
平朔露天煤矿植被恢复区物种天然更新格局及其驱动因素
2016-04-22 06:55:12毕润成吴兆飞宋厚娟白中科郭东罡
环境与可持续发展 2016年2期
张 青 毕润成 吴兆飞 宋厚娟 白中科 郭东罡
(1.山西师范大学生命科学学院,山西 临汾 041004;2.山西大学环境与资源学院,太原 030006;
3.中国地质大学土地科学技术学院,北京 100083;4.国土资源部土地整治重点实验室,北京 100035)
平朔露天煤矿植被恢复区物种天然更新格局及其驱动因素
张青1毕润成1吴兆飞2宋厚娟2白中科3,4郭东罡2
(1.山西师范大学生命科学学院,山西 临汾041004;2.山西大学环境与资源学院,太原030006;
3.中国地质大学土地科学技术学院,北京100083;4.国土资源部土地整治重点实验室,北京100035)
【摘要】以安太堡露天煤矿植被恢复区0.8ha“刺槐+榆树+油松混交模式”长期固定监测样地为研究平台,基于2010年和2015年两次幼苗幼树的调查数据,对植被恢复过程中主要更新物种的数量特征和空间分布格局的变化进行了分析。研究表明:2010-2015年间,样地内幼苗个体数由8587株减少为3673株,幼树个体数由307株减少为172株;从群落水平来看,幼苗在各生境类型下个体数量的分布略有差异。2010年的幼苗在样地四个角和中部呈聚集分布。幼苗个体数量的分布表现为高坡>高谷>山谷>低坡。而2015年呈高坡>山谷>高谷>低坡的趋势。幼树在不同生境类型下的个体数相差不多。无论在群落水平还是物种水平,幼苗幼树高度和生物因子共同影响幼苗幼树的存活。本研究为揭示不同植物配置模式种群的天然更新和物种共存及维持机制提供基础资料,同时为优化复垦土地植被恢复模式和促进矿区土地可持续利用提供理论依据。
【关键词】植被恢复;更新;空间格局;时空动态
安太堡大型露天煤矿大规模的开采,造成了土地资源大面积的损毁和矿区生态环境的安全隐患。关于矿区生态恢复的研究,基本认可植被恢复是矿区损毁土地进行生态恢复的前提和基础[1-2]。利用植物群落自组织原理等相关理论构建起来的人工植物群落,在植被的演替进程中是否可以实现自我更新,直接关系到重建植被是否与当地的环境条件之间形成了协调的动态平衡关系。在树木的生活史中,幼苗到幼树阶段是树木定植的瓶颈阶段,被认为是个体生长最为脆弱、对环境变化最为敏感的时期[3],因此研究人工恢复植物群落在不同阶段的更新幼苗和幼树的生态学特征及其影响因子,对于更好地了解植被恢复过程中物种间同的新方式,进而揭示影响人工重建植被群落动态的根本原因,合理评价人工重建植被的适应性具有重要意义。
采用CTFS(Centre for Tropical Forest Science)的技术规范,于2010年在安太堡露天煤矿排土场植被恢复区建立的固定监测样地,为研究矿区生态恢复过程、物种空间分布格局、群落演替动态等提供了重要的研究平台。采用同样的监测标准,我们于2015年进行了第一次样地复查,探讨幼苗幼树时空动态及其环境限制因子,了解生态恢复过程中人工重建植被物种的新策及新动态。
1研究地概况与研究方法
1.1研究地概况
研究地位于山西省北部的朔州市平鲁区境内的安太堡煤矿南排土场,地理坐标为39°24′ N~39°38′ N,112°11′ E~113°32′ E。该地区属于温带半干旱大陆性季风气候,年平均降水量为428.2mm,年平均气温为5.5℃,≥10℃的年积温为2300~2500℃,无霜期为117d。矿区地带性植被属于草原类型。
安太堡露天煤矿南排土场属于外排土场,最终标高1465m,容量1.16亿m3,排土高度150m,1985年开始排弃,1989年排弃结束,1993年开始植被重建,是安太堡矿生态恢复较早的区域之一,期间除定植期进行浇水外,至今均未采取人工浇水与施肥等管理措施,目前已经形成刺槐(Robinia pseudoacacia)、油松(Pinus tabulaeformis)、榆树(Ulmus pumila)等为主的生物多样性丰富的植被恢复区,基本覆盖了排土场原有的裸露地表,其生态环境得到了有效恢复[4]。
1.2样方设置与调查
刺槐+榆树+油松混交林固定监测样地位于南排土场生态复垦区,样地为长方形(0.8m×10m),面积为0.8ha。整个样地较为平缓,平均海拔1786m。2010年,参照CTFS的方法,使用全站仪将样地划分为80个10m×10m的样方,每个10m×10m的样方又划分为4个5m×5m的小样方。对样方内所有木本植物进行物种鉴定、高度和胸径测量,定位及挂牌标记。
1.2.1幼苗幼树监测
幼苗和幼树调查的标准和方法为:样方内所有胸径<3cm的乔木记为幼苗,3cm≤胸径<7cm 的乔木记为幼树,记录其树种名称、高度、胸径、空间坐标等信息。
1.2.2林下光环境测定
采用半球面影像法(Digital hemisphere photograph)监测林下光环境。该方法利用180°鱼眼镜头拍摄某点在某一高度水平半球面上所有物体的投影照片。当镜头垂直地面向上拍摄时,就可以获得整个天空的半球面照片。本研究使用鱼眼镜头为AF Fisheye Nikkor 16mm f12.8D镜头,机身是Nicon D700,拍摄方向为正北方,拍摄高度为1.3m,全阴天或者太阳升起前及落山后拍摄。在每个10m×10m的样方每条对角线等距离布设三个点,两对角线交叉处不需重复拍摄,每个样方共计5个点,每个点重复拍摄3次,最后选取5个点的平均值作为10m×10m样方的林下光环境特征参数。
1.3数据处理
1.3.1幼苗幼树存活驱动因子的相关性分析
2010年个体数≥100的幼苗幼树到2014年的存活状态与否,1(活)或0(死)作为因变量,使用广义线性混模型(以下简称GLMM),用幼苗幼树高度、生物因素和非生物因素构建了4个GLMM:①幼苗幼树高度模型(幼苗幼树样方和物种作为随机效应,高度作为固定效应);②非生物因子模型(在高度型基础上,只考虑地形和光环境作为固定效应的影响);③生物因子模型(在高度型基础上,只考虑幼苗和大树邻体作为固定效应的影响);④全因子模型(在高度模型基础上,考虑上述所有变量作为固定效应的影响),在群落水平和物种水平上检验幼苗幼树高度、生物因素和非生物因素对幼苗幼树存活的影响及各因子的相对重要性。各模型之间的比较运用赤池信息量准则(Akaike′s Information Criterion,AIC),值越小,代表模型越好,为最优模型。生物因素为每个幼苗样方内每株幼苗的同种和异种幼苗数、每株幼苗的同种和异种大树个体数、以及以目标幼苗为圆心,10m为半径,计算此范围内每株大树的胸高断面积与大树和目标幼苗距离之比,并分同种和异种大树分别计算该比值之和。非生物因素包括海拔、坡度、坡向以及用直射光透射率和散射光透射率代表林下光环境。
1.3.2生境类型的划分
生境类型具体划分方法:根据每个10×10m样方顶点的海拔值,得到每个样方的平均海拔值。凹凸度即是目标样方的平均海拔值与周围所有相邻样方平均海拔的差值,若凹凸度为为正值说明目标样方比周围样方高,反之则低,若为零则高度相等。坡度为实测值。根据每个样方海拔(H)(四个顶点海拔的平均值)和坡度(S)的中值以及凹凸度(C),四种栖息地类型划分标准:①山谷(valley,V):S≤median(S),H
1.3.3光因子数据处理
使用GLA软件(Gap Light Analyser software,version2.0;Frazer et al. 1999)分析半球面图像,所有照片均由同一个人处理,以减少阈值选择的人为误差。获取使用的光环境特征参数包括:冠层开阔度(canopy openness,CO)、叶面积指数(LAI)、入射光特征参数(包括散射光入射率(Trans.Diffuse)、直射光入射率(Trans.Direct)、总辐射入射率(Trans.Total))等。
采用R、Excel 2007对数据进行相关统计分析。
2结果与分析
2.12010-2015年样地内幼苗幼树的时间动态
样地2分析结果表明,2010-2015年5年间,物种数由3种(刺槐、油松和榆树)变为5种,三裂绣线菊和沙棘由于出现新个体而成为2个新增物种(见表1)。
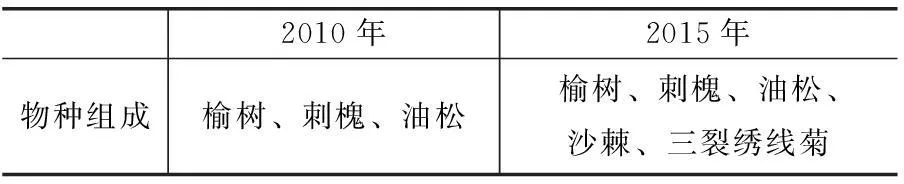
表1 物种组成表
2010-2015年,幼苗由8587株减少至3673株,幼树由307株减少至172株,幼苗幼树死亡量远远大于新增量。5年间榆树幼苗新增数量最多的物种,也是幼苗和幼树死亡量最大的物种,但刺槐幼苗(72.85%)和幼树(51.12%)的死亡率仍大于榆树幼苗(49.84%)和幼树(20.59%)的死亡率。油松只有死亡个体而无新增个体(见表2和表3)。
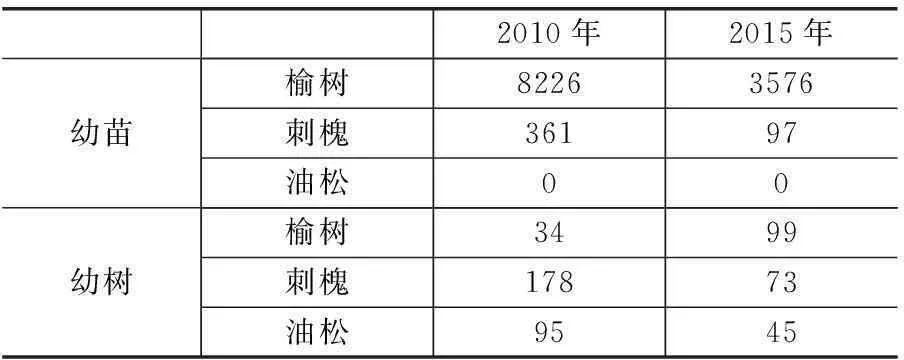
表2 各物种幼苗幼树的数量统计
2.2幼苗幼树在空间上的动态
不同的生境会对不同物种的幼苗幼树生长形成不同的影响,由于样地的地形条件差异,依据平均海拔、凹凸度和坡度将样地划分为高坡、高谷、山谷和低坡4种地形。在样地中不同的地形条件下,各物种的幼苗幼树的出生、死亡和生长变化也存在一定的差异,对三个优势物种幼苗幼树在不同地形条件下的动态与空间分布进行了统计和分析。结果表明,2010年的幼苗聚集在样地中部与东北、东南、西北、西南四个顶点处,以中部最为集中。从生境条件上看,幼苗个体数量的分布情况为高坡>高谷>山谷>低坡。到2015年幼苗仍表现出在样地中部的集中分布,幼苗个体数量的分布情况为高坡>山谷>高谷>低坡。2010年幼树在样地内为均匀分布,在各生境条件下的分布也较为均匀,没有表现出明显的聚集。到2015年幼树的分布依然比较均匀。

表3 各物种幼苗幼树存活动态
2.2.1榆树幼苗幼树的空间分布
2010年的榆树的幼苗聚集在样地中部与东北、东南、西北、西南四个顶点处,以中部最为集中,从生境条件上看,榆树幼苗个体数的分布情况为高坡>高谷>山谷>低坡。到2015年在小范围内聚集分布,榆树幼苗个体数的分布情况为高坡>山谷>高谷>低坡。2010年和2015年由于数量较少,榆树的幼树在样地内呈随机分布。
2.2.2刺槐幼苗幼树的空间分布
2010年刺槐幼苗在样地内分布较均匀,只在个别样方内呈现密集分布,个体数在高坡分布最多,高达130株。到2015年大量幼苗死亡,数量大大减少,在样地内呈现随机分布。2010年与2015年刺槐幼树在样地内都是均匀分布,在高坡和山谷分布比较多。
2.2.3油松幼苗幼树的空间分布
在2010年和2015年两次调查中,我们都未曾发现油松幼苗。2010年油松幼树在样地内呈均匀分布,山谷中的幼树数量最多。2015年油松幼树数量减少,但仍为均匀分布,也是山谷中数量最多。
2.3幼苗幼树存活的驱动因素
幼苗幼树的存活受到多种因子影响,这些因子可大致分为生物因子和非生物因子两类[7]。研究表明,周围邻近个体数是影响乔木幼苗存活的重要因素[8-9]。此外,乔木幼苗高度也是影响幼苗存活的关键因子,幼苗存活概率会随着幼苗高度的增加变大[10-11]。
2.3.1群落水平上幼苗幼树存活与光、地形和生物因素的关系
在群落水平上,影响样地幼苗存活的最优模型为幼苗高度模型,影响幼树存活的最优模型为非生物模型。在影响幼树存活的模型中,坡度与之有较强烈的正相关关系,同时,直射光透射率、散射光透射率、海拔、坡向等非生物因素都与幼树的存活有着密切的关系。但是,幼苗高度的影响也是不容忽视的。这就说明在有着海拔差异的样地2内,不单单是幼树高度,非生物因子对样地内整体幼树的更新与分布也是存在一定影响(表5、6和图1、2)。

表5 群落水平上幼苗存活模型的AIC值
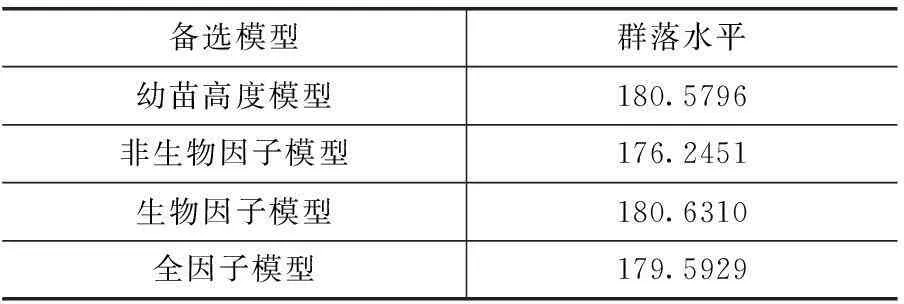
表6 群落水平上幼树存活模型的AIC值

图1 幼苗群落水平
2.3.2物种水平上幼苗幼树存活与光、地形和生物因素的关系
在物种水平上,榆树、刺槐幼苗的最优模型均为生物模型,与此同时,同种大树胸高断面积、同种大树个体数、异种大树胸高断面积、异种大树个体数等生物因子也在不同程度的影响其存活与分布(表7、8 图3、4、5)。

表7 物种水平上幼苗存活模型的AIC值
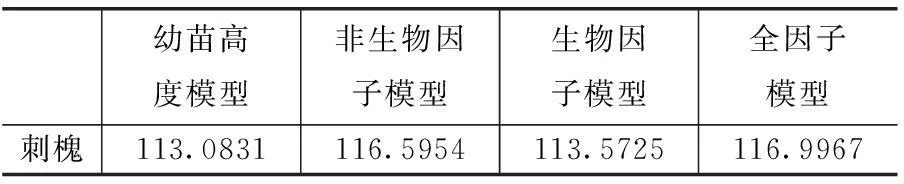
表8 物种水平上幼树存活模型的AIC值
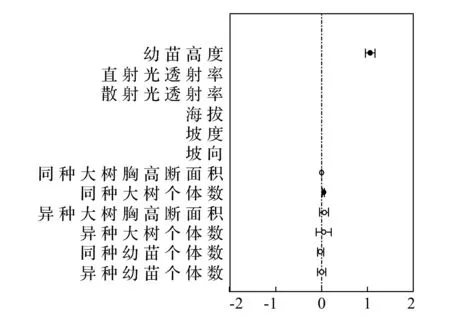
图3 榆树幼苗
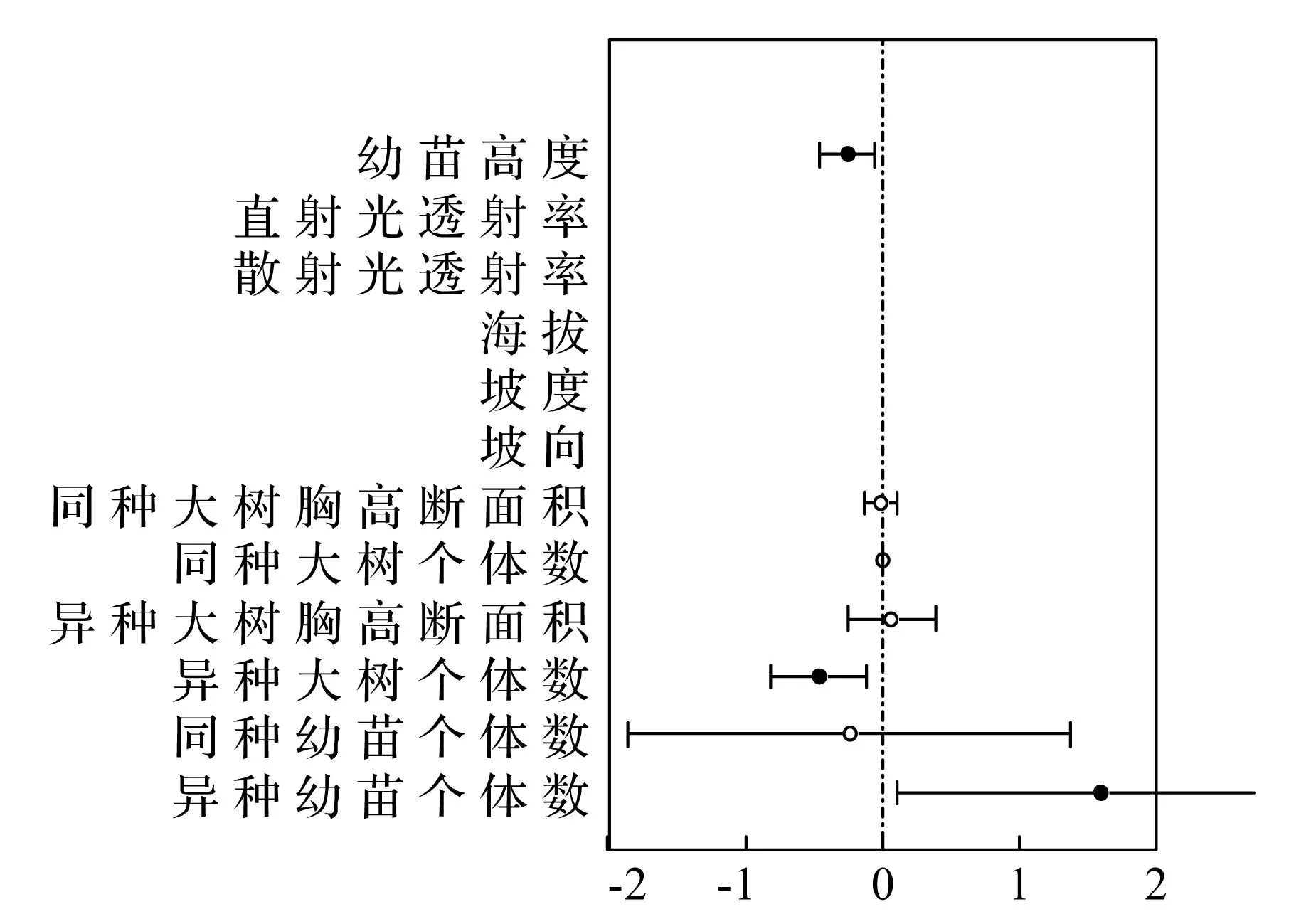
图4 刺槐幼苗

图5 刺槐幼树
3讨论
对样地内2010年调查数据与2015年数据对比的结果表明,无论是幼苗还是幼树,2010年的个体数越多,其死亡数量就越大;榆树无论幼苗还是幼树,其存活率皆大大高于刺槐;更新能力的差异将会使样地内的配置模式从一个平衡状态向另一个平衡状态转换[12],幼苗的存活率是决定各物种在样地中地位的关键因素,但是其过低的存活率成为其实现自我更新的瓶颈,其优势地位可能被样地内其他树种所取代。
空间分布格局是指种群个体在水平空间的配置状况或分布状况,反映了种群个体在水平空间上彼此间的相互关系,是由种群本身生物学特性和环境条件的综合影响所决定的[5],不仅是对过去生态学过程的反映,而且更重要的是对未来生态学过程有非常重要的作用[6]。通过对比观察样地内两年的幼苗幼树空间分布格局图,可以看出2015年主要树种的幼苗幼树分布情况基本与2010年的分布情况保持一致;样地中,2010年分布密集的区域,到2015年分布也是较为密集的;在2010年分布稀疏的区域,2015年分布也不会很多。这也与物种的数量特征相呼应,2010年数量多的,2015年的新增苗也较多,分布区域与之前基本保持一致。
在本文分析的广义线性模型中,幼苗幼树的高度与幼苗幼树的存活有着非常紧密的关联。在样地内,幼苗的高度与幼苗的存活情况呈现出显著的正相关性,虽然在不同的水平上,幼苗高度对幼苗存活的影响的显著程度并非完全一致,但是我们可以发现,幼苗的高度越高,它的存活率就越高,就越不容易死亡。这一结论与匡旭等在长白山阔叶红松林样地中得出的结论类似[7]。
越来越多的研究表明,生物因子和非生物因子是共同影响着幼苗的存活。本研究中,我们发现生物因子在对幼苗幼树的存活中起着更显著的作用,而非生物因子对幼苗幼树存活的影响却相对有限,这一研究结果与Shibata等[13]在日本温带森林群落树木更新影响因子的研究结果一致。但是同时我们也发现,周围同异种大树个体数、胸高断面积以及幼苗幼树的个体数都对幼苗幼树的存活产生着影响,有时起正效应,有时起反作用,可见,个体的表现受到周围邻近个体的限制[14-15],即密度制约在本研究区也是存在的。同时,非生物因素对幼树的影响也较大,海拔、坡度等因素与幼树的存活呈负相关关系,这说明,在该地区,海拔和坡度、坡向在很大程度上限制了幼树的更新。从不同生境类型下的死亡率来看,幼苗死亡率由高到低为高谷>高坡>山谷>低坡,而幼树则表现出了高坡>山谷>高谷>低坡,幼苗幼树均在低坡死亡率最低。幼树的存活于非生物因子当中的坡度存在着显著的负相关关系,坡度愈大,幼树的存活率越低。
本研究只是基于样地两年两次的调查数据,这对于监测人工林的更新状态来说时间还不是很充分。并且在分析影响幼苗幼树的存活因素时,我们选取的评价因子并不全面,例如土壤因子、微地形等更多非生物因子因素,因此,还需要更加长期的定位监测和考虑更多的因素来全面揭示植被恢复区物种的更新机制。
参考文献:
[1]Guo Donggang,Zhao Bingqing,Shangguan Tieliang,et al.Dynamic parameters of plant communities partially reflect the soil quality improvement in eco-reclamation area of an opencast coal mine. Clean-Soil,Air,Water,2013,41(10):1018-1026.
[2]刘卫华,赵冰清,白中科,等.半干旱区露天矿生态复垦土壤养分与植物群落相关分析.生态学杂志,2014,33(9):2369-2375.
[3]Wright S.,Muller-Landau H.,Calderon N.,et al.Annual and spatial variation in seed fall and seedling recruitment in a neotropical forest.Ecology,2005,86:848-860.
[4]李晋川,白中科.露天煤矿土地复垦与生态重建—平朔露天矿的研究与实践.北京:科学出版社,2000.
[5]郭忠玲,马元丹,郑金萍,等.长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究.应用生态学报,2004,15(11):2013-2018.
[6]闫海冰,韩有志,杨秀清,等.华北山地典型天然次生林群落的树种空间分布格局及其关联性.生态学报,2010,30(9):2311-2321.
[7]匡旭.长白山阔叶红松林乔木幼苗动态及其驱动因素.中国科学院沈阳应用生态研究所,2014.
[8]Queenborough,S.A.,D.F.Burslem,N.C.Garwood,et al.Habitat niche partitioning by 16 species of Myristicaceae in Amazonian Ecuador.Plant ecology,2007,192(2):193-207.
[9]Das,A.,J.Battles,P.J.van Mantgem,et al.Spatial elements of mortality risk in old-growth forests.Ecology,2008,89(6):1744-1756.
[10]Svenning,J.C.,T.Fabbro,S.Wright.Seedling interactions in a tropical forest in Panama.Oecologia,2008,155(1):143-150.
[11]Lin,L.,L.S.Comita,Z.Zheng,et al.Seasonal differentiation in density‐dependent seedling survival in a tropical rain forest.Journal of Ecology,2012,100(4):905-914.
[12]杨京平,卢剑波.生态恢复工程技术.北京:化学工业出版社,2002.
[13]Shibata,M.,T.Masaki,H.Tanaka,et al.Effects of abiotic and biotic factors and stochasticity on tree regeneration in a temperate forest community.Ecoscience,2010,17(2):137-145.
[14]Chesson,P.Mechanisms of maintenance of species diversity.Annual review of Ecology and Systematics,2000,31(1):343-366.
[15]Volkov,I.,J.R.Banavar,F.He,S.P.Hubbell,A.Maritan.Density dependence explains tree species abundance and diversity in tropical forests.Nature,2005,438(7068):658-661.
Temporal-spatial Dynamics of Natural Regeneration of Species in Ecological Reclamation of ATB Opencast Coal Mine
ZHANG Qing1BI Runcheng1WU Zhaofei2SONG Houjuan2BAI Zhongke3,4GUO Donggang2
(1.Life Science,Shanxi Normal University,Linfen,Shanxi Province 041004,China;2.College of Environmental and Resources,Shanxi University,Taiyuan 030006,China;3.School of Land Sciences and Technology,China University of Geosciences,Beijing 100083,China;4.Key Lab of Land Consolidation and Rehabilitation,the Ministry of Land and Resources,Beijing 100035,China)
Abstract:A 0.8-ha permanent plot at the eco-reclamation area in Antaibao Opencast Coal mine of China Coal Pingshuo Coal Company Limited was established after reforestation of a mixed plantation of “locust (Robinia pseudoacacia)+elm(Ulmus pumila)+Chinese pine (Pinus tabuliformis)”.Applying the R programming language and based on the two census data in 2010 and 2015,an analysis was made on characteristics of qualities and changes in spatial distribution pattern of the regeneration of main tree species,the law of natural regeneration and drivers of the seedlings and saplings of the plot in 2010-2015.It provides a scientific basis for the biodiversity conservation and artificial vegetation continuing operations in this area. Also, it provides basic information for natural regeneration and species coexistence mechanisms of different plant configuration modes.
Keywords:vegetation restoration;regeneration;spatial pattern;temporal-spatial dynamics
中图分类号:X21
文献标识码:A
文章编号:1673-288X(2016)02-0172-05
通讯作者:毕润成,教授,研究方向为植被生态学
作者简介:张青,硕士,研究方向为植被生态学
项目资助:国土资源部土地整治重点实验室资助的开放基金项目“山西平朔煤矿土地植被恢复物种变化过程研究”(项目编号:20140136)
引用文献格式:张青等.平朔露天煤矿植被恢复区物种天然更新格局及其驱动因素[J].环境与可持续发展,2016,41(2):172-176.
猜你喜欢
天津农业科学(2016年12期)2017-01-11 20:23:42
南水北调与水利科技(2016年6期)2017-01-06 13:10:22
新教育时代·教师版(2016年23期)2016-12-06 04:07:41
现代经济信息(2016年22期)2016-10-26 20:57:26
现代园艺(2016年13期)2016-09-02 09:54:27
科教导刊·电子版(2016年11期)2016-06-03 21:28:27
现代经济信息(2016年6期)2016-05-31 12:36:55
现代经济信息(2016年1期)2016-01-25 01:22:27
