
辽东山区紫椴、花曲柳碳素积累及主要营养元素分布特征1)
2016-05-06 07:43:51毛沂新尤文忠张慧东王睿照魏文俊颜廷武
东北林业大学学报 2016年3期
关键词:紫椴
毛沂新 尤文忠 张慧东 王睿照 魏文俊 颜廷武
(辽宁省林业科学研究院,沈阳,110032)
辽东山区紫椴、花曲柳碳素积累及主要营养元素分布特征1)
毛沂新尤文忠张慧东王睿照魏文俊颜廷武
(辽宁省林业科学研究院,沈阳,110032)
摘要以辽东山区天然阔叶混交林重要组成树种紫椴和花曲柳为研究对象,通过生物量实测法,探究紫椴和花曲柳的碳素积累的变化特征以及不同器官氮(N)、磷(P)、钾(K)元素的分配格局。结果表明:花曲柳树干的平均固碳量和连年固碳量整体上高于紫椴,随林龄的增长差距有逐渐增大的趋势;40年生花曲柳树干平均碳素密度为322.18 kg·m(-3),36年生紫椴为272.63 kg·m(-3)。在养分分配方面,紫椴和花曲柳单株N、P、K元素的积累量顺序一致(N>K>P),但在不同器官中各元素积累顺序有些差异。紫椴各器官养分积累总量顺序为:树干>树根>树枝>树叶;花曲柳各器官养分积累总量顺序为:树根>树干>树枝>树叶。紫椴与花曲柳树叶的养分归还量占各自总体的比例分别为4.11%和17.32%,而树叶、树枝与根的养分积累比重合计分别为57.07%和75.31%,三者在生态系统养分循环中占有重要作用。
关键词紫椴;花曲柳;固碳量;碳密度
分类号S718.55+4.2
Carbon Accumulation and Distribution of Main Nutrient Elements ofFraxinusrhynchophyllaandTiliaamurensisin the Mountainous Region of Eastern Liaoning Province, China
Mao Yixin, You Wenzhong, Zhang Huidong, Wang Ruizhao, Wei Wenjun, Yan Tingwu
(Liaoning Academy of Forestry, Shenyang 110032, P. R. China)//Journal of Northeast Forestry University,2016,44(3):21-25.
Based on the biomass data ofFraxinusrhynchophyllaandTiliaamurensisin broad-leaf mixed forest in the mountainous region of eastern Liaoning Province, we studied the features of carbon accumulation and nutrient distribution (N, P, K) in organs of these two species. The average stem carbon sequestration and the annual stem carbon sequestration ofF.rhynchophyllawere both higher than those ofT.amurensis, and there were a gradually increasing gap of the average stem carbon sequestration and the annual stem carbon sequestration between two species during the growing age. Average stem carbon density of 40-yearF.rhynchophyllawas 322.18 kg·m-3, and that of 36-year-oldT.amurensiswas272.63 kg·m-3. Nutrients accumulation descending order of two species was N, K, and P, but there were some variations of nutrients accumulation order in different organs of two species. The total accumulation of N, P and K between organs ofT.amurensiswas decreased as follows: stem, root, branch, and leaf, and those ofF.rhynchophyllawere listed in descending order of root, stem, branch, and leaf. The proportions of nutrients accumulation in leaves annually released to soil byF.rhynchophyllaandT.amurensisaccounted for 4.11% and 17.32%, respectively. Total proportions of nutrients accumulation in leaves, branches and roots of two species were 57.07% and 75.31%, respectively.
KeywordsTilia amurensis; Fraxinus rhynchophylla; Carbon sequestration; Carbon density
林木的固碳释氧与营养物质积累是森林生态系统服务功能的重要组成部分[1-3]。通过对森林生态系统服务功能进行准确评估,以应对全球气候变化、土壤养分流失等诸多生态环境问题[4-6]。同时,为我国制定林业政策及森林经营管理等方面提供科学依据,为国家生态文明建设发挥重要的支撑作用[7-9]。
天然阔叶混交林是辽东山区主要森林类型之一,天然阔叶林具有生长速度快、林分蓄积量大等特点[10-11]。花曲柳(Fraxinusrhynchophylla)和紫椴(Tiliaamurensis)作为该地区天然阔叶混交林重要的组成树种,也是我国东北地区重要的、具有较高经济价值的硬阔树种[12-14]。由于对木材长期依赖和不合理采伐等历史原因,致使这些珍贵阔叶树种资源急剧下降[11]。国内外多在大尺度上对森林碳储量研究较多,而对于林分以下等小尺度以及某一特定树种的固碳能力、养分积累与分配的研究相对较少。本文以辽东山区天然阔叶混交林中紫椴和花曲柳为研究对象,对二者树干碳素积累过程及不同器官的营养元素积累与分配格局进行对比分析。旨在探索紫椴和花曲柳固碳能力与养分积累的规律,为制定科学的林业政策和天然林保护工程的实施、实现森林资源的可持续发展提供理论依据。
1研究地概况
试验地设置于辽宁白石砬子国家级自然保护区内,该区域在辽东山区宽甸县北部,地理坐标为东经124°44′~124°57′,北纬40°50′~40°57′。该研究区属长白山余脉,平均海拔866.5 m,最高海拔1 270.5 m。该地区属温带大陆性季风气候,由于受地形条件的影响,具有典型的山地气候。年平均气温5.3 ℃,最高温度33.7 ℃,最低温度-32.5 ℃;年均日照时间1 841 h,无霜期132 d,年均降水量1 349 mm,年均蒸发量885 mm;年均相对湿度73%,雨热同季,集中于7—9月份。植物资源丰富,组成树种有花曲柳、蒙古栎(Quercusmongolica)、紫椴、胡桃楸(Juglansmandshurica)、色木槭(Acermono)、水曲柳(Fraxinusmandschurica)、黄檗(Phellodendronamurense)等。林分郁闭度为0.7~0.9,灌、草层盖度分别为0.32~0.67和0.50~0.81。土壤主要为暗棕色森林土和棕色森林土。
2研究方法
2.1样地设置与野外调查
在研究区内选择有代表性的、扰动少的天然阔叶混交林。设置20 m×30 m样地3块,对样地内树木进行每木检尺,同时调查样地立地因子和各项生长因子(见表1)。在3块样地中分别选取紫椴和花曲柳标准木各3株,总计18株。前期已通过方差分析结合差异显著性多重比较分别对紫椴与花曲柳同一树种的不同地区的样品进行分析,结果并没有出现显著性差异,说明样地的选择和取样对同一树种在不同地区之间并没有造成实质性的影响,以避免由样地而引起的差异。紫椴和花曲柳生长情况见表2。

表1 研究样地基本情况

表2 紫椴、花曲柳的生长概况
2.2样品采集与制备
将标准木从根颈处伐倒,分别在树干的底部、1.3、3.6 m处分别截取5 cm厚的圆盘,剩余梢头每2 m处分段,截取5 cm厚的圆盘,带回实验室对其进行树干解析。同时,对树干、树枝、树叶采用全收获法在现场称取各部分鲜质量。在树冠上、中、下3层分别对枝、叶取样。对标准木树干在各分段底部进行取样。采用全挖法收获全部树根,称取鲜质量,随机截取根鲜样,密封保存带回实验室,用流动水浸泡、漂洗。将以上各植物器官样品分别进行等比例混合,65 ℃恒温烘干至恒质量,称取各组分生物量。对紫椴和花曲柳树叶、树枝、树干和树根样品,分别烘干、粉碎、过筛后,进行有机碳和营养元素测定。碳(C)采用浓H2SO4-K2CrO4水合热法测定,氮(N)采用凯氏法测定,磷(P)采用钼锑抗比色法测定,钾(K)采用火焰光度计法测定。
2.3数据处理
根据树干碳质量分数和木材密度将树木解析得出的材积连年生长量、平均生长量和总生长量转换为树干平均固碳量(Cθ)、连年固碳量(CZ)和碳素总储量(C)。连年固碳量为树木在龄阶内(2 a)的平均碳储量;平均固碳量为树木生长至某一年龄时的平均碳储量,总碳素储量为林木生长至某一年龄时的实际碳储量。平均碳素密度是用来表征单位材积树干碳素储量的高低。其中:ρ=B/V;C=V×ρ×c;Cθ=θ×ρ×c;CZ=Z×ρ×c;D=C/V。式中:ρ为木材密度;B为树干生物量;V为树干材积;C为树干总固碳量;Cθ为树干平均固碳量;CZ为树干连年固碳量;θ为材积平均生长量;Z为材积的连年生长量;c为树干碳质量分数;D为树干碳密度。
养分积累量为各器官养分元素质量分数与其生物量的乘积[7]。各器官养分积累量为不同高度各段养分积累量之和。采用SPSS16.0软件对各树种养分分布的差异性进行方差分析(ANOVA),同时进行差异显著性多重比较(LSD)。
3结果与分析
3.1紫椴和花曲柳空间生物量与碳储量分配状况
由表3可知,紫椴和花曲柳生物量与固碳量在空间上的分布规律较为一致。两树种单株各器官生物量与固碳量由大到小的顺序均为:干、根、枝、叶。其中,二者的干材生物量与碳储量所占比重最大,达50%以上;枝、叶所占比重较小,合计在20%以下。

表3 紫椴和花曲柳空间生物量与树干碳储量分配状况
3.2紫椴和花曲柳树干碳素积累过程
由图1可知,紫椴在前10 a单株碳储量增长趋势比较平缓(固碳量为0.21 kg,贡献率为0.01%);>10~20 a间,单株固碳增速逐步加快(固碳量为3.02 kg,贡献率为12.62%);>20~36 a间,单株固碳增速进一步提升(固碳量为20.67 kg,贡献率为86.48%)。因此,36年生的紫椴90.79%的固碳储量主要集中在生长期第18 a以后。
花曲柳在前12 a单株碳储量变化不大(固碳量为0.55 kg,贡献率为0.01%);>12~22 a期间单株碳储量增速逐步加快(固碳量为5.40 kg,贡献率为8.62%);>22~40 a期间单株碳储量增速明显提升(固碳量为56.68 kg,贡献率为90.50%)。因此,40年生的花曲柳90.50%的固碳储量主要集中在生长期第22 a以后。

紫椴和花曲柳单株树干固碳能力与碳储量为各树种标准木的平均结果,由于各标准木的林木年龄略微有所差异,故对于各树种固碳能力的计算均取最短林龄的年限,即紫椴为36 a内的平均值,花曲柳为40 a内的平均值。
图1紫椴和花曲柳树干碳储量
由图2可知,在0~36 a间,紫椴单株树干连年固碳量为0.002~2.085 kg,固碳速度随林龄增长逐渐增大。在10 a以前,碳积累速度变化不大,使得总固碳量增长缓慢;>10~22 a间,固碳速率逐步提高,连年固碳量达到0.858 kg;>22~36 a间,前期固碳量增长幅度有所下降,之后随林龄的增长再次明显增大,连年固碳量达到2.085 kg。紫椴单株树干平均固碳量在前10 a中比较稳定,>10~36 a间,平均固碳量逐步稳定增加,其增长幅度总体低于连年固碳量。
0~40 a间,花曲柳单株树干连年固碳量为0.001~5.578 kg。在前12 a花曲柳连年固碳量积累缓慢,使得总固碳量增长缓慢;在>12~32 a中,连年固碳量迅速增高,由0.123 kg达到3.264 kg;在>32~40 a间,连年固碳量开始变缓,达到5.058 kg。花曲柳单株树干平均固碳量前10 a期间缓慢上升,在>10~40 a期间,平均固碳量逐步平稳上升达到1.566 kg。
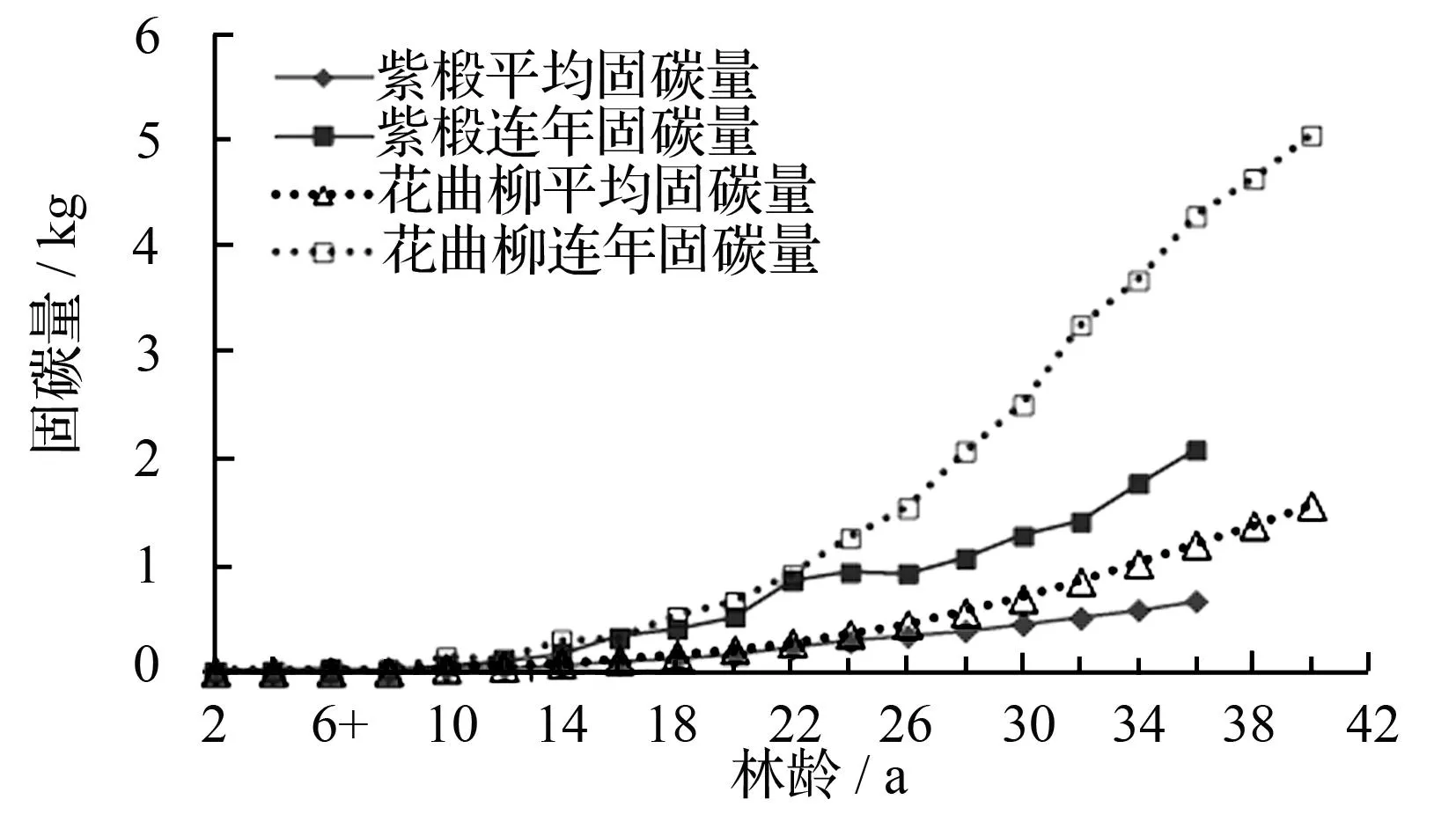
图2 紫椴和花曲柳树干碳素积累过程
3.3各器官主要营养元素分布与积累
由表4可知,紫椴不同器官中(N、P、K)各营养元素的平均质量分数由大到小的顺序均为:树叶、树枝、树根、树干;花曲柳不同器官中(N、P、K)各营养元素的平均质量分数由大到小的顺序均为:树叶、树根、树枝、树干。除花曲柳树干中的K质量分数最高外,紫椴和花曲柳各养分元素平均质量分数在各器官中分布总体一致(N最高、K次之、P最低)。
表4紫椴和花曲柳各器官营养元素平均质量分数

g·kg-1
注:表中数值为“平均值±标准误”;同列不同小写字母表示差异显著(p<0.05)。
由表5可知,3种主要养分元素(N、P、K)在紫椴和花曲柳单株中总积累量分别为(555.01±128.00)g和(1 724.37±252.06)g,地上各器官养分积累量分别为(350.55±85.37)g和(1 061.84±106.75)g,占各自总积累量的63.16%和61.58%;紫椴叶、枝、干、根养分积累量所占比重分别为4.11%、16.12%、42.93%、36.84%;花曲柳叶、枝、干、根养分积累量所占比重分别为17.32%、19.57%、24.69%、38.42%。
紫椴和花曲柳单株各养分元素积累量由大到小顺序均为:N、K、P。两树种地上部分、地下部分与单株中各元素积累量大小顺序一致。养分元素的积累顺序在两树种各器官中,除花曲柳树干中N元素积累量低于K外,总体也呈N积累量始终最高,K次之,P最低。
同种养分元素的积累量在不同器官中的分配情况:P和K元素的积累量在紫椴和花曲柳中均为树干>树根>树枝>树叶;N元素的积累量在紫椴中为树根>树干>树枝>树叶;在花曲柳中,N元素在树干中积累量显著低于根部。紫椴中同种养分元素积累量与积累总量在不同器官中的分配特征为树干与树根之间,树枝与树叶之间均无显著性差异(p<0.05);在花曲柳中,树枝与树叶之间无显著性差异(p<0.05)。
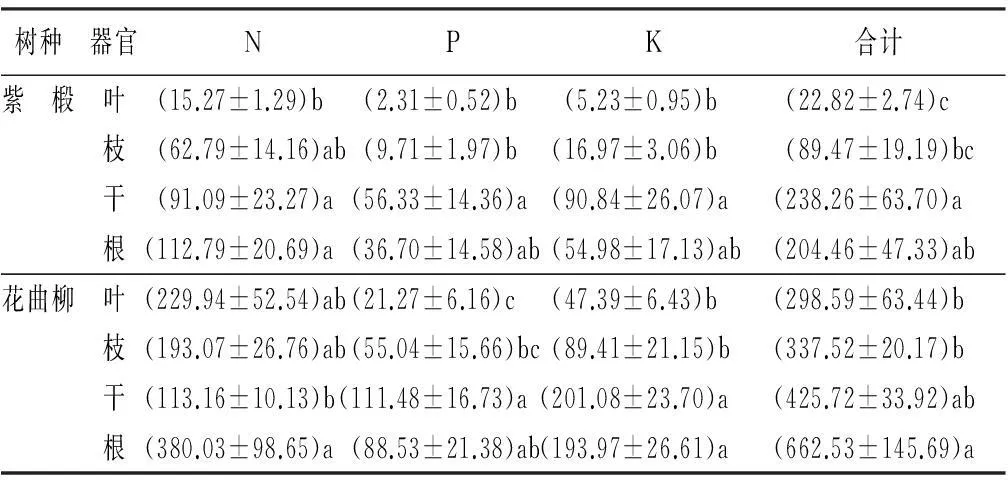
表5 紫椴和花曲柳各器官营养元素积累量 g
注:表中数值为“平均值±标准误”;同列不同小写字母表示差异显著(p<0.05)。
4讨论
林木生物量是林地生产力与养分积累的基础,是反映林分结构与功能的主要标志之一,也是估算森林固碳能力的重要参数[15-19]。本研究中紫椴与花曲柳地下生物量占全株生物量的比重分别为21%和24%,与前人研究成果较为一致[20]。花曲柳树干的平均固碳量和连年固碳量整体上高于紫椴,且随林龄的增长这种差距有逐渐增大的趋势,这是由于花曲柳的单株木材密度、树干碳质量分数和树干材积均高于紫椴。在第36 a时,花曲柳树干总碳储量为43.22 kg,紫椴树干碳储量为23.90 kg,花曲柳树干总碳储量是紫椴的1.81倍。干材作为乔木最主要的固碳组成部分,紫椴与花曲柳的树干碳储量分别占其单木碳储量的75%和58%。花曲柳树干平均碳素密度(322.18 kg·m-3)是紫椴(272.63 kg·m-3)的1.18倍,表明单位材积花曲柳的固碳量比紫椴大。两树种的树干碳素积累的快速增长期大致在10~12 a后,而90%的碳储量主要集中在18~20 a以后,说明在幼龄的中后期与中龄前期是两树种生长发育与碳素积累的关键阶段。所以,在森林资源可持续经营与管理的过程中,要格外重视两树种生长的关键阶段,通过适时的林分抚育(如:适度定株修枝、抚育间伐、科学合理地改善林地水肥条件),促进目标树种生长,有效提高林木品质与固碳能力[21-26]。由于历史原因,选择该地区混交林的林龄结构多为近熟林,不能完整的反映出两树种在整个生命周期中碳素积累的整体趋势,所以不能够对两树种的固碳成熟年龄准确预测。
森林生态系统在特定时空范围内,光照、水、热等外界影响因子通常变化不大或者呈一定的规律性,系统的养分循环对于森林生态系统的影响更为突出[27-30]。不同树种对养分元素的吸收与利用是森林生态系统养分循环研究的重点。由于受植物本身的生理特性和不同养分元素对植物生理作用的影响,使得不同树种间及其各营养器官对所需营养元素种类和数量均存在差异[31]。紫椴与花曲柳单株N、P、K各元素的积累量顺序一致,但在不同器官中各元素积累有一定差异。本研究对养分积累量的计算是根据各器官中养分含量与生物量的乘积得出的,所以养分积累量受不同器官的生物量及其养分含量的共同作用[32-33]。尽管养分含量在树干、树根中偏低,但树干、树根生物量却高于其他各器官,使得树干与树根养分积累总量高于树枝、树叶。作为落叶阔叶树种,紫椴和花曲柳树叶中的养分积累量为当年积累形成,在生长季后期以落叶分解形式将养分回归到土壤层[34-36]。其中,紫椴和花曲柳树叶的主要养分归还量占各自总体的比例为4.11%和17.32%,而树叶、树枝与根的养分积累比重合计分别为57.07%和75.31%,三者在森林生态系统养分循环中的作用不可低估[37]。因此,不合理的森林采伐与利用,将从森林生态系统中的带走大量养分,造成植物对土壤养分归还量的损失,降低土壤养分的有效供给与林地肥力。在采伐过程中,将树根、枝、叶保留在林地内,最大限度的减小因采伐造成的养分输出,这对保持地力、维持林地生产力稳定是十分必要的[38]。在森林资源有限的前提下,从而实现森林的社会、经济与生态综合效益的最大化。
参考文献
[1]NELSON E, SANDER H, HAWTHORNE P, et al. Projecting global land-use change and its effect on ecosystem service provision and biodiversity with simple models[J]. Plos One,2010,5(12):1-22.doi:10.1371/journal.pone.0014327.
[2]POLASKY S, NELSON E, PENNINGTON D, et al. The impact of land-use change on ecosystem services, biodiversity and returns to landowners: A case study in the state of Minnesota[J]. Environmental & Resource Economics,2011,48(2):219-242.
[3]于贵瑞,李海涛,王绍强,等.全球变化与陆地生态系统碳循环和碳蓄积[M].北京:气象出版社,2003.
[4]WATSON R T, NOBLE IR, BOLIN B. Land use, land-use change and forestry[M]. Cambridge: Cambridge University Press,2000.
[5]中国森林资源核算及纳入绿色GDP研究项目组.绿色国民经济框架下的中国森林核算研究[M].北京:中国林业出版社,2010.
[6]中国森林生态服务功能评估项目组.中国森林生态服务功能评估[M].北京:中国林业出版社,2010.
[7]黄建辉,陈灵芝,韩兴国.辽东栎枝条分解过程中几种主要营养元素的变化[J].植物生态学报,1998,22(5):398-402.
[8]蒋俊明,费世民,李吉跃,等.苦竹各器官主要营养元素分布及采伐的养分输出[J].四川林业科技,2007,28(2):15-19.
[9]刘广全,赵士洞,土小宁,等.秦岭松栎林带生物量及其营养元素分布特征[J].林业科学,2001,37(1):28-36.
[10]刘足根,朱教君,袁小兰,等.辽东山区次生林主要树种种群结构和格局[J].北京林业大学学报,2007,29(1):12-18.
[11]吴可,殷鸣放,周永斌,等.白石砬子国家级自然保护区森林蓄积连年生长率的研究[J].西北林学院学报,2010,25(6):203-206.
[12]项凤武,刘艳华.花曲柳的分布、生长及天然更新的研究[J].吉林林学院学报,1997,13(4):212-215.
[13]马微微,许颖,穆立蔷.紫椴的研究进展及其资源的开发利用[J].中国林副特产,2009(2):88-90.
[14]聂绍荃,关文彬,杨国亭,等.紫椴种群生态学研究[M].哈尔滨:东北林业大学出版社,1992.
[15]MERINO A, BALBOA M A, SOALLEIRO R R, et al. Nutrient exports under different harvesting regimes in fast-growing forest plantations in southern Europe[J]. Forest Ecology and Management,2005,207(3):325-339.
[16]THIFFAULT E, HANNAM K D, PARÉ D, et al. Effects of forest biomass harvesting on soil productivity in boreal and temperate forests-A review[J]. Environmental Reviews,2011,19(1):278-309.doi:10.1139/a11-009.
[17]罗云建,张小全,王效科,等.森林生物量的估算方法及其研究进展[J].林业科学,2009,45(8):129-134.
[18]余新晓,鲁绍伟,靳芳,等.中国森林生态系统服务功能价值评估[J].生态学报,2005,25(8):2096-2102.
[19]杨明,汪思龙,张伟东,等.杉木人工林生物量与养分积累动态[J].应用生态学报,2010,21(7):1674-1680.
[20]HOUGHTON R A, LAWRENCE K T, HACKLER J L, et al. The spatial distribution of forest biomass in the Brazilian Amazon: a comparison of estimates[J]. Global Change Biology,2001,7(7):731-746.
[21]尤文忠,魏文俊,邢兆凯,等.辽东山区落叶松人工林和蒙古栎天然次生林的固碳功能[J].东北林业大学学报,2011,39(10):21-24.
[22]FOOTEN P W, HARRISON R B, STRAHM B D. Long-term effects of nitrogen fertilization on the productivity of subsequent stands of Douglas-fir in the Pacific Northwest[J]. Forest Ecology and Management,2009,258(10):2194-2198.
[23]HOOVER C, STOUT S. The carbon consequences of thinning techniques: stand structure makes a difference[J]. Journal of Forestry,2007,105(5):266-270.
[24]王群超.基于固碳效益的森林最优轮伐期的确定[J].东北林业大学学报,2011,39(6):98-100.
[25]王祖华,刘红梅,王晓杰,等.经营措施对森林生态系统碳储量影响的研究进展[J].西北农林科技大学学报(自然科学版),2011,39(1):83-88.
[26]张鼎华,叶章发.抚育间伐对人工林土壤肥力的影响[J].应用生态学报,2001,12(5):672-676.
[27]聂道平.森林生态系统营养元素的生物循环[J].林业科学研究,1991,4(4):435-440.
[28]俞白楠.人工林经营与森林可持续发展[J].福建林业科技,2000,27(4):24-27.
[29]张晓娟,魏天兴,荆丽波,等.晋西黄土区天然次生林营养元素分配与积累研究[J].北京林业大学学报,2008,30(3):57-62.
[30]丁宝永,孙继华.水曲柳天然林生物生产力及营养元素的积累与分布的研究[J].东北林业大学学报,1989,(4):1-9.
[31]蒋高明.植物生理生态学[M].北京:高等教育出版社,2004.
[32]PERI P L, GARGAGLIONE V, PASTUR G M. Above-and belowground nutrients storage and biomass accumulation in marginal Nothofagus antarctica forests in Southern Patagonia[J]. Forest Ecology and Management,2008,255(7):2502-2511.
[33]尹娜.黄土区人工林生物量及养分积累分布研究[D].北京:北京林业大学,2008.
[34]肖洋,陈丽华,余新晓.北京密云麻栎人工混交林凋落物养分归还特征[J].东北林业大学学报,2010,38(7):13-15.
[35]姚迎九.樟树人工林生物量和养分积累与分布[D].长沙:中南林学院,2003.
[36]张晓娟.山西吉县天然次生林生物量和营养元素积累与分布研究[D].北京:北京林业大学,2008.
[37]曹建华,李小波,赵春梅,等.森林生态系统养分循环研究进展[J].热带农业科学,2007,27(6):68-79.
[38]项文化,田大伦.不同年龄阶段马尾松人工林养分循环的研究[J].植物生态学报,2002,26(1):89-95.
收稿日期:2015年9月16日。
第一作者简介:毛沂新,男,1985年11月生,辽宁省林业科学研究院,工程师。E-mail:dmoomoo@163.com。通信作者:尤文忠,辽宁省林业科学研究院,教授级高级工程师。E-mail:wzhyou2002@163.com。
1)林业公益性行业科研专项(201204101、201404303);国家科技支撑计划项目(2012BAD22B04);辽宁省农业领域青年科技创新人才培养计划项目(2014015)。
责任编辑:王广建。
猜你喜欢
辽宁林业科技(2023年5期)2023-11-08 07:26:20
北华大学学报(自然科学版)(2022年6期)2022-11-22 02:28:46
北华大学学报(自然科学版)(2022年6期)2022-11-22 02:28:44
防护林科技(2022年6期)2022-10-24 08:08:16
林业科技(2021年5期)2021-12-24 17:05:13
中国林副特产(2020年6期)2020-12-16 00:24:28
农家科技下旬刊(2019年11期)2019-12-26 01:55:03
农民致富之友(2019年36期)2019-01-13 09:47:39
现代农业科技(2018年10期)2018-11-24 10:40:48
现代园艺(2018年2期)2018-03-15 08:00:00
