红和榆树为该群落的次优势树种。
关键词半干旱地区;野生山里红;种群;分布格局;群落特征
1研究区基本现状
彰武县地形地貌属辽西丘陵与辽河平原过渡地带,东西部为丘陵区,北部属科尔沁沙地南缘,中、南部为松辽平原坡水地,本次调查是以彰武实验林场为研究区域进行调查取样,该林场位于辽宁省彰武县阿尔乡镇境内,与内蒙古科左后旗相毗邻。地理坐标122°30′57″—122°32′42″ E,42°47′58″—42°48′60″ N,东西长约2.3 km,南北宽约1.8 km,规划面积2 300 hm2。研究区内有流动沙丘、半流动沙丘、固定沙丘、风蚀沟(风蚀平洼地)、丘间低地。
2调查方法
2.1取样方法
全面踏查的基础上,根据地形特征和物种分布状况等,在样地内先从东向西而后由南向北选取样方,一共选取样方32个。每个样方的选取尽可能满足典型而又能够反映样地整体大环境的要求,具有一定的代表性。之后对每个样方内所有木本植物进行调查。每个样方面积为50 m×50 m。对样方内乔木的种类、数量以及胸径进行测量并记录。
2.2计算方法
2.2.1空间分布格局植物种群空间分布格局是指在种群水平上,种群内个体在一定范围空间的分布状况。它是植物生物学特性、中间关系以及各种环境因素综合作用的结果[3-5]。

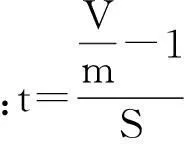
其集群强度用丛生指数(I)、负二项式参数(K)来度量,其具体的计算方法如下:
丛生指数(I):I=(V/m)-1;
负二项参数(K):K=m2/(V-m),K值愈小,聚集度越大[7]。
2.2.2物种多样性物种丰富度是以样地内出现的物种数来衡量,用Margalef指数来衡量;物种多样性用Shannon-Wiener指数来表示和衡量;群落的均匀度表征了群落中各个种的多度或重要值的均匀程度,用Pielou指数来表示。各项指标的计算见下述有关公式:
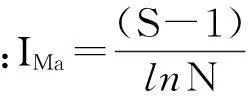
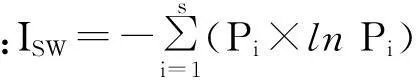

式中:S为物种数目;N为物种的个体总数;Pi=Ni/N,Ni指样方中第i种物种的个体数,并且∑Ni=N[8,9]。
2.2.3重要值重要值是用综合数值表示植物物种在群落中的相对重要性。
针对乔木而言:重要值=(相对多度+相对频度+相对显著度)/3
频度是指一个种在所做的全部样方中出现的频率,相对频度指某种在全部样方中的频度与所有种频度和之比。
相对频度=(该种的频度/所有种的频度总和)×100%
显著度(优势度):指样方内某种植物的胸高断面积除以样地面积。
相对显著度(相对优势度)=(样方中该种个体胸高断面积和/样方中全部个体胸高断面积总和)×100%
密度(D)=某样方内某种植物的个体数/样方面积
相对密度(RD)=(某种植物的密度/全部植物的总密度)×100% =(某种植物的个体数/全部植物的个体数)×100%[10]
3结果与分析
3.1野生山里红种群空间分布格局
通过计算种群空间分布格局指数——方差/均值率以及统计量t值,丛生指数和负二项指数,运算结果如表1。

表1 野生山里红种群空间分布格局检验
由表1可见,山里红种群分布格局表现为聚集分布,而且其负二项参数K值较小,而丛生指数I值较大,说明野生山里红种群聚集强度高,反映出种群趋于成斑块甚至连续的集群。该种群特征是种群个体成群、成簇,成块地密集分布,各群的大小,群间的距离,群内个体的密度都不相等,但各群内部多是随机分布。聚集分布是绝大多数自然种群的稳定分布格局。
种群分布格局的形成是物种与环境长期相互适应和相互作用的结果,一方面决定于物种的生态生物特性和种间竞争等生态学过程,另一方面与群落环境密切相关。半干旱风沙区野生山里红呈集群分布是外因和内因共同作用的结果。首先,野生山里红适应性强,在沙丘地带也能较好生长。据考察,过去彰武地区野生山里红在沙丘地带分布较多,如今野生山里红的分布多集中在沙丘地带深处。据了解这主要是由于近年来开荒种地、砍伐当柴及大量盲目采种所造成的。这也导致了如今野生山里红呈聚集分布的状态。其次,山里红多分布于沙丘阴坡中上部,为浅根系植物,其根系水平延伸可达几米至十几米[1]。因此,该地区的野生山里红多成群分布,所以常在母树周围形成幼树的聚集分布。半干旱风沙区野生山里红分布极其不均,在所选32个样方中有15个样方中没有野生山里红种群,而且有的样方中野生山里红只有零星几株分布(如图1所示)一方面是由于随着年龄的增长,种群对光照、养分、水分的需求增大,种内、种间竞争激烈,导致自疏和他疏,使种群聚集强度下降,也导致了分布不均。另一方面与当地的气候以及土壤条件有关。彰武县气候干燥,降水量较少,蒸发量较大,加上冬季气温低,土质瘠薄,多为沙丘且流动性较大等不利因素导致野生山里红生长困难,分布不均。
3.2植物多样性特征及影响因子分析
由于当地气候条件恶劣,土壤瘠薄,因此乔木种类较少。本次调查所选样地中共有乔木11种,分别为榆树、山里红、杨树、桑树、黄檗、桃叶卫矛、旱柳、鼠李、山杏、樟子松以及五角枫等。计算Margalef丰富度指数(IMa)、Shannon-Wiener多样性指数(ISW)以及Pielou均匀性系数(E),所得结果如表2所示。

表2 样地的植物多样性特征
由表2中结果可以看出,样地的物种丰富度指数以及多样性指数均比较低,说明样地内物种单一,且数量较少,因此物种多样性指数也偏低。样地的均匀性系数也较低,如表2所示,说明样地内物种分布不均匀。
这3个指数均偏低的主要原因在于半干旱风沙地区恶劣的气候以及土壤条件。首先,该地区以前为草原,但是由于各种气候以及人为因素,如今草原退化,荒漠化严重,土壤上层浮沙厚积,沙丘流动性大,土壤贫瘠,因此该地区多为对环境适应性较强的树种,如樟子松、杨树等。很多植物受土壤条件限制在此地无法生长。因此,该地区植物种类较少。其次,该沙区常年为有风天气,尤以春季有风次数最多,而且风速较大。彰武地区地处科尔沁沙地南缘,该地区地表土层疏松,一旦遇到风,风害与沙害同时发生,风蚀沙害是当地典型的恶劣天气。在沙地及退化耕地上,干旱年份遇到大风则风沙危害更严重,更易发生沙尘天气。而风蚀沙害对植物的正常生长有很大的影响,以樟子松为例,樟子松2年生裸根苗在沙化土地上造林时偶尔在背风的周围有灌木和草本做掩护的低洼地能看见成活的樟子松幼苗,冬季不防寒也能越冬;樟子松裸根苗在沙地上造林时基本全部死亡;迎风口处中幼龄樟子松的针叶往往枯萎,而单株幼树往往整株死亡。同样,阔叶树新萌发的嫩芽、嫩叶被风蚀沙割后正常生长也会受到影响[11]。第三,彰武地区年平均气温7.2 ℃,极端最高气温37.4 ℃,最低气温-30.4 ℃。冬季气温低,植物容易发生冻害,不利于植物生长。第四,彰武地区年均降水量为510.3 mm,蒸发量为1 831.5 mm。蒸发量远远大于降水量,而且随着全球气候变暖,温室效应显著,使得该地区干旱年份数量增加。从而导致该地区气候干燥,空气平均相对湿度61%,最大相对湿度78%,最小相对湿度48%。同时,土壤的含水量低,2006年伏旱,0~100 cm土层土壤平均湿度为2.15%,2007年伏旱和秋旱同时发生,0~100 cm土层土壤平均湿度仅为1.5%~2.0%。所有干旱年都不同程度地减小了树木生长量,不同程度地降低了造林成活率,严重的可能导致树木提前落叶,树势衰弱,感染病害,更严重的可导致树木干枯死亡。调查发现,连年干旱对植被危害最大,也反映了当地土壤调节水分能力极弱。地下水位下降使林木利用土壤水分受到很大限制,影响其健康生长。第五,本次调查的区域地势不平,因此样方设在山丘底部以及背风处时,样方内植被分布较多,而设在坡顶以及迎风处时植被稀少。还有一部分样方由于距离樟子松防护林带较近,因此乔木数量较多,种类较丰富,而且长势较好。在地势平坦处植被分布较均匀。从总体上来看,样地内乔木数量大小不一(如图2),整体分布不均,且差异明显。因此,均匀性系数较低。
3.3结构特征
样地内乔木由于树龄大小差异较大,因此树高差距明显。乔木层高0.5~11 m,最矮的为去年栽植的樟子松林,最高的为人工栽植的速生杨。树木胸径断面积在0.000 314~0.321 536 m2,胸径断面积最小的是去年栽植的樟子松,最大的是野生黄檗。在整个群落中杨树重要值为1.127 008,远高于其他树种,占绝对优势,为该群落优势种;次优势树种为野生山里红和榆树,重要值分别为0.582 096和0.499 783,此外按重要值大小依次为大果榆、樟子松、五角枫、旱柳、鼠李、黄檗、桑树、山杏和桃叶卫矛(如表3)。
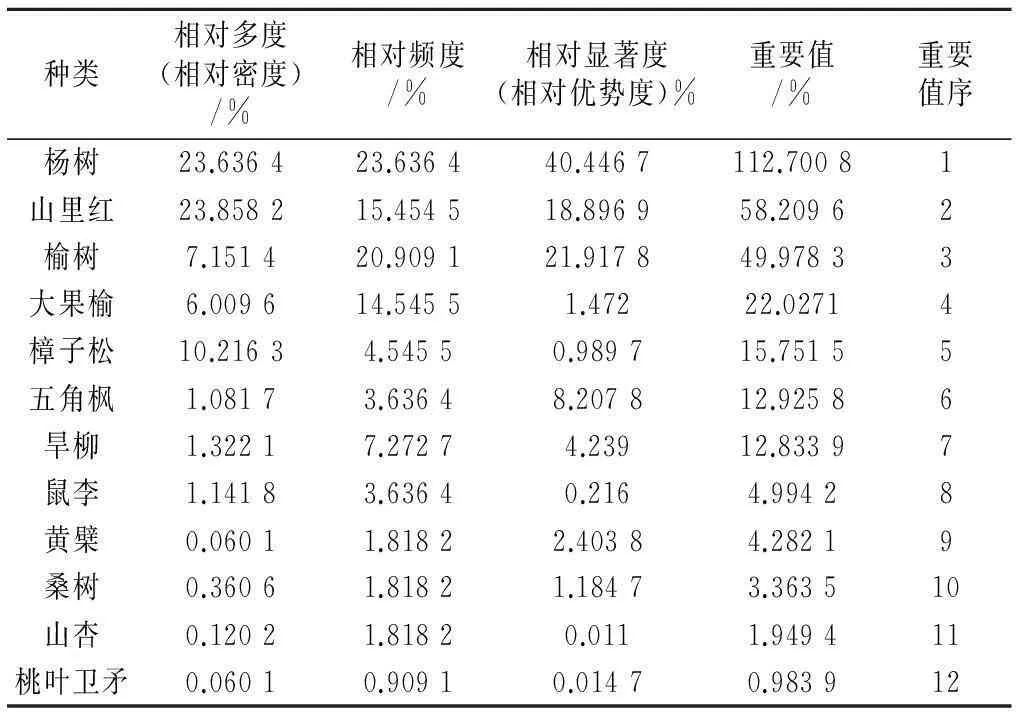
表3 乔木层植物重要值
样地紧邻彰北防护林,因此样地内的杨树多数为人工栽植的防护林树种,数量较多,且长势较好。有一小部分为野生的加杨。样地内速生杨和加杨的总数为809株(图3)。而样地内的野生山里红和榆树都是野生树种,在数量上与杨树相差较多,导致杨树的相对多度和相对显著度远高于野生山里红和榆树,重要值远大于野生山里红和榆树,成为该群落的优势树种。由表3中还可以看出,山里红的相对多度较榆树大,但是其相对频度和相对显著度较榆树小。主要是由于样地内榆树总数量为119株,而山里红多为丛生,聚集分布,总数量为397株(见图3),因此在数量上山里红远大于榆树,使得其相对密度大于榆树的相对密度。但是从总体分布以及树胸径断面积来看,榆树分布较广泛,且树龄大,胸径断面积大,而山里红多聚集分布,树龄大小不一,母树高大,但其旁丛生的多为矮小植株,胸径断面积远小于榆树(如图4)。因此从总体上来看,山里红的重要值大于榆树,但是两者相差不多,同为本地区的次优势树种。样地内的樟子松虽然也是人工栽植的防风固沙树种,但是其数量远远小于杨树的数量,仅有170株。而且样地内的樟子松多为近2年新栽植的,树龄小,株高多在1~2 m,少有大树,其胸径断面积也较小(如图4),因此樟子松的重要值远小于杨树、榆树和野生山里红的重要值。样地内大果榆的数量为100株,多以灌木丛状分布,因此其重要值较大,为22.027 1。样方内旱柳数量为22株,五角枫18株,鼠李19株(见图3),数量远远小于樟子松,在样方内零星分布,其相对多度值较低,但是由于其相对频度以及相对优势度值较高,因此,其重要值与樟子松相差不多。黄檗、桑树、山杏以及桃叶卫矛在样地内分布稀少,多为零星几株。桑树总数为6株,山杏2株,而黄檗和桃叶卫矛只有一株。因此其重要值远远小于其他树种。
野生山里红是该样地的次优势树种,说明野生山里红在该样地群落中具有生长优势。由于当地气候以及土壤条件的限制,多数乔木在此地不能正常生长,致使当地乔木种类较少,而且分布范围窄,数量少。野生山里红为乡土树种,其适应性极强,在沙丘地带也可以正常生长。因此在整个乔木层中就凸显出野生山里红的生长优势,成为当地的次优势树种。
野生山里红虽然是该群落的次优势树种,但是从植物自身来看,其优势度并不明显,其主要原因还是在于当地恶劣的气候以及贫瘠的土壤。本次野外调查发现,当地土地荒漠化程度十分严重,而且气候恶劣,如若不及时防治土地沙漠化,该地区的生态系统将受到严重破坏,植被生长受到严重影响,导致植被在此地无法生长,进而导致此地风沙更大,形成恶性循环,因此对该地区进行土地荒漠化治理十分必要。
4结论与讨论
野生山里红种群空间分布格局呈聚集分布,且聚集程度高。呈聚集分布的原因主要是由野生山里红种的生物学特性以及半干旱风沙区的环境因素所决定。
野生山里红群落物种丰富度指数、多样性指数以及均匀度指数均偏低,该地区物种类型较为单一,且分布不均匀、多样性小。半干旱风沙区气候、土壤条件恶劣,植物种类及数量稀少,因此该群落的物种多样性低。
野生山里红群落中,杨树重要值最大,为绝对优势树种,野生山里红和榆树为该群落的次优势树种。杨树多为人工栽植的防护林树种,数量多,因此优势明显。而野生山里红和榆树是天然散生林,数量远小于杨树,优势不明显,因此是该群落的次优势树种。
关于种群空间格局的成因,不同的物种具有不同的原因,一般认为小规模的聚集是由植物本身的特征所决定的。大规模的聚集主要是植物从原分布中心扩散,由环境因子的影响所决定的。影响种群分布格局的环境因子主要有土壤、水分、土壤养分、环境的异质性、动物、火烧等[12]。
由于彰武地区特殊的气候以及土壤条件,环境对野生山里红种群的空间格局的形成也起到了关键作用。由环境条件引起的种内种间竞争以及气候土壤等外在环境条件对野生山里红植物种群生长的影响导致了彰武地区野生山里红的种群空间分布格局为聚集分布,环境因素对野生山里红种群空间分布格局的影响程度在一定程度上起到了决定性作用。本文认为,该地区由气候、土壤等外在条件对山里红空间格局的影响大于由环境引起的种内种间竞争对其格局的影响,因此,在考察野生山里红种群空间分布格局的影响因素时不仅要对种群本身的生物学特性进行调查研究,对其气候、土壤等外在环境条件对种群空间格局的影响进行调查研究十分必要。
采用样方法取样进行种群分布格局研究存在一些缺点。对于一个聚集分布的种群,不同的样方大小会得到完全不同的结果。假如样方大小远大于聚集分布的斑块大小,测定结果会是一个均匀分布的格局;而样方面积太小,取样资料会得出一个随机分布格局;只有当样方面积与斑块大小接近以至相等时,测定结果才可能为聚集型分布。在对一个种群分布格局未加研究以前,并不清楚其斑块大小,也就不可能确定相应样方大小。而且一个聚集分布产生的原因很多。斑块大小也不一致。因此,在森林群落中为种群分布格局研究进行样方取样时,可选取不同大小的样方进行取样,最后综合不同大小的样方数据,确定种群空间分布格局。但是由于本次调查不仅要研究山里红的种群空间分布格局,还对其群落特征进行研究,因此样方大小无法及时调整。
彰武县阿尔乡北部属科尔沁沙地南缘,属草原与沙丘过渡带,为稀疏草原地带。沙漠化过程是一个物种多样性衰退的过程,首先,沙漠化导致特有种的绝灭,其次为稀有种和普通种[13]。本次调查的样地中特有种以及稀有种已经绝灭,普通种的种类和数量也很少。物种多样性的调查旨在保护生物多样性,恢复群落的生物多样性,恢复生态系统的内部结构和功能。因此,对该地区进行物种多样性的调查十分必要。
通过本次调查说明,该地区的物种类型少、物种丰富度低,而且多样性指数偏低,植被覆盖率低,荒漠化程度高,生态系统破坏严重。虽然部分地区已经实现退耕还林,且已配置了防护林带,但是其作用并不明显,新栽植的樟子松林由于各种自然和人为因素成活率低,部分退耕还林地区并没有真正地种植树木来防风固沙。因此,该地区应加大防风固沙、退耕还林力度。在已经退耕还林的地方应栽植速生杨、樟子松等防护林树种,切实做到退耕还林。同时应该加强防风固沙、退耕还林的监督力度,减少人为因素对防护林树种的影响,提高成活率,让防护林起到防风固沙的作用。
参考文献:
[1] 李作轩,张德学,陆胜友,等.彰武沙丘地带野生山里红资源考察报告[J].辽宁果树,1986(4):26-27
[2] 胡秀娟,程积民,杨晓梅,等.黄土高原子午岭林区辽东栎种群分布格局及群落特征研究[J].西北农报,2010, 25(5):1-6
[3] 李鹏远,程积民,万惠娥,等.云雾山自然保护区优势植物种群分布格局的分形特征[J].生态学报, 2008,28(10):5161-5165
[4] Qu Z X,Wu Y S,Wang H X,et al.Plant Ecology(Second Edition)[M].Beijing:Higher Education Press,1983:190-193
[5] Zu Y G,Ma K M.Fractal theory and ecology[M]//Li Bo.A course of modern Ecology.Beijing:Science Press,1995.65-72
[6] 刘富强,王延平,杨阳,等.黄河三角洲柽柳种群空间分布格局研究[J].西北林学院学报,2009,24(3): 7-11
[7] 张金屯.模糊数学及其应用[J].生态学报,1992,12(4):325-331
[8] 刘俊霞,袁登胜,袁秀英.花棒群落物种多样性及其种群空间分布格局的研究[J].西北林学院学报,2008, 23(5):55-59
[9] 杨澄.桥山天然栋林树种多样性及生态位分析[J].西北林学院学报,1998,13(4):28-32
[10] 程瑞梅,等.河南宝天曼锐齿栎林群落学特征[J].生态学杂志,1999,19(4):25-30
[11] 韩辉,王国晨,白雪峰,等.彰武沙区异常恶劣气候对植被恢复的影响与对策[J].防护林科技,2001(6): 95-97
[12] 郑元润.大青沟森林植物群落主要木本植物种群分布格局及动态研究[J].植物学通报,1998,15(6): 52-58
[13] 尚占环,姚爱兴.草原生物多样性研究及其保护[J].宁夏农学院学报,2002,23(2):70-75
Population Characteristics of WildCrataeguspinnatifidain Semiarid Sandy Area
Li Shengzhi
(Forest Inventory and Planning Institute, Liaoning Province, Shenyang 110122,China)
AbstractZhangwu areas in Liaoning Province is a semi-arid sandy area, with harsh environment;wild Crataegus pinnatifida is one of the main tree species in the region and plays an important role in local ecological environment.Through field investigation & laboratory analysis methods,population distribution patterns and community characteristics of wild Crataegus pinnatifida in Zhangwu areas were studied. Result shows that : (1) Spatial distribution pattern of wild Crataegus pinnatifida population in Zhangwu area is aggregated distribution, with higher aggregation-degree; (2) Richness, diversity and evenness index of species for wild Crataegus pinnatifida in Zhangwu area of Liaoning Province are all low, the type of species in the region is single, and the distribution is uneven, diversity being small;(3) Among wild Crataegus pinnatifida communities in Zhangwu area of Liaoning Province,the important value of poplar are the largest, being the absolute dominant tree species;wild Crataegus pinnatifida and Ulmus pumila are subdominant tree species for this community.
Key wordssemiarid area;wild Crataegus pinnatifida ;population;distribution pattern;community characteristics
中图分类号:S718.54
文献标识码:A
doi:10.13601/j.issn.1005-5215.2016.04.019
作者简介:栗生枝(1987-),男,辽宁沈阳人,大学,工程师,现从事林业资源调查和研究工作,Email:85799984@qq.com
收稿日期:2016-01-06
文章编号:1005-5215(2016)04-0055-05
猜你喜欢
今日农业(2022年15期)2022-09-20
湖南电力(2021年1期)2021-04-13
甘肃林业(2019年4期)2019-10-08
红土地(2018年7期)2018-09-26
生物学教学(2018年3期)2018-08-08
数学物理学报(2018年1期)2018-03-26
中学生物学(2018年8期)2018-03-01
贺州学院学报(2015年1期)2015-02-28
当代畜禽养殖业(2014年2期)2014-08-22
当代畜禽养殖业(2014年10期)2014-02-27
