
坡向对青藏高原东北缘高寒草甸植被构成和养分特征的影响
2016-05-10 09:36:14徐长林
草业学报 2016年4期
徐长林
(甘肃农业大学草业学院,甘肃 兰州 730070)
坡向对青藏高原东北缘高寒草甸植被构成和养分特征的影响
徐长林*
(甘肃农业大学草业学院,甘肃 兰州 730070)
摘要:坡向是重要地形因子之一,通过影响光照、温度及土壤养分等进而影响群落植物种的组成和分布。本研究通过对青藏高原东北缘不同坡向高寒草甸草地的植被构成和养分特征进行了分析,揭示坡向对植物群落组成和空间分布形成的影响。结果表明,1) 草地群落优势种和亚优势种构成,阳坡以禾本科的赖草、西北针茅和洽草等为主,阴坡以莎草科的矮嵩草和线叶嵩草及豆科的阴山扁宿豆等为主;2)草层高度、植物物种数和α多样性指数为阴坡>阳坡,β多样性指数在坡向之间无显著差异;3) 阳坡和阴坡草地植物生活型以地面芽和地下芽为主,分别占草地群落总物种数的71.0%和89.0%,1/2年生及地上芽植物为阳坡>阴坡,地面芽植物为阴坡>阳坡,地下芽植物在坡向之间无显著差异;4) 草地群落生物量构成,阳坡为禾本科(78.3%)>莎草科(8.3%)>菊科(6.4%)>其他科(7%),阴坡为莎草科(54.3%)>豆科(16.4%)和禾本科(14.8%)>蓼科(5.3%)和菊科(4.6%)>其他科(4.0%);除菊科外,所有各科植物生物量在坡向之间均差异显著,地上总生物量在坡向之间无显著差异;5) 0~20 cm土壤N含量为阴坡>阳坡,10~20 cm土壤P、K、Cu、Zn含量为阳坡>阴坡;6) 0~30 cm土温为阳坡>阴坡,0~40 cm土壤含水量阴坡>阳坡。总之,坡向对高寒草甸草地群落植物种构成和土壤理化特性的变化起着重要作用。关键词:高寒草甸;坡向;群落构成;物种数;生活型;生物量;养分
高寒草甸是青藏高原主要植被类型之一,是适应高原隆起与长期低温环境形成的特殊产物[1],约占青藏高原可利用草地面积的35%。它不仅是支撑高原畜牧业发展、维系地方民族经济发展的物质基础,而且对长江中下游平原及黄河、长江流域地下水、地表水的丰歉,防止风、旱、涝、沙尘暴、水土流失等自然灾害的发生,起着不可替代的屏障作用[2]。
地形是一些生态过程形成的基本因素,也是环境时空异质性的重要来源,其主要通过影响非生物资源,如光辐射、温湿度及土壤养分等的分配格局,而进一步影响植物群落的组成和分布。坡向作为一个最重要地形因子,不同坡向因光照、水分、温度、土壤等差异,而使植物物种组成、物种丰富度、植物生活型和生物量等不同[2-9];通常,阳坡受较多太阳辐射影响,而具较高的温度,分布耐旱的植物群落,阴坡则生长喜阴或湿生植物群落。同时,土壤是植物生长的物质基础,与植物的生长发育和分布等存在着重要关系。研究表明,土壤养分和水分的有效性,对植物生态系统的生产力具有很强的控制作用[10],土壤含水量和养分决定草地的质量及生产力[11];而且植物生物量与土壤养分相互影响、相互作用,土壤养分及其空间异质性影响植物群落的结构、功能及动态[12-13]。李昆和陈玉德[14]对云南干热河谷人工林地水分研究发现,阳坡的土壤水分显著低于阴坡。一般情况下,阴坡土壤的通透性、结构及持水性都比阳坡的好。邱波等[15]对青藏高原东北部高寒草甸不同生境植物物种a和β多样性分布特性进行研究发现,当生境的变化方向为阴坡-滩地-阳坡时,植物a多样性指数呈降低变化,而其β多样性呈先减少后增加再减少的变化方式。Gong等[16]对内蒙古锡林河盆地坡向对植物物种组成及生产力的影响研究表明,阴坡的物种多样性和生产力比阳坡高,阴坡生产力的主要限制因素是土壤可利用养分浓度,而阳坡生产力的主要限制性因素是土壤水分。王婧[17]研究了延河流域环境梯度对植物群落结构变化的影响,发现因阴坡湿度大,植被发育良好,而使阴坡植物的Shannon-Wiener多样性指数和相对丰富度指数比阳坡高。因此,草地群落植物种的构成和物种多样性的形成,与其所处的生境、气候及水热条件密切相关。
综合分析地形尤其是坡向因子对植被和养分的影响,认为虽然阴坡-阳坡的尺度范围较小,但由于光照、水分及温度等诸多因子受坡向的影响,使不同坡向之间的植被构成和种群结构、物种多样性、土壤养分及生态系统功能等产生较大差异,从而其完整地展示了一个生境梯度。坡向是导致植物物种多样性组成和结构特征产生差异的间接环境因子,它通过改变光照、温度、水分和土壤等生态因子而对生物多样性、植物生长发育、生产力以及生态系统功能等产生重要影响。因此,研究植物构成及其分布与坡向之间的关系,明晰阴阳坡植物生存策略,对深入揭示生境对植物群落空间格局形成的作用机制具有重要理论意义。而青藏高原高寒草甸不同生境(特别是坡向)下植物功能性状和土壤养分分布的报道相对较少,且青藏高原地区主要坡向是阴坡和阳坡。基于此,本研究以青藏高原东北缘草地群落为研究对象,比较两种生境(阳坡和阴坡)的植被功能构成和养分特征,为高寒草甸的管理及其生态系统服务功能评价提供科学依据。
1材料与方法
1.1研究区概况
研究区位于青藏高原东缘的甘肃省天祝县抓喜秀龙乡的高寒草甸上,地理坐标为E 102°40′-102°47′,N 37°11′-37°14′,海拔2960 m。年均气温-0.1℃,1月均温-18.3℃,7月均温12.7℃,>0℃年积温 1380℃;年降水量416 mm,无绝对无霜期,仅分冷热两季,土壤类型为亚高山草甸土。草地主要植物种有赖草(Leymussecalimus)、矮嵩草(Kobresiahumilis)、线叶嵩草(Kobresiacapillifolia)、西北针茅(Stipakrylovii)、紫花针茅(Stipapurpurea)、珠芽蓼(Polygonumviviparum)、阴山扁宿豆(Medicagorutheniavar.inschanica)和球花蒿(Artemisiasmithii)等。
1.2样地设置
在实地调查基础上,依据坡向不同,于2012年5月中下旬,在研究区植被构成明显不同的阴坡(shady,S)和阳坡(terrace,T)两种生境,多年放牧牦牛的冷季草地利用率为80%~90%的高寒草甸上,各设置面积为0.3~0.5 hm2的样地3块为试验样地,样地间距约1 km,共设置6个样地。
1.3测定指标和方法
1.3.1植物群落特征2012年7月下旬-8月下旬,在各样地植被相对均一的草地上,分别随机设置0.5 m×0.5 m的样方15个,调查各样方内所有植物种的高度、密度、盖度。然后齐地刈割收获地上生物量。将收获的地上生物量混合后,均分为2份。其中,1份于70℃下烘干测干重后,用于植物养分分析;另1份的地上生物量先按绿色物质和死物质分开,再将绿色物质按不同植物种分开,装入信封袋,置于70℃烘箱内烘干称干重。按公式IV=(RC+RH+RB)/3计算植物种重要值,式中,IV(important value)为植物种重要值,RC、RH和RB分别为植物种的相对盖度、相对高度和相对生物量。
利用样方调查数据,按公式DGl=S/lnA[18],计算物种α多样性指数DGl(Gleason指数);式中,S为群落中的物种数目,A为样方面积。并按公式βW=(S/ma)-1[18],计算物种β多样性指数βW(Whittaker指数);式中,S为研究系统中观测物种总数,ma为各样方或样本的平均物种数。
此外,亦利用调查样方数据,统计草地植物群落生活型谱数据。
1.3.2土壤和牧草养分在地上植被特征测定同期,分别在各样地内设置3条200 m的样线,等距离设置10个点,用直径3.5 cm的土钻采集0~10 cm和 10~20 cm深度的土样,剔除土壤中的根系、石块等杂物后,置于标记好的自封袋内,带回实验室阴干,然后过0.15 mm筛,测定土壤养分。
P和N含量测定采用凯氏定氮法;K、Na、Mg、Ca、Mn、Zn、Cu和Fe含量测定采用原子吸收光谱法。具体分析方法见杨胜[19]和鲁如坤[20]及《草原生态化学实验指导书》[21]。所有指标数据均换算为干物质基础数据。
1.3.3土壤温度和水分2012年7月末,在晴朗天气,连续3 d早上9-12时,采用金属插入式温度计,测定各样地0~10 cm、10~20 cm、20~30 cm和30~40 cm土层温度。同时,2012年5月末、7月末和9月末,在晴朗天气,分别在各样地内布置3条200 m的样线,等距离设置10个点,用直径3.5 cm的土钻分别采集各样地0~10 cm、10~20 cm、20~30 cm和30~40 cm层次的土样,置于70℃干燥箱中烘干称重,测定土壤含水量。
1.4数据分析
用Excel进行数据处理及制图。SPSS 16.0软件的One-Way ANOVA分析坡向对植物群落特征和生物量及土、草养分和水分特征的影响(F-检验),并将数据表示为均值±标准误(standard error of mean)。
2结果与分析
2.1草地主要植物种重要值
由表1可知,阳坡草地群落构成以禾本科植物为主,其优势种为赖草和西北针茅,亚优势种为洽草,主要伴生种有扁穗冰草、矮嵩草、早熟禾、大籽蒿、蓝花韭、阴山扁宿豆、冷蒿、线叶嵩草和二裂委陵菜等;阴坡草地群落构成以莎草科植物为主,其优势种为矮嵩草,亚优势种为线叶嵩草和阴山扁宿豆,主要伴生种有异针茅(Stipaaliena)、珠芽蓼、洽草、球花蒿、垂穗披碱草、苔草、高山嵩草、瑞香狼毒、蓝花韭和甘肃棘豆等。这说明,两种生境的草地群落主要植物种重要值差异较大。

表1 不同坡向草地植物群落主要植物种重要值(IV≥0.01)
2.2草地植物群落特征
由表2可知,坡向对草地植物群落高度、盖度和物种数以及物种α多样性指数DGl均具极显著影响。阳坡草地植物群落的高度显著高于阴坡(P<0.001),为后者的1.2倍;阴坡草地植物群落的盖度、物种数和α多样性指数DGl显著高于阳坡(P<0.01和P<0.001),前者分别为后者的1.1,1.8和1.6倍,但其β多样性指数βW在坡向之间差异不显著(P>0.05)。说明坡向对草地植物物种丰富度构成影响大,而对草地群落的物种替代速率影响小。
草地群落的植物种生活型谱构成结果显示,阳坡生境中,1或2年生(annual/biennial,A/B)及地上芽(chamaephyte, CP)植物比例均极显著和显著高于阴坡(P<0.001和P<0.05),且前者分别为后者的5.6和1.5倍;阴坡生境中,地面芽植物比例极显著高于阳坡(P<0.05),且前者为后者的1.4倍;而地下芽植物比例在阳坡和阴坡之间差异不显著(P>0.05)(图1)。两种坡向高寒草甸群落植物构成以地面芽(hemicryptophytes,HP)和地下芽(geophytes,GP)为主;阳坡和阴坡的HP分别为41.9%和57.1%,GP分别为29.0%和31.4%,二者约占草地群落总物种数的71%和89%,且其1/2年生植物以十字花科为主,地上芽植物以菊科植物为主。这说明,不同坡向之间植物种的构成不同,可导致其植被群落特征及植物种生活型谱的差异。

表2 不同坡向高寒草甸草地植物群落特征
注:NS,*, ** 和***分别表示差异不显著(P>0.05)以及在P<0.05、P<0.01和P<0.001水平上差异显著,下同。
Note: NS, *, ** and *** mean no significant difference, and significant differences at 0.05, 0.01 and 0.001 levels, respectively. The same below.
2.3草地植物生物量构成
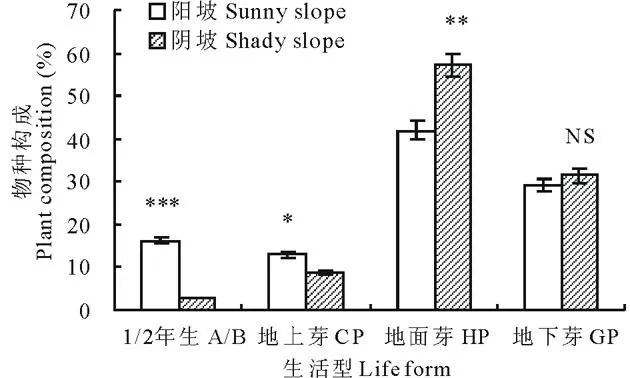
图1 不同坡向植物种生活型谱构成Fig.1 The life-form spectrum of plants of different slopes NS,*, ** 和***分别表示差异不显著(P>0.05)以及在P<0.05、P<0.01和P<0.001水平上差异显著,下同。NS, *, ** and *** mean no significant difference, and significant difference at 0.05, 0.01 and 0.001 levels, respectively. The same below.
由表3可以看出,阳坡草地群落绿色物质量主要由禾本科植物构成(78.3%),其次为莎草科(8.3%)和菊科(6.4%),其他科植物仅约占7%;阴坡草地群落绿色物质量主要由莎草科植物构成(54.3%),其次为豆科(16.4%)、禾本科(14.8%)、蓼科(5.3%)和菊科(4.6%),其他科植物仅约占4.0%。这与主要植物种重要值结果一致(表1),说明草地群落生物量主要由少量植物种构成。
两种生境草地植物生物量构成除菊科、活物质和总生物量无显著差异(P>0.05)外,所有各科植物的生物量均在坡向之间差异极显著(P<0.01或P<0.001)(表3)。其中,阳坡植物群落中禾本科、百合科、蔷薇科、唇形科、藜科、十字花科、鸢尾科和紫草科的生物量极显著高于阴坡(P<0.01或P<0.001),且前者的禾本科、百合科和蔷薇科植物的生物量分别是后者的4.8,3.2,4.0倍;阴坡植物群落中莎草科、豆科、蓼科、大戟科、虎耳草科、龙胆科、毛茛科、茜草科、瑞香科、石竹科和玄参科以及死物质(立枯体+凋落物)生物量极显著高于阳坡(P<0.01),且前者的莎草科和豆科植物以及死物质生物量分别为后者的6.9, 9.5和1.6倍。这表明,阳坡适于耐旱的禾草和杂类草植物生长,阴坡适于喜阴或湿生的莎草、菊科、豆科和其他植物生长。
植物群落生物量构成结果还显示,蓼科、大戟科、虎耳草科、茜草科、瑞香科和玄参科植物在阳坡中的生物量约为0,唇形科、藜科、十字花科、鸢尾科和紫草科在阴坡中的生物量约为0(表3)。这表明蓼科的珠芽蓼和圆穗蓼(Polygonummacrophyllum)、大戟科的狼毒大戟(Euphorbiafischeriana)、虎耳草科的细叉梅花草(Parnassiaoreophila)、茜草科的蓬子菜(Galiumverum)、瑞香科的狼毒和玄参科的兰石草(Lanceatibetica)等不适于阳坡生长,唇形科的异叶青兰(Dracocephalumheterophyllum)、藜科的灰藜(Chenopodiumalbum)、十字花科的播娘蒿(Descurainiasophia)、荠菜(Capsellabursa-pastoris)和独行菜(Lepidiumapetalum)、鸢尾科的马蔺(Irislacteavar.chinensis)和紫草科的微孔草(Microulasikkimensis)等不适于阴坡生长。
2.4草地植物和土壤养分
草地植物群落矿质养分含量结果显示,牧草P、Na、K、Fe、Cu、Zn等元素含量在两种生境之间无显著差异(P>0.05);阴坡植物群落的Ca和Mg元素含量极显著高于阳坡(P<0.001),前者分别为后者的1.8和1.9倍;牧草Mn元素含量则为阳坡极显著高于阴坡(P<0.01),前者为后者的1.3倍(表4)。表明,高寒草甸牧草Ca、Mg和Mn元素含量对坡向响应敏感。
0~10 cm和 10~20 cm土壤矿质养分含量结果显示,除 0~10 cm土壤N含量阴坡比阳坡高外(P<0.05),两种生境 0~10 cm土壤P、Na、K、Mg、Fe、Cu、Mn和Zn含量无显著差异(P>0.05);两种生境 10~20 cm土壤Na、Mg、Fe和Mn含量无显著差异(P>0.05),阴坡 10~20 cm土壤N含量极显著高于阳坡(P<0.01),阳坡 10~20 cm土壤Cu和Zn含量极显著高于阴坡(P<0.01),且阳坡10~20 cm土壤P和K含量显著高于阴坡(表4)。说明坡向对高寒草甸草地土壤N含量影响大,土壤矿质养分含量因土层深度不同而有差异。

表3 不同坡向植物群落生物量构成
2.5草地土壤温度和水分
由图2可以看出, 0~10 cm、 10~20 cm和20~30 cm土壤温度均为阳坡>阴坡(P<0.05或P<0.01),土壤30~40 cm温度在阳坡和阴坡之间无显著差异(P>0.05),且土壤越深,温度越低(P<0.05)。土壤水分结果显示,5月 0~10 cm和30~40 cm的土壤含水量为阴坡>阳坡(P<0.05或P<0.01),阳坡和阴坡 10~20 cm和20~30 cm土壤含水量无显著差异(P>0.05);7和9月的 0~10 cm、10~20 cm、20~30 cm 和30~40 cm土壤含水量均为阴坡>阳坡(P<0.05或P<0.01或P<0.001);土壤含水量随土壤深度的加深呈显著降低变化(P<0.05)。可见,一般阳坡土壤温度比阴坡高,阴坡土壤含水量高于阳坡。
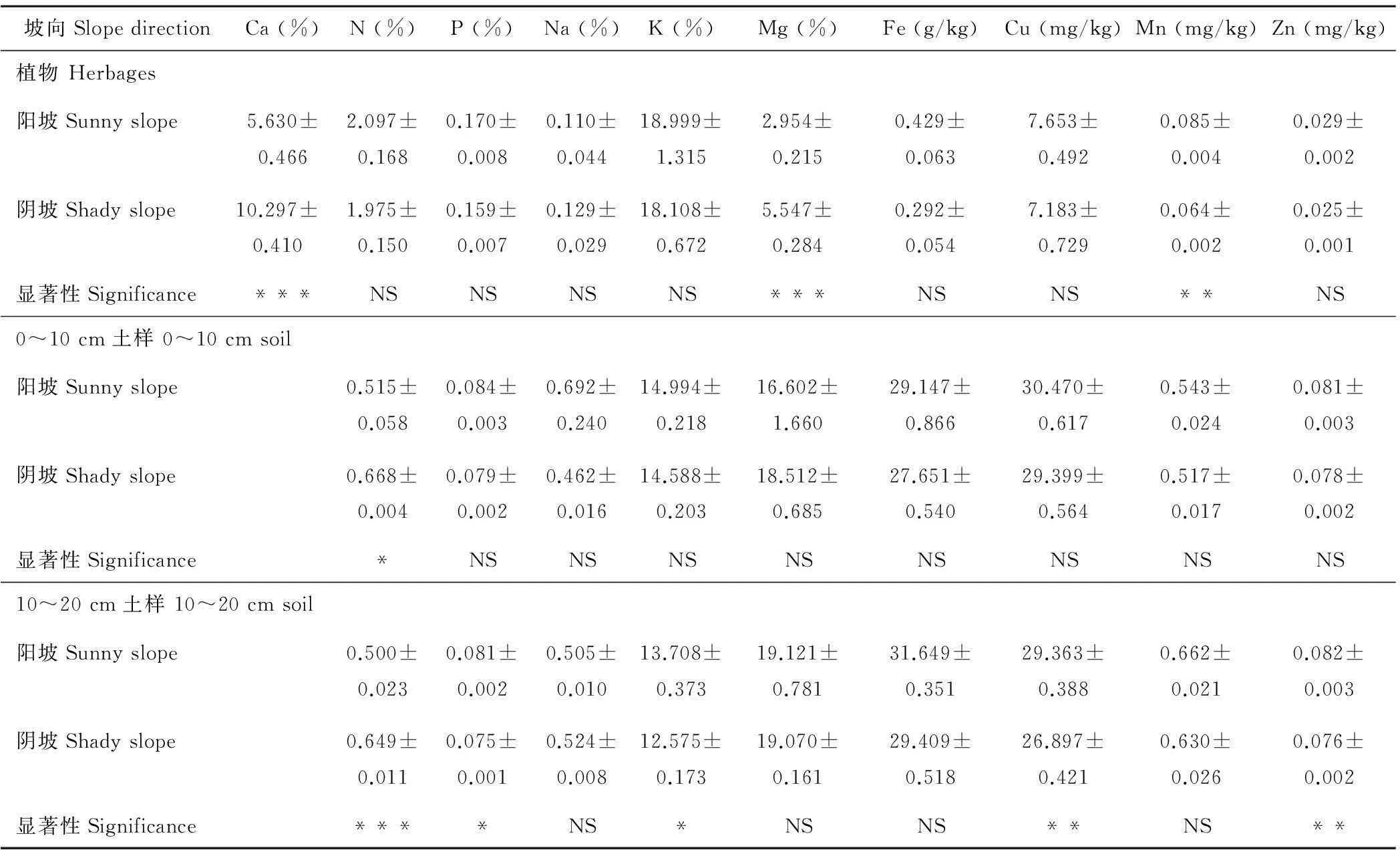
表4 不同坡向高寒草甸草地植物和土壤养分
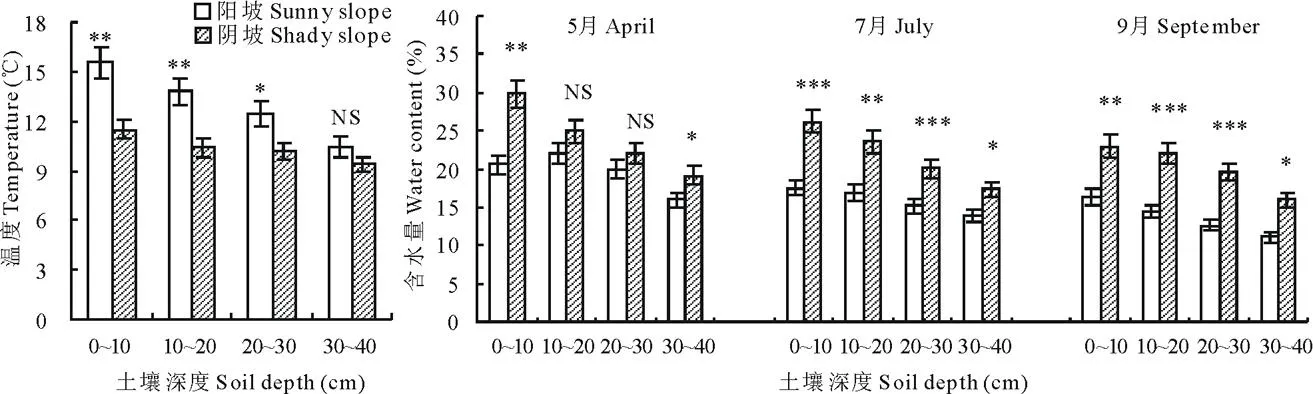
图2 不同坡向土壤温度和水分Fig.2 Temperature and moisture in soils of different slopes
3讨论与结论
坡向通过影响光照、土壤水分及养分分配等而影响植物群落的构成和分布。自然状态下,植物群落物种组成是物种对环境适应的结果,在漫长的演化过程中,植物群落形成特定的结构,以及物种与环境之间的相互依存关系,是植物对生态气候条件长期适应的综合反映。物种间、生物与环境的紧密关系将随时、空尺度变化而变化,它表现出的多样性与生产力的关系,能真实地反映物种多样性在长期进化过程中对生产力的长期稳定效应。本研究中阳坡和阴坡草地植物高度及禾草和莎草等生物量比例差异很大,是由于阳坡草地群落以个体植株较大的赖草、西北针茅、洽草和扁穗冰草等禾本科植物为主构成,阴坡草地群落以个体植株较小的莎草科植物矮嵩草和线叶嵩草及豆科植物阴山扁宿豆等为主构成。形成这种现象的原因与草地表面上热量、水分的分布密切相关。从而,禾草是阳坡的优势功能群,莎草和杂草是阴坡的优势功能群,阴阳坡优势功能群的改变,也是资源利用策略改变的一种反应。
植物物种丰富度表示群落所含物种的多寡,β多样性表示不同生境群落之间物种组成的相异性及共有种的差异。本研究草地群落植物物种数和α多样性指数均为阴坡>阳坡,与以往研究结果一致[15,17,22];而本研究草地的β多样性指数βW在坡向之间差异不显著,这与以往研究存在分异,可能是本研究植被调查中每重复样方数多,而使群落中出现的植物种总数较多导致。草地群落植物种的构成和物种多样性的形成,与其所处的生境、气候及水热条件密切相关[16-17]。本研究阳坡光照强,土壤温度高,水分蒸发大,而有利于深根型、耐旱喜阳环境的长根茎型禾草如赖草、针茅、冰草和洽草等植物的生长和分蘖;阴坡由于光照弱,土壤温度低,土壤蒸发量小湿度大,以及土壤和植被的高低不平分布而形成诸多微环境,这有利于喜阴湿环境植物如嵩草和苔草等分蘖和生长及物种多样性的形成,从而使本研究草地植物构成及其生活型因坡向不同而差异很大。据此认为,草地植物群落分布格局是物种对环境适应的结果,不同坡向因生境条件的差异,可导致物种多样性的差异。
太阳辐射是地球生态系统最主要和最直接的能量来源,也是地表温度的主要来源。土壤水分变化又通过改变土壤热特性影响土壤温度,土壤含水量越高,土壤容积比热容越大,土壤增温越缓慢。因此,土壤水分和温度之间具有密切联系,二者相互作用,相互影响。在阴坡坡面上,太阳辐射越小,使得草地潜在蒸散就越小;反之,阳坡辐射大,使得草地潜在蒸散就越大,导致草地土壤水分含量低,在不同坡向上也是这样的,土壤含水量的变化都是由太阳辐射的差异引起的。事实上,坡向内地表自然形态因素决定了太阳辐射量的吸收;草地地被覆盖物、土壤湿度、利用类型等地被覆盖因素共同作用决定了能量吸收、转化和放射太阳辐射热量。所以一般阳坡土壤温度比阴坡高。
虽然一个地区主要植被类型由气候条件决定,但群落植物的具体分布还更多受土壤因子的影响[23-25]。土壤中的水分和营养条件决定植物的生长状况,土壤水分和养分因子之间存在密切的关系[26]。一般来讲,土壤水分既影响养分在土壤中的转化和运移,又影响植物吸收水分与利用养分的情况,而养分又通过影响植物根系形态与生长而影响植物的水分情况。本研究除土壤TN含量为阴坡>阳坡以及 10~20 cm土壤P、K、Cu、Zn含量为阳坡高于阴坡外,坡向对其他各土壤养分指标影响小。但多数研究认为,阴坡土壤养分一般比阳坡好[16,27]。这可能是本研究阳坡和阴坡两种生境各有优缺点,阳坡虽然样地平坦、温、光条件优越,植物养分转化快,但土壤含水量低,植被稀疏;阴坡虽然土壤水分含量高,但受光照时间长短不同,使阴坡的太阳光线与坡面的夹角比平地小,进而阴坡吸收的太阳辐射能量比阳坡少,从而阴坡温度比阳坡低。本研究阴坡温、光条件差,坡度大,从而其植物分解慢,养分易流失,而使两种生境养分差异小。此外,水热条件变化所引起的物种选择、资源竞争、生境的变化也是影响物种多样性的重要因素。研究表明,短期增温对高寒植被有正效应,而温度持续增高则对植被产生负效应[28]。张庆等[29]对内蒙古草原短花针茅(Stipabreviflora)群落的DCCA分析结果反映出群落结构格局主要受到热量、水分梯度变化的影响。因此,水热条件也是影响植物群落变化的主要因素,本研究中也得出类似结果。
总之,坡向作为重要的地形因子,通过改变光照、温度,间接影响土壤水分的含量和分布,进而对生物多样性、植被类型和生产力产生重要影响。土壤养分的差异,也对植物群落的物种组成、生物量和群落演替产生重要作用。坡向影响生境的不同,进而导致植物群落随坡向而变化。
References:
[1]Li Y N, Zhao L, Zhao X Q,etal. Effects of a 5-years mimic temperature increase to the structure and productivity ofKobresiahumilismeadow. Acta Agrestia Sinica, 2004, 12(3): 236-239.
[2]Long R J. Functions of ecosystem in the Tibetan grassland. Science & Technology Review, 2007, 25(9): 26-28.
[3]Wen C J. Influence of the relief on conditions of water and heat in the Hengduan Mountains region. Mountain Research, 1989, 7(1): 65-73.
[4]Chinese Academy of Sciences. Tibet Vegetation[M]. Beijing: Science Press, 1988: 277-292.
[5]Yu Y D, Liu L H, Zhang J H. Vegetation regionalization of the Hengduan Mountains region. Mountain Research, 1989, 7(1): 47-55.
[6]He Z J. The vertical distribution zone of forest soils on the north slope of Dabashan mountain. Shaanxi Forest Science and Technology, 1993, (2): 6-8.
[7]Busby J R, Bliss L C, Hamilton C D. Microclimate control of growth rates and habitats of the boreal forest mosses,TomenthypnumnitensandHylocomiumsplendens. Ecological Monographs, 1978, 48: 95-110.
[8]Holz I, Gradstein S R, Kappelle H M. Bryophyte diversity, microhabitat differentiation, and distribution of life forms in costa rican upper montaneQuercusforest. Bryologist, 2002, 105: 334-348.
[9]Sadler K D, Bradfield G E. Microscale distribution patterns of terrestrial bryophytes in a subalpine forest: the use of logistic regression as an interpretive tool. Community Ecology, 2000, 1: 57-64.
[10]Hao W F, Liang Z S, Han R L,etal. Study on the relationship between soil properties and different type vegetation in Loess Plateau. Acta Botanica Boreali-Occidentalia Sinica, 2002, 22(6): 1545-1550.
[11]Wang Q, Jia D J. Soil moisture and nutrient affect grassland production in alpine meadow. Chinese Journal of Grassland, 1990, 3: 6-13.
[12]Du F, Liang Z S, Xu X X,etal. Community biomass of abandoned farmland and its effects on soil nutrition in the Loess hilly region of Northern Shaanxi, China. Acta Ecologica Sinica, 2007, 27(5): 1673-1684.
[13]Trangmar B B, Yost R S, Wade M K,etal. Spatial variation of soil properties and rice yield on recently cleared land. Soil Science Society of America Journal, 1987, 51: 668-674.
[14]Li K, Chen Y D. Study on input of water regime and soil water content in the plantation of the Yuanmou dry and hot river valley. Forest Research, 1995, 8(6): 651-657.
[15]Qiu B, Ren Q J, Du G Z,etal. Study on α diversity and β diversity of plant community of different habitats in alpine meadow. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(4): 655-661.
[16]Gong X, Brueck H, Giese K M,etal. Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia, China. Journal of Arid Environments, 2008, 72(4): 483-493.
[17]Wang J. The Response of Vegetation Community Structure to Environment Gradients in Yanhe River Basin[D]. Yangling:Northwest A&F University, 2011.
[18]Qin L. Statistical Ecology[M]. Beijing: China Forestry Publishing House, 2009: 75-77.
[19]Yang S. Feed and Feed Quality Detection Technology[M]. Beijing: China Agricultural University Press, 1999.
[20]Lu R K. Soil Agricultural Chemical Analysis Method[M]. Beijing: China Agricultural Science and Technology Press, 2000.
[21]Gansu Agricultural University. Grassland Ecological Chemical Experiment Instruction[M]. Beijing: Agriculture Press, 1987.
[22]Nie Y Y, Li X E, Wang G. Variation mode of diversity and diversity of plant community of different habitat gradients from south-facing slope to north-facing slope and its relation with different environmental factors. Journal of Lanzhou University(Natural Sciences), 2010, 46(6): 73-78.
[23]Huang C Y. Agrology[M]. Beijing: China Agriculture Press, 2000: 3-9.
[24]Wang C T, Long R J, Wang Q J,etal. Distribution of organic matter, nitrogen and phosphorus along an altitude gradient and productivity change and their relationships with environmental factors in the Alpine meadow. Acta Prataculturae Sinica, 2005, 14(4): 15-20.
[25]Dong Q M, Zhao X Q, Li Q Y,etal. Responses of soil nutrient contents and water to stocking rates for yaks inKobreciaparvealpine meadow. Chinese Journal of Soil Science, 2005, 26(4): 493-500.
[26]Lu H, Cong J, Liu X,etal. Plant diversity patterns along altitudinal gradients in alpine meadows in the three river headwater region China. Acta Prataculturae Sinica, 2015, 24(7): 197-204.
[27]Liu Y X, Ma J Z. Feature variations of plant functional traits and environmental factor in south- and north-facing slope. Research Soil and Water Conservation, 2013, 20(1): 102-106.
[28]Xu M H, Xue X. A research on summer vegetation characteristics & short-time responses to experimental warming of alpine meadow in the Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 2013, 33(7): 2071-2083.
[29]Zhang Q, Niu J M, Buyantuyev A,etal. Ecological analysis and classification ofStipabrevifloracommunities in the Inner Mongolia region: the role of environmental factors. Acta Prataculturae Sinica, 2012, 21(1): 83-92.
参考文献:
[1]李英年, 赵亮, 赵新全, 等. 5年模拟增温后矮嵩草草甸群落结构及生产量的变化. 草地学报, 2004, 12(3): 236-239.
[2]龙瑞军. 青藏高原草地生态系统之服务功能. 科技导报, 2007, 25(9): 26-28.
[3]文传甲. 横断山区地形对水热条件的影响. 山地研究, 1989, 7(1): 65-73.
[4]中国科学院. 西藏植被[M]. 北京: 科学出版社, 1988: 277-292.
[5]余有德, 刘伦辉, 张建华. 横断山区植被分区. 山地研究, 1989, 7(1): 47-55.
[6]何忠俊. 大巴山北坡森林土壤垂直带谱. 陕西林业科技, 1993, (2): 6-8.
[10]郝文芳, 梁宗锁, 韩蕊莲, 等. 黄土高原不同植被类型土壤特性与植被生产力关系研究进展. 西北植物学报, 2002, 22(6): 1545-1550.
[11]王钦, 贾笃敬. 高寒草甸土壤水和营养对草地生产的影响. 中国草地学报, 1990, 3: 6-13.
[14]李昆, 陈玉德. 元谋干热河谷人工林地的水分输入与土壤水分研究. 林业科学研究, 1995, 8(6): 651-657.
[15]邱波, 任青吉, 杜国祯, 等. 髙寒草甸不同生境类型植物群落的α及β多样性研究. 西北植物学报, 2004, 24(4): 655-661.
[17]王婧. 延河流域植物群落结构对环境梯度变化的响应[D]. 杨凌: 西北农林科技大学, 2011.
[18]覃林. 统计生态学[M]. 北京: 中国林业出版社, 2009: 75-77.
[19]杨胜. 饲料分析及饲料质量检测技术[M]. 北京: 中国农业大学出版社, 1999.
[20]鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科学技术出版社, 2000.
[21]甘肃农业大学. 草原生态化学实验指导书[M]. 北京: 农业出版社, 1987.
[22]聂莹莹, 李新娥, 王刚. 阳坡-阴坡生境梯度上植物群落α多样性与β多样性的变化模式及与环境因子的关系. 兰州大学学报(自然科学版), 2010, 46(6): 73-78.
[23]黄昌勇. 土壤学[M]. 北京: 中国农业出版社, 2000: 3-9.
[24]王长庭, 龙瑞军, 王启基, 等. 高寒草甸不同海拔梯度土壤有机质氮磷的分布和生产力变化及其与环境因子的关系. 草业学报, 2005, 14(4): 15-20.
[25]董全民, 赵新全, 李青云, 等. 小嵩草高寒草甸的土壤养分因子及水分含量对牦牛放牧率的响应-Ⅱ冬季草场土壤营养因子及水分含量的变化. 土壤通报, 2005, 26(4): 493-500.
[26]卢慧, 丛静, 刘晓, 等. 三江源区高寒草甸植物多样性的海拔分布格局. 草业学报, 2015, 24(7): 197-204.
[27]刘昱霞, 马建祖. 阴阳坡植物功能性状与环境因子的变化特征. 水土保持研究, 2013, 20(1): 102-106.
[28]徐满厚, 薛娴. 青藏高原高寒草甸夏季植被特征及对模拟增温的短期响应. 生态学报, 2013, 33(7): 2071-2083.
[29]张庆, 牛建明, Buyantuyev A, 等.内蒙古短花针茅群落数量分类及环境解释. 草业学报, 2012, 21(1): 83-92.
Variations in vegetation composition and nutrient characteristics related to aspect in an alpine meadow in the northeast margin of the Qinghai-Tibet Plateau
XU Chang-Lin*
CollegeofPrataculturalScience,GansuAgriculturalUniversity,Lanzhou730070,China
Abstract:Aspect is an important topographical factor that affects illumination, temperature, and soil nutrients, all of which affect the composition and distribution of plant species. In this study, we evaluated the effect of aspect on the soil nutrient characteristics and on the composition and spatial distribution of plant communities in an alpine meadow in the northeast margin of the Qinghai-Tibet Plateau. The dominant and subdominant species in grassland communities were Leymus secalinus, Stipa krylovii, and Koeloria cristata (Gramineae) on sunny slopes and Kobresia humilis and Kobresia capillifolia (Cyperaceae) and Melilotoids ruthenica var. inschanica (Leguminosae) on shady slopes. The herbage height, α-diversity index, and number of plant species in grasslands were greater on shady slopes than on sunny slopes, while the β-diversity indexes of grasslands were similar between shady and sunny slopes. The plant life-form spectrum both on sunny and shady slopes mainly consisted of hemicryptophytes and geophytes, accounting for 89% and 71% of the total number of species, respectively. There were more annual and biennial plants and hemicryptophytes on sunny slopes than on shady slopes, but more hemicryptophytes on shady slopes than on sunny slopes. There were similar numbers of geophytes on sunny and shady slopes. The plant families contributing to total biomass in grassland communities on sunny slopes were Gramineae (78.3%)>Cyperaceae (8.3%)>Asteraceae (6.4%)>others (7%); and to grassland communities on shady slopes were Cyperaceae (54.3%)>Leguminosae (16.4%), Gramineae (14.8%)>Polygonaceae (5.3%), Asteraceae (4.6%), and others (4%). For all of the plant families except for the Asteraceae, the contribution to total biomass differed significantly between sunny and shady slopes, but the contribution to total aboveground biomass was similar between sunny and shady slopes. The nitrogen content in the 0-20 cm soil layer was higher on the shady slope than on the sunny slope, while the phosphorus, potassium, copper, and zinc contents in the 10-20 cm soil layer were higher on the sunny slope than on the shady slope. The soil temperature in the 0-30 cm layer was higher on the sunny slope than on the shady slope, whereas the water content in the 0-40 cm soil layer was higher on the shady slope than on the sunny slope. Together, these results show that aspect plays an important role in the composition of vegetation communities and in the physical and chemical properties of soil in alpine meadows.
Key words:alpine meadow; slope; community composition; species number; life-form; biomass; nutrients
*通信作者
Corresponding author.
作者简介:徐长林(1956-),男,甘肃天祝人,副研究员。E-mail: xucl@gsau.edu.cn
基金项目:国家自然基金项目(31460566)和甘肃省农业科技创新项目《草业生态系统教育部重点实验室开放课题(GNCX-2014-27)资助。
*收稿日期:2015-10-15;改回日期:2015-11-30
DOI:10.11686/cyxb2015481
http://cyxb.lzu.edu.cn
徐长林. 坡向对青藏高原东北缘高寒草甸植被构成和养分特征的影响.草业学报, 2016, 25(4): 26-35.
XU Chang-Lin. Variations in vegetation composition and nutrient characteristics related to aspect in an alpine meadow in the northeast margin of the Qinghai-Tibet Plateau. Acta Prataculturae Sinica, 2016, 25(4): 26-35.
猜你喜欢
今日农业(2020年19期)2020-12-14 14:16:52
测绘学报(2019年11期)2019-11-20 01:31:42
成才之路(2017年21期)2017-07-27 20:52:40
安徽农学通报(2017年9期)2017-05-19 23:03:05
现代农业科技(2016年22期)2017-03-24 12:21:57
土壤与作物(2015年3期)2015-12-08 00:46:55
中国科技信息(2015年2期)2015-11-16 08:18:32
学周刊·下旬刊(2015年10期)2015-07-22 12:36:17
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
植物营养与肥料学报(2012年3期)2012-10-26 02:54:52
