
低频电刺激对低氧高二氧化碳大鼠比目鱼肌肌纤维类型的影响
2016-05-06 04:02黄世园蒋贤逊沈洁王小同
温州医科大学学报 2016年3期
关键词:电刺激
黄世园,蒋贤逊,沈洁,王小同
(温州医科大学附属第二医院 脑科、康复中心,浙江 温州 325027)
低频电刺激对低氧高二氧化碳大鼠比目鱼肌肌纤维类型的影响
黄世园,蒋贤逊,沈洁,王小同
(温州医科大学附属第二医院脑科、康复中心,浙江温州325027)
[摘 要]目的:探讨低频电刺激对低氧高二氧化碳模型大鼠比目鱼肌肌纤维类型转变的影响。方法:成年SD大鼠40只随机分成4组:正常组10只,捆绑组10只,造模组10只,低频电刺激组10只。造模组、捆绑组及低频电刺激组大鼠置于低氧高二氧化碳舱内造模,每次持续8 h,总共4周。低频电刺激组在持续造模2周后,用电刺激仪对大鼠进行30 min,30 Hz,1:1循环模式的低频电刺激,低频电刺激维持2周;捆绑组仅做捆绑及贴电极片处理。HE染色检测大鼠比目鱼肌病理改变,免疫组织化学观察比目鱼肌肌纤维类型转变,qRT-PCR检测MHC-I、MHC-IIa/IIx mRNA水平,Western Blot检测比目鱼肌MHC-I、雌激素相关受体γ(ERRγ)、过氧化物酶体增殖物受体β(PPARβ)蛋白含量的变化。结果:相比正常组,捆绑组及造模组大鼠比目鱼肌局部炎症反应明显,肌纤维横截面积明显下降,MHC-I以及PPARβ蛋白含量明显下降,MHC-I mRNA下降,MHCIIa/IIx mRNA含量升高。相比捆绑组或造模组,低频电刺激组大鼠比目鱼肌局部炎症反应减轻,MHC-I、ERRγ 及PPARβ蛋白含量明显增加,MHC-I mRNA含量明显升高,MHC-IIa/IIx mRNA明显下降。结论:低氧高二氧化碳可致大鼠比目鱼肌MHC-I向MHC-IIa/IIx型纤维转变,而低频电刺激可部分逆转低氧高二氧化碳所致的肌纤维类型转变。PPARβ、ERRγ蛋白可能参与其中调节。
[关键词]低氧高碳酸血症;电刺激;大鼠;比目鱼肌;肌球蛋白重链
慢性阻塞性肺疾病(chronic obstructive pulmonary disease,COPD)是一类以持续性进行性气流受限为主要特征的疾病,目前COPD是引起人类死亡的第四大诱因[1]。COPD患者往往伴发多种并发症,其中骨骼肌功能障碍与其高病死率密切相关[2]。COPD患者骨骼肌功能障碍可表现为肌萎缩、肌耐力下降、肌力下降,低氧、氧化应激、废用、系统性炎症等都是潜在的致病因素[3-5]。导致肌耐力下降的直接原因是以I型肌纤维为主的慢肌纤维含量下降,这在COPD患者中比较常见[6]。通过电刺激兴奋骨骼肌的电刺激疗法对于改善COPD患者骨骼肌功能已得到证实[7-8]。不同频率电刺激对肌肉影响不同,低频电刺激可模拟支配慢肌纤维的神经放电模式,能有效提高慢肌耐力[9]。比目鱼肌主要由I型肌纤维组成,其在姿势维持、步行等方面起重要作用。El-Khoury等通过分离低氧模型大鼠比目鱼肌,发现低氧可引起比目鱼肌肌耐力下降[10],但其具体机制并未明了。本研究采用低氧高二氧化碳大鼠模型,探讨低频电刺激对模型大鼠比目鱼肌肌纤维类型的影响。
1 材料和方法
1.1材料
1.1.1实验动物及分组:成年SPF级雄性SD大鼠40只,购自温州医科大学实验动物中心,初始体质量为160~180 g。将大鼠编号后按照随机数表法,分成正常组、捆绑组、造模组、低频电刺激组,每组10只。
1.1.2仪器:动物实验复合模拟舱(潍坊华信氧业有限公司),HANS-200A电刺激仪。
1.2方法
1.2.1低氧高二氧化碳模型建立[11]:将造模组、捆绑组、低频电刺激组大鼠放入氧舱内饲养,饲料及水充足。选用低氧高二氧化碳模式,维持舱内氧气浓度在7%~11%,二氧化碳浓度为5%~6%,舱内温度:22~23 ℃。造模时间为每天8 h,每周6 d,共造模4周,其余时间在正常条件下饲养。对照组置于对照氧舱中饲养(吸入空气)。
1.2.2电刺激方法:低频电刺激组在造模2周后进行电刺激。首先剔去大鼠两侧的腓肠肌上的毛发直至暴露皮肤,再将大鼠固定在捆绑台上,把电极片捆绑在其两侧腓肠肌,保证电极片与腓肠肌处皮肤充分接触。电刺激参数:方波频率30 Hz,工作循环1:1,电流强度通过观察大鼠反应适当调整。每次刺激时间30 min,每周7 d,持续2周。捆绑组大鼠除了不进行电刺激外,其他处理与低频电刺激组相同。
1.2.3骨骼肌分离提取:配置10%的水合氯醛,称量每只大鼠的体质量后,按照0.4 mL/100 g的剂量,经腹腔注射麻醉药,待大鼠进入麻醉状态后,迅速地分离其腓肠肌以及比目鱼肌,解剖位置参考Armstrong的研究[12]。分离后的组织一部分迅速放入配置好的4%多聚甲醛固定液中固定24 h,后经梯度酒精脱水、二甲苯透明、透蜡、包埋。另一部分组织不经处理迅速放入-80 ℃冰箱保存。大鼠最后经过量的水合氯醛注射处死,实验过程符合实验动物伦理学标准。
1.2.4骨骼肌组织HE染色:经石蜡包埋后的骨骼肌,用石蜡切片机(Leica)切片,厚度为3 μm,切片放入60 ℃烤箱烤片1 h,再放入二甲苯中脱蜡15 min,重复2次,随后经梯度酒精复水,进行HE染色。染色的试剂盒购自碧云天(C0105)。苏木素染色7 min后,置入自来水中清洗6 min,随后用伊红染色37 s,梯度酒精脱水后置入二甲苯内透明15 min,最后用中性树胶封片,在显微镜下观察。随机在各组中选取5张切片,每张切片再随机选择5个视野,视野内各肌纤维横截面积大小通过Image J软件分析得出。
1.2.5免疫组织化学染色方法:选取4 μm的石蜡切片,放入60 ℃烤箱中烤片3 h,后放入二甲苯中脱蜡40 min,然后经梯度酒精复水,再将切片放入已经配置的枸橼酸钠(中杉金桥)缓冲液中,后将缓冲液放入已煮沸的高压锅内进行抗原修复,维持12 min。修复完成后,将缓冲液合并切片在常温下自然冷却,将切片放入PBS中洗3次,每次5 min,然后用10%山羊血清封闭2 h,甩净切片上的山羊血清,最后滴加MHC-I抗体,抗体浓度为1:4 000,4 ℃孵育过夜;第2天先用PBS清洗切片3次,每次5 min,然后根据免疫组织化学二抗试剂盒(中杉PV9002)操作步骤,先用10%过氧化氢封闭内源性过氧化氢酶10 min,经PBS清洗后用聚合物辅助剂孵育20 min,PBS清洗后孵育二抗30 min,用DAB显色液显色,最后用Olympus显微镜观察并摄取相应图片。
1.2.6Western Blot法:取70 mg的比目鱼肌充分剪碎,放入装有PMSF和RIPA混合液(PMSF:RIPA=1:99)的匀浆管内,将匀浆管置于匀浆器上匀浆,放置于冰上静置20 min,用离心机以3 000 r/min,4 ℃下离心5 min,经过超声裂解后,再经12 000 r/min离心20 min,吸取上清,放入-80 ℃冰箱保存。蛋白上样体系配置:先用BCA试剂盒(碧云天)测定蛋白浓度,加双蒸水及Loadingbuffer配置成上样体系。配置好后经100 ℃ 5 min将体系变性,放入-20 ℃冰箱保存。电泳时选用8%分离胶进行蛋白分离,后湿转至PVDF膜(Millipore)。转膜结束后,5%脱脂牛奶封闭2 h,TBST洗后,放入经稀释的MHC-I(Anti-Slow Skeletal Myosin Heavy chain,ab11083),雌激素相关受体γ(ERRγ,Abcam,ab128930),过氧化物酶体增殖物受体β(PPARβ,Santa Cruz,H-74)一抗中4 ℃孵育过夜,抗体稀释浓度分别为1:2 500,1:2 000,1:200;MHC-I蛋白二抗选用山羊抗小鼠(Biosharp),PPARβ、ERRγ二抗选用山羊抗兔,分别室温孵育2 h,二抗浓度1:2 000;内参Tubulin来自碧云天公司,稀释倍数为1:2 000,二抗选用山羊抗小鼠(Biosharp)室温孵育2 h,二抗浓度1:2 000。
1.2.7qRT-PCR RNA抽提方法:取150 mg组织研磨后加1 mL Trizol(Invitrogen)抽提试剂加入200 μL氯仿/异戊醇(24:1),振荡混匀30 s,放置5 min;用高速冷冻离心机,12 000 r/min,4 ℃离心10 min;将上清液小心转移到去酶的1.5 mL离心管里,加入与上清等体积的异丙醇和1.0 μL的糖原,室温下放置5 min;再经过高速冷冻离心机离心8 min (12 000 r/min,4 ℃);移去上清液,用70%酒精洗涤2次,每次700 μL,再分别离心2 min和5 min (12 000 r/min,4 ℃);待酒精挥发完全后,用15 μL DEPC-H2O溶解RNA。反应体系包括:0.1 mol/L DTT(Invitrogen)2 μL,40 U/μL Rnasin(Promega)0.5 μL,10 mmol/L dNTP(Takara)1.0 μL;Random Primer 1.0 μL,5×first Strand Buffer 4.0 μL,5 U/μL DnaseI(Takara)1.0 μL,RNA 10.5 μL。反转录:先放入冰中浴5 min,后加入40 U/μL RNasin 0.5 μL,200 U/μL SSII 0.5 μL。反应程序为:25 ℃(10 min),42 ℃(1 h),52 ℃(15 min),70 ℃(15 min)。Real-time PCR:反应体系25 μL,包括:5×R-PCR Buffer(5.0 μL),250 mmol/L Mg2+(0.3 μL),10 mmol/L dNTP(0.75 μL),10 μmol/L上游引物(0.5 μL),10 μmol/L下游引物(0.5 μL),25×SYBR GreenI(Bio-Rad,1.0 μL),10-3×Calibration(Bio-Rad,1.0 μL),5 U/μL HS-Ex-Taq(Takara,0.25 μL),ddH2O(14.7 μL),cDNA(1.0 μL)。反应程序:95 ℃(90 s),95 ℃(5 s),58 ℃(30 s)共40个循环,产生荧光信号监测点。然后5 ℃(1 min),58 ℃(1 min),58 ℃(10 s)。
1.3统计学处理方法 采用SPSS11.0统计学软件进行统计分析。计量资料用±s表示,用x2检验比较各组比目鱼肌肌纤维构成比,各组间肌纤维截面积及Western blot、PCR数值比较采用LSD-t检验。P<0.05为差异有统计学意义。
2 结果
2.14组比目鱼肌病理形态比较 正常组大鼠肌细胞形态饱满,排列有序,细胞核大小正常,见图1A;与正常组比,造模组及捆绑组,骨骼肌局部炎症反应明显,肌细胞核肥大,肌间质纤维化,大量形状异常的肌细胞;造模组、捆绑组和低频电刺激组肌纤维横截面积明显变小;与造模组和捆绑组比,低频电刺激组大鼠比目鱼肌周围血管数目增加,肌细胞形态正常,见图1和表1。
2.24组肌纤维类型构成比较 与正常组比,造模组和捆绑组大鼠比目鱼肌I型肌纤维数量明显下降,差异有统计学意义(P<0.05),见图2和表2;与造模组和捆绑组比,低频电刺激组大鼠比目鱼肌I型肌纤维数目明显增加,差异有统计学意义(P<0.05),见表2和图2。与正常组比,捆绑组和造模组大鼠比目鱼肌MHC-I mRNA明显下降,MHC-IIa、MHC-IIx mRNA明显升高,差异均有统计学意义(P<0.05);而低频电刺激组,大鼠比目鱼肌MHC-I mRNA含量相比捆绑组和造模组明显升高(P<0.05),而MHC-IIa、MHC-IIx mRNA含量明显下降(P<0.05)。

图1 比目鱼肌HE染色(×200)
表1 4组比目鱼肌平均截面积(n=5,±s,μm2)
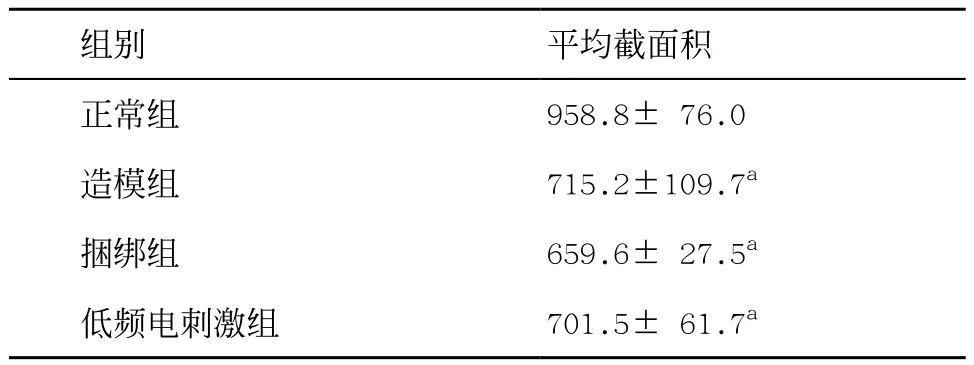
表1 4组比目鱼肌平均截面积(n=5,±s,μm2)
与正常组比:aP<0.05
组别 平均截面积正常组 958.8± 76.0造模组 715.2±109.7a捆绑组 659.6± 27.5a低频电刺激组 701.5± 61.7a
2.34组MHCI及其上游调节蛋白表达比较 造模组和捆绑组大鼠比目鱼肌MHC-I、PPARβ蛋白含量比正常组明显下降,差异有统计学意义(P<0.05);4组ERRγ表达水平差异没有统计学意义;与造模组和捆绑组比,低频电刺激组大鼠比目鱼肌MHC-I、PPARβ、ERRγ蛋白含量明显增加,差异有统计学意义(P<0.05)。见图4。
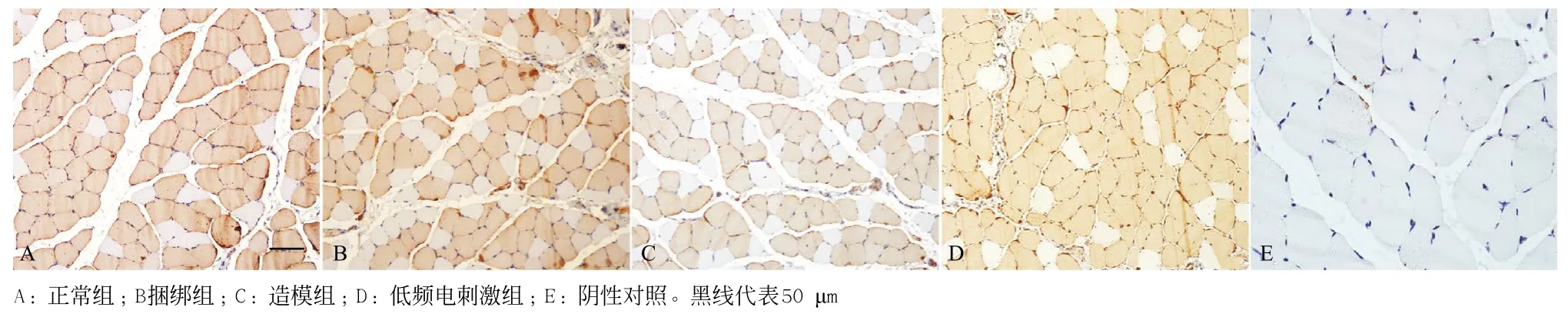
图2 4组免疫组织化学I型肌纤维染色(×200)
表2 4组比目鱼肌各型肌纤维相对mRNA含量(n=5,±s)
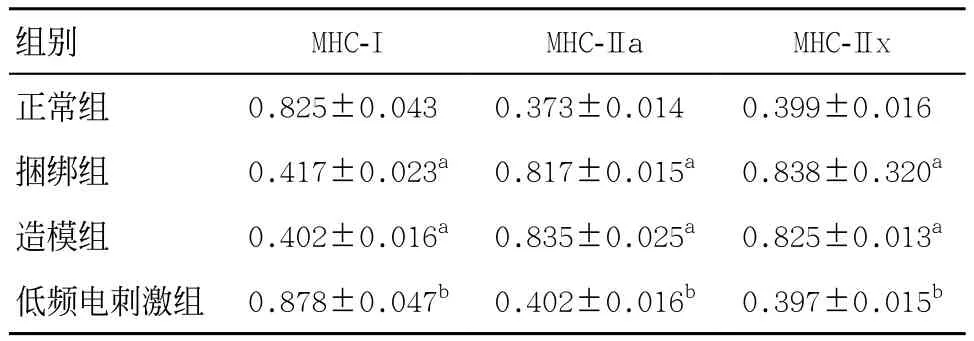
表2 4组比目鱼肌各型肌纤维相对mRNA含量(n=5,±s)
与正常组比:aP<0.05;与造模组和捆绑组比:bP<0.05
组别 MHC-I MHC-IIa MHC-IIx正常组 0.825±0.043a 0.373±0.014a 0.399±0.016a捆绑组 0.417±0.023a 0.817±0.015a 0.838±0.320a造模组 0.402±0.016a 0.835±0.025a 0.825±0.013a低频电刺激组 0.878±0.047b 0.402±0.016b 0.397±0.015b

图3 4组大鼠比目鱼肌各型肌纤维相对mRNA含量


图4 4组大鼠比目鱼肌中各蛋白Western Blot结果
3 讨论
COPD的肺外并发症在近年受到了越来越多的重视,诸多并发症中,因骨骼肌功能障碍与患者病死率密切相关,因而受到了关注。低频电刺激可有效促进骨骼肌卫星细胞增殖[13],增加COPD患者骨骼肌I型肌纤维含量[8],然而其对COPD患者骨骼肌功能的影响以及具体机制仍不十分明确。本研究发现:低氧高二氧化碳可引起大鼠比目鱼肌损伤及I型肌纤维向IIa/IIx转化,而低频电刺激可有效逆转该过程。低氧高二氧化碳及低频电刺激对比目鱼肌肌纤维形态及肌纤维类型的影响可能与ERRγ、PPARβ蛋白表达改变有关。
引起慢性低氧高二氧化碳模型大鼠比目鱼肌骨骼肌功能障碍的因素可有很多,通常认为骨骼肌炎症反应是重要诱因之一[14]。COPD患者骨骼肌炎症反应可能与其血液循环中白细胞含量增加有关[15],而肌细胞核肥大、肌间质纤维化均可因局部肌肉炎症引起[16],骨骼肌的炎症反应还与骨骼肌萎缩的发生关系密切[17]。骨骼肌横截面积与COPD患者运动能力呈现明显的正相关;同时轻症COPD患者,也可出现较明显的骨骼肌萎缩及运动能力下降[18]。本研究中模型大鼠肌纤维横截面积相比正常组明显下降,可能因大鼠长期暴露于低氧环境所致[19],但也可能由炎症反应等其他因素引起。不过,De Theije建立的低氧模型小鼠却并未发现比目鱼肌肌纤维横截面积有明显改变[20],产生不同结果的原因可能与建模方式有关。此外,我们观察到低频电刺激未明显增加模型大鼠肌纤维截面积,这表明低频电刺激尚不能逆转慢性低氧高二氧化碳所致模型大鼠比目鱼肌萎缩。
I型肌纤维线粒体含量十分丰富,且其能量代谢以有氧代谢为主,故其抗疲劳能力较强。II型纤维的供能方式为无氧酵解混合有氧代谢(IIa、IIx纤维)或者纯无氧酵解(IIb纤维),故其抗疲劳能力相对弱。骨骼肌各纤维的组成比并非一成不变,当受到低氧、废用或耐力运动等外界因素干预时,不同类型的肌纤维之间可互相转化,进而使肌肉功能能够适应外界环境的变化,保持相应的工作状态。本研究中,低氧高二氧化碳暴露后大鼠比目鱼肌I型肌纤维明显减少,而IIa/IIx型肌纤维含量明显升高,这表明低氧高二氧化碳可造成大鼠比目鱼肌由I型纤维向IIa/IIx这两类中间型肌纤维转化。而当采取低频电刺激大鼠比目鱼肌后,我们发现其I型肌纤维含量有明显升高,同时伴随IIa/IIx型肌纤维含量明显下降。这表明,低频电刺激可成功逆转由低氧高二氧化碳所致的大鼠比目鱼肌肌纤维类型转变。
为了进一步探讨造模及低频电刺激引起肌纤维类型转变的可能机制,我们还研究了PPARs。PPARs是核激素受体家族中的配体激活受体,其有三个亚型,分别是PPARα、PPARβ、PPARγ。研究发现,低氧处理的C2C12细胞,其PPARβ mRNA水平明显下降[21]。Gan等[22]通过建立运动小鼠模型发现PPARβ与小鼠骨骼肌I型肌纤维的表达密切相关,并且PPARβ是通过直接激活ERRγ实现的。ERRγ属于核受体(nuclear receptor,NR)超家族的一员,是一种在慢肌纤维内丰富表达的蛋白。研究表明,ERRγ在调控骨骼肌纤维类型、线粒体功能、血管生成方面起重要作用[23]。本研究中模型组动物比目鱼肌PPARβ水平相比正常组明显下降,然而比目鱼肌ERRγ水平却没有明显改变,这与其I型肌纤维含量的改变并不平行,为此我们通过相关文献查阅发现:研究ERRγ蛋白过表达处理后的比目鱼肌发现比目鱼肌I型肌纤维含量并没有明显改变[24],同样ERRγ敲除后小鼠比目鱼肌I型肌纤维同样没有明显改变[22],这表明ERRγ可能并未参与比目鱼肌I型肌纤维的调节,PPARβ则可能是直接或间接地通过其他通路来调节比目鱼肌肌纤维类型。前期鲜有研究关注低频电刺激对骨骼肌ERRγ表达的影响,本研究显示,低频电刺激可显著提高骨骼肌ERRγ的表达,同时,其上游蛋白PPARβ的水平亦有明显的升高。然而考虑到ERRγ可能并未参与到比目鱼肌I型肌纤维的调控,为此我们猜测低频电刺激引起的I型肌纤维含量上升可能与PPARβ的水平升高有关。尽管ERRγ未参与调节比目鱼肌I型肌纤维,但低频电刺激后其含量亦明显增加,结合观察到该组大鼠比目鱼肌内血管数目有所增加,我们猜测低频电刺激很有可能是增加了ERRγ表达进而促进了比目鱼肌的血管生成,但确切机制仍有待进一步研究。
本研究发现,低频电刺激可逆转因低氧高二氧化碳所致的I型纤维向II型纤维转化,其机制可能与PPARβ蛋白含量升高有关,而ERRγ则可能与低频电刺激诱导的血管增生有关。
参考文献:
[1]DECRAMER M,JANSSENS W,MIRAVITLLES M.Chronic obstructive pulmonary disease and comorbidities[J].Lancet Respir Med,2013,1(1):73-83.
[2]SWALLOW E B,REYES D,HOPKINSON N S,et al.Quadriceps strength predicts mortality in patients with moderate to severe chronic obstructive pulmonary disease[J].Thorax,2007,62(2):115-120.
[3]BARREIRO E,DE LA PUENTE B,MINGUELLA J,et al.Oxidative stress and respiratory muscle dysfunction in severe chronic obstructive pulmonary disease[J].Am J Respir Crit Care Med,2005,171(10):1116-1124.
[4]KOECHLIN C,MALTAIS F,SAEY D,et al.Hypoxaemia enhances peripheral muscle oxidative stress in chronic obstructive pulmonary disease[J].Thorax,2005,60(10):834-841.
[5]PUENTE-MAESTU L,LAZARO A,HUMANES B.Metabolic derangements in COPD muscle dysfunction[J].J Appl Physiol (1985),2013,114(9):1282-1290.
[6]ELIASON G,ABDEL-HALIM S M,PIEHL-AULIN K,et al.Alterations in the muscle-to-capillary interface in patients with different degrees of chronic obstructive pulmonary disease[J].Respir Res,2010,11(1):97-101.
[7]BOURJEILY-HABR G,ROCHESTER C L,PALERMO F,et al.Randomised controlled trial of transcutaneous electrical muscle stimulation of the lower extremities in patients with chronic obstructive pulmonary disease[J].Thorax,2002,57(12):1045-1049.
[8]ABDELLAOUI A,PREFAUT C,GOUZI F,et al.Skeletal muscle effects of electrostimulation after COPD exacerbation:a pilot study[J].Eur Respir J,2011,38(4):781-788.
[9]SILLEN M J,FRANSSEN F M,DELBRESSINE J M,et al.Efficacy of lower-limb muscle training modalities in severely dyspnoeic individuals with COPD and quadriceps muscle weakness:results from the DICES trial[J].Thorax,2014,69 (6):525-531.
[10]EL-KHOURY R,BRADFORD A,O’HALLORAN K D.Chronic hypobaric hypoxia increases isolated rat fast-twitch and slow-twitch limb muscle force and fatigue[J].Physiol Res,2012,61(2):195-201.
[11]徐漫欢,范小芳,王小同,等.吸入低氧高二氧化碳对小鼠右心室收缩压及肺小动脉重建的影响[J].浙江医学,2008,30(1):41-43.
[12]ARMSTRONG R B,PHELPS R O.Muscle fiber type composition of the rat hindlimb[J].Am J Anat,1984,171(3):259-272.
[13]WAN Q,YEUNG S S,CHEUNG K K,et al.Optimizing electrical stimulation for promoting satellite cell proliferation in muscle disuse atrophy[J].Am J Phys Med Rehabil,2016,95(1):28-38.
[14]LANGEN R C,GOSKER H R,REMELS A H,et al.Triggers and mechanisms of skeletal muscle wasting in chronic obstructive pulmonary disease[J].Int J Biochem Cell Biol,2013,45(10):2245-2256.
[15]OUDIJK E J,NIJHUIS E H,ZWANK M D,et al.Systemic inflammation in COPD visualised by gene profiling in peripheral blood neutrophils[J].Thorax,2005,60(7):538-544.
[16]MANN C J,PERDIGUERO E,KHARRAZ Y,et al.Aberrant repair and fibrosis development in skeletal muscle[J].Skelet Muscle,2011,1(1):21.
[17]MUSCARITOLI M,ANKER S D,ARGILES J,et al.Consensus definition of sarcopenia,cachexia and pre-cachexia:Joint document elaborated by special Interest Groups (SIG) “cachexia-anorexia in chronic wasting diseases” and “nutrition in geriatrics”[J].Clin Nutr,2010,29(2):154-159.
[18]SHRIKRISHNA D,PATEL M,TANNER R J,et al.Quadriceps wasting and physical inactivity in patients with COPD [J].Eur Respir J,2012,40(5):1115-1122.
[19]包绍智,房春燕,王小同,等.慢性低氧高二氧化碳对小鼠骨骼肌形态的影响[J].温州医学院院报,2009,39(1):8 -10.
[20]DE THEIJE C C,LANGEN R C,LAMERS W H,et al.Differential sensitivity of oxidative and glycolytic muscles to hypoxia-induced muscle atrophy[J].J Appl Physiol,2014,118(2):200-211.
[21]SLOT I G,SCHOLS A M,VOSSE B A,et al.Hypoxia differentially regulates muscle oxidative fiber type and metabolism in a HIF-1alpha-dependent manner[J].Cell Signal,2014,26(9):1837-1845.
[22]GAN Z,RUMSEY J,HAZEN B C,et al.Nuclear receptor/ microRNA circuitry links muscle fiber type to energy metabolism[J].J Clin Invest,2013,123(6):2564-2575.
[23]NARKAR V A,FAN W,DOWNES M,et al.Exercise and PGC-1alpha-independent synchronization of type I muscle metabolism and vasculature by ERRgamma[J].Cell Metab,2011,13(3):283-293.
[24]MATSAKS A,YADAV V,LORCA S,et al.Revascularization of ischemic skeletal muscle by estrogen-related receptor-γ[J].Circ Res,2012,110(8):1087-1096.
(本文编辑:吴彬)
·论著·
Effect of low frequency electrical stimulation on soleus muscle fiber type of hypoxia-hypercapnia rats
HUANG Shiyuan,JIANG Xianxun,SHEN Jie,WANG Xiaotong.Center of Neurology and Rehabilitation,the Second Affiliated Hospital of Wenzhou Medical University,Wenzhou,325027
Abstract:Objective:To explore the effect of low frequency electrical stimulation on transition of rats’ soleus muscle fibre type after the long term chronic hypoxia-hypercapnia exposure.Methods:Forty male Sprague Dawley rats were randomly divided into four groups:the Normal Control (NC),Hypoxia-hypercapnia (HH),Hypoxia-hypercapnia+Tie-up (HT) and Low-frequency electrical stimulation (LFES).The HH,HT and LFES were placed in a chamber with hypoxia-hypercapnia for 8 h/d,4 weeks in all.After 2 weeks,30 min,30 Hz,1:1 cycle electrical stimulation was applied in the LEFS,this procedure lasted 2 weeks.The HT were only tied up and placed with electrode plates.Hematoxylin and Eeosin staining was used to observe pathological change of soleus muscle.Soleus fiber composition was visualized with immunochemistry,qRT-PCR was used to detect mRNA levels of MHC-I,MHC-IIa and IIx.Proteins expression of MHC-I,ERRγ,PPARβ was tested by Western Blot.Results:Increased inflammation cells and MHC-IIa/IIx mRNA,decreased cross section area,MHC-I,PPARβ protein and MHC-I mRNA were found in the HH and HT group compared with the NC.Compared with the HH and HT respectively,inflammation cells and MHC-IIa/IIx mRNA level were decreased,while mRNA level of MHC-I and protein expression of MHC-I,ERRγ and PPARβ were significantly increased after LFES.Conclusion:Hypoxia-hypercapnia leads to soleus muscle transition of MHC-I into MHC-IIa/IIx.However,LFES may reverse the muscle fiber transition induced by HH.PPARβ and ERRγ may have participates in this process.
Key words:hypoxia-hypercapnia; electrical stimulation; rats; soleus; myosin heavy chain
通信作者:王小同,教授,硕士生导师,Email:wangxt22@163.com。
作者简介:黄世园(1991-),男,浙江台州人,硕士生。
基金项目:浙江省自然科学基金资助项目(Y2080503)。
收稿日期:2015-07-26
[中图分类号]R337.5
[文献标志码]A
DOI:10.3969/j.issn.2095-9400.2016.03.001
猜你喜欢
中国当代医药(2017年2期)2017-03-18
中国当代医药(2016年32期)2017-02-23
中国当代医药(2016年31期)2017-02-23
科技创新与应用(2016年36期)2017-02-21
中国医药科学(2016年14期)2017-02-06
中国实用医药(2016年27期)2016-11-30
上海医药(2016年16期)2016-09-21
现代养生·下半月(2015年6期)2015-09-07
中国当代医药(2014年31期)2014-12-09
右江医学(2014年3期)2014-08-08
