
竞争和景观格局相互作用对外来入侵物种传播影响的动态模拟
2016-04-13 00:45齐相贞林振山刘会玉
生态学报 2016年3期
齐相贞, 林振山, 刘会玉
1 南京师范大学地理科学学院, 南京 210023
2 江苏省环境演变与生态建设重点实验室, 南京 210023
竞争和景观格局相互作用对外来入侵物种传播影响的动态模拟
齐相贞1,2, 林振山1,2,*, 刘会玉1,2
1 南京师范大学地理科学学院, 南京210023
2 江苏省环境演变与生态建设重点实验室, 南京210023
摘要:通过中性景观模型和元胞自动机模型模拟了不同景观格局、不同竞争力和关键种群特征的外来入侵物种的传播动态,模拟结果表明:(1)竞争力强的外来入侵物种,可利用生境面积越大,集聚度越高,越有利于其入侵和传播,而对竞争力弱的外来入侵物种来说,可利用生境面积越小,越分散,越有利于其生存。竞争力强可以有效利用整块的高集聚度的资源,而弱竞争力的外来入侵物种为了逃避土著种的竞争往往趋向于分布在分散的小生境中;(2)可利用生境面积大于50% 时,景观格局集聚度越小,竞争力弱的外来入侵物种适应环境的弛豫时间越长;(3)外来入侵物种的传播与种子产量呈正相关关系,与繁殖年龄呈负相关关系,拟合关系因景观格局和竞争而异;(4)平均迁移距离对外来入侵物种传播的影响最大,在管理过程中应根据不同的目标,不同的景观格局、竞争力和种群特征选择合适的管理措施。
关键词:竞争; 景观格局; 传播; 动态模拟
传播是外来物种入侵过程中最关键的环节之一,它可以使外来入侵物种能够达到高密度和大尺度的空间分布[1- 3]。目前,对外来入侵物种传播过程的研究在实践和理论上都做了很多工作[4- 12],特别是理论模型研究,最早可以追溯到Fisher[4]提出的反应-扩散模型。理论模型可以将植物入侵过程中不同因子结合起来理解、预测和控制入侵,相对其他传统方法更为经济有效[13]。总结模型的发展历程,主要侧重于生长和扩散两个方面。就生长而言,从最初的指数增长、逻辑斯蒂增长到包括生活史阶段等种群统计特征参数的研究。另一方面,从扩散来看,从简单的随机短距离扩散、长距离扩散到各种不同现象、机制扩散模型的发展,对外来物种入侵传播的研究一步步深入。但是,外来入侵物种的传播不仅与生长和扩散本身有关,还与景观格局和种间相互作用联系密切[14- 18]。With[19]从景观生态学的角度比较全面的分析了景观格局对外来入侵物种扩散的影响,Kinezaki等[20]探讨了周期性的生境变化对外来入侵物种传播的影响。Andrew等[21]应用RS和数学模型分析方法探讨扩散时间异质性和景观格局对外来入侵物种传播的影响。Marco等[22]从模型和实例两方面探讨了长短距离传播对外来物种入侵的影响。Brown等[12]分析了斑块化对外来物种入侵传播的影响。Coutts等[23]综合了影响外来物种传播的各种驱动力从入侵管理的角度进行了分析。然而在所有的影响因素中,竞争是一个重要的驱动力[15],大多数研究者对其描述比较简单,表现为对空元胞的占据或对不同生境的喜好程度。实际上,除此之外,植物受竞争影响程度还取决于自身及相邻植物的大小、数量等方面[17,24]。Connie等[25]对44种邻体植物和Lythrumsalicaria目标种及10种邻体植物和Penthorumsedoides目标种进行了种间竞争试验,通过多元线性分析表明,植物个体数量特征与其竞争能力显著相关。因此,基于当前的研究背景,本文拟利用元胞自动机模型,通过景观中性模型构建不同景观格局,探讨不同情景下外来物种的入侵特征,分析在不同景观格局、不同竞争能力、不同种群统计特征的外来物种的入侵动态和入侵的关键驱动因子。
1模型构建
1.1景观格局构建
尽管真实景观异常复杂,但中性模型提供了一套参照系与真实景观相对照。通过研究景观中性模型所产生的景观格局,不仅可以了解景观格局和过程的相关因素,还可以通过控制这些因素来模拟景观格局和过程的变化情况,对于真实景观具有一定的预测和调控能力[26- 27]。本文选用RULE中性景观模型,构建具有不同程度的面积丧失和集聚度(干扰度)的空间结构来模拟真实的景观格局。通过赋值D表示栖息地面积丧失的程度,D在0—1之间变化,D越接近1,栖息地丧失的面积越大,D=0时,表示栖息地未受破坏。用H表示空间聚集度(干扰度),H在0—1之间变化,越接近1,表示景观结构在空间上越聚集(干扰越小);越接近0,表示破碎化程度越高,干扰越大。同时变化D和H,可以产生一系列复杂的具有不同破碎化程度和面积比例的景观格局。本文假设9种景观空间构型:3种不同程度的面积丧失D=0.25、0.5和0.75,同时,每个面积丧失等级均设立了3种不同程度的空间聚集水平(干扰水平)H=0、0.5和1(图1)。其中,图幅大小为27×27。

图1 不同干扰与面积比例的景观格局 Fig.1 Landscape patterns with disturbance and loss in different degrees D:生境丧失比例The proportion of habitat loss; H: 聚集度 Habitat aggregation
1.2元胞自动机模型
本文中,根据图1产生的9种不同干扰和面积比例的景观格局构建27×27的元胞空间。每个元胞都有3种状态,即永久毁坏,空白可占据,以及被某一物种所占据。如果元胞状态为0,则该元胞表示为永久毁坏,不能被任何物种所占据;如果状态为j=1,2,3,…,n(n为本地种的数目),则该元胞被土著种j所占据;如果元胞处于n+1的状态,则表示被外来种所占据;如果元胞处于n+2的状态,则表示该元胞为空白可占据。本文以外来种入侵两本地种组成的土著群落为例进行分析,根据外来种、土著种1和土著种2的初始多度(表1)随机的对元胞进行状态赋值,在引入外来种之前,根据两土著种的初始赋值(表1),对其进行了15a的共存模拟实验,保证了入侵前均衡的入侵环境。
在元胞自动机模型中,元胞是最小的单位,每个元胞最多只被一个成年植株占据,相邻元胞间的距离为1。模型中用到的参数包括:tmax,物种的平均寿命;q,成年个体的存活概率;tm繁殖年龄;n,种子平均产量;fg种子平均萌发概率;Ps,幼苗存活概率;成年植株存活概率为q,即死亡率为1-q。
入侵种和当地种的相互作用可以分为两种情况,一种是直接竞争,即根据元胞周围的临域植株数而定;一种是对空元胞的竞争。对空元胞i的占有概率依赖于在时刻t到达元胞i的种子的数目Si,只要有一个种子萌发并长到幼苗的平均年龄则空元胞即被占据,占有概率为[8]:
Pi=1-(1-Psfg)Si
(1)
式中,fg为种子的萌发概率,Ps幼苗的存活概率。Si表示空元胞i接收的距离该元胞为r的植株产生的种子数,可以通过nf(r)计算。n为植株产生的种子数,f(r)是种子传播的扩散方式,本文根据陈玲玲等[28]的扩散方式,描述长、中、短扩散的扩散方式,即:
(2)
式中,r为理论传播距离,在此为元胞个数。α代表种子的根本性质,其生物学意义与种子散布机制相联系,表示在自然状态下进行长短距离传播的能力,α<1时,曲线呈“粗尾”分布,为典型的长距离传播类型;α>1时,曲线呈“细尾”分布,为典型的短距离传播类型;α=1时,为中距离传播。μ为距离系数,μ越小,可达到的距离越远,密度峰值越小。α和μ共同确定了种子的密度分布(具体见参考文献[28])。
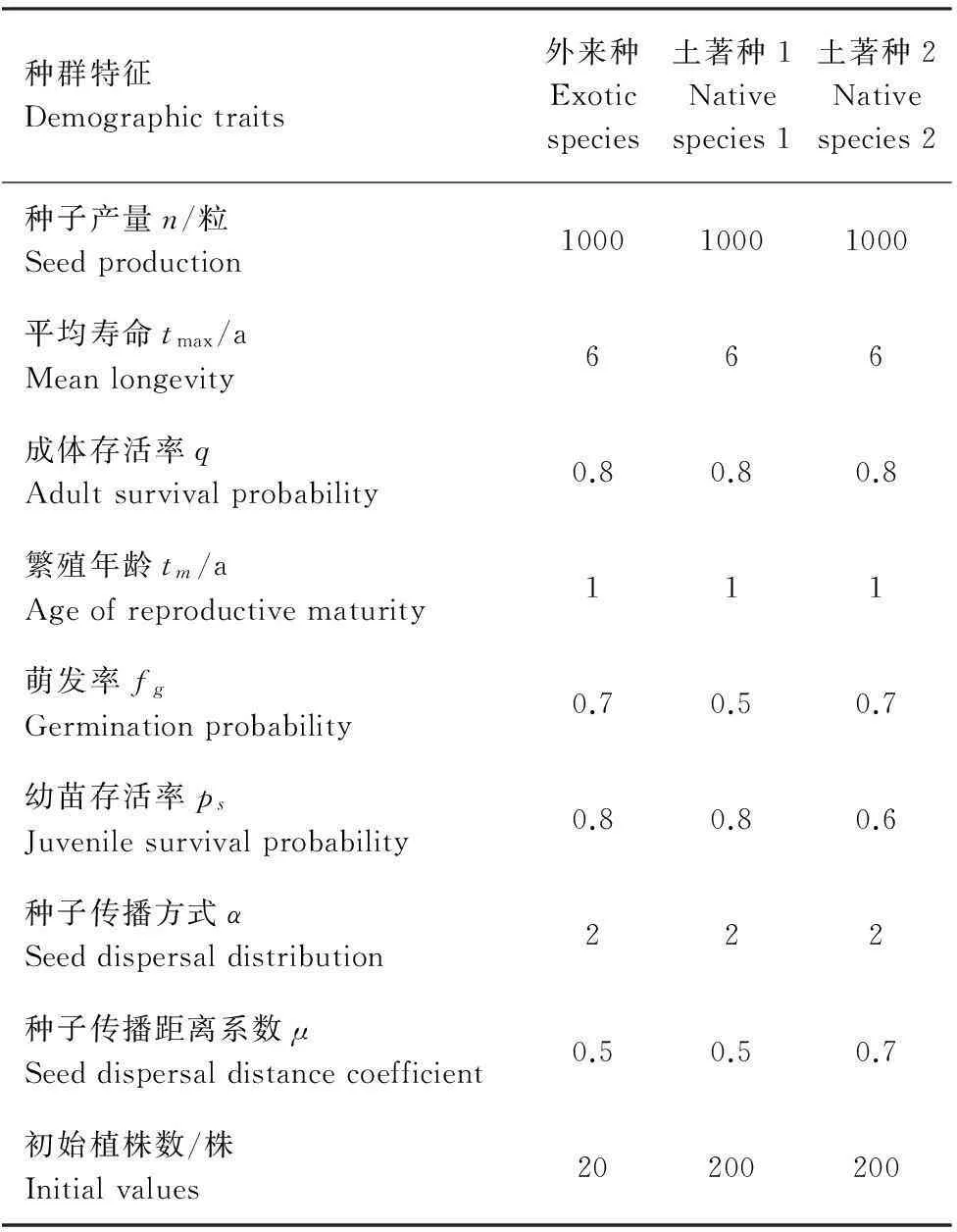
表1 外来入侵物种和土著种初始参数值
多物种相互作用的元胞自动机模型在模拟过程中遵循以下转换原则:
(1)模型是基于个体的,每个元胞单位时间仅仅被一个成年个体所占据;
(2)对空元胞的竞争,根据方程1计算不同物种的占有概率,与独立的随机数比较大小。由于考虑到物种占有概率大的不一定占有该元胞,本文累积各物种的概率和并分别比较各物种所占的比例,然后与随机数比较决定哪个物种占据该元胞。
(3)外来入侵物种和土著物种之间的直接竞争依据不同的竞争能力决定。若以八邻为例,外来入侵物种的竞争能力可以分为8级,同样的道理土著种的竞争能力也相应地分为8级。因此,根据不同的临域植株数量的多少反映外来入侵物种和土著种之间的竞争关系。比如,外来入侵物种周围存在两个土著种时外来种死亡,即为土著种的竞争力强于外来入侵物种。据此,可以划分为外来入侵物种竞争力强、外来入侵物种和土著种竞争力相当和外来入侵物种竞争力弱3种情况。考虑到各种竞争情况的复杂和篇幅所限,在本文中,只考虑土著种周围存在2个外来入侵物种时,土著种即死亡(外来入侵物种竞争力强)和外来入侵物种周围存在2个土著种时,外来入侵物种即死亡(外来入侵物种竞争力弱)两种情景。
本文所有模拟都在matlab 2008a中进行。
2模拟与分析
2.1景观格局对外来物种入侵的影响
景观格局与生态过程相互作用,驱动着景观的整体动态,在图2中不同的景观集聚度和面积丧失对不同竞争能力的外来种入侵的影响不同。对于竞争力强的外来入侵,从图2可以看出:(1)根据可利用生境比例(1-D)从 75%、50% 到 25% 变化过程中,外来入侵物种多度呈现对应的3个层次;(2)同一可利用生境比例下,聚集度越小(斑块越离散),外来入侵物种多度越少,越聚集越有利于外来物种的入侵;(3)聚集度从 1、0.5 到 0,可利用生境比例从 75%、50% 到25% 的变化过程中,外来入侵物种多度间的差距逐渐增大。随着可利用生境的变化,集聚度(干扰度)扮演的角色越显著,可利用生境越少,聚集度对外来物种入侵的影响越大。
而对于竞争力弱的外来物种入侵来说,景观格局对其影响程度要复杂得多,主要表现为:(1)生境占有率 75% 时,外来入侵物种多度随集聚度的变化不明显;(2)生境占有率 50% 时,外来入侵物种多度变化表现为:聚集度1 <聚集度0.5 <聚集度0;(3)生境占有率 25% 时,外来入侵物种多度变化同样表现为:聚集度1 <聚集度0.5 <聚集度0;(4)生境越聚集越不利于外来物种的入侵,尤其是可利用生境比例较低的情况尤为突出。H=0,D=0.75时,外来入侵物种在9种景观格局下多度最大,空间扩展力表现最强。在外来入侵物种竞争力弱的情况下,可利用生境越少越分散,竞争的作用越小,随机性对外来物种的入侵影响越大。

图2 外来物种入侵9种不同景观格局的动态曲线(外来入侵物种竞争力强和外来入侵物种竞争力弱)Fig.2 population dynamics of invasive alien-species in 9 landscape patterns (Strong competitor and poor competitor)
2.2外来物种种子产量对其入侵的影响
图3给出了不同景观格局下,种子产量(繁殖力)从1000到4000的变化对外来入侵物种多度的影响曲线,从图3可以看出,当外来种竞争力较强时,干扰度从1到0的变化过程中,外来入侵物种多度都表现为基本相似的曲线形态,生境破碎度为25%和50%时,弧形曲线逐渐降低,当破碎度为75%时,似直线形曲线分段下降。可利用资源越少、越随机,种子繁殖力对外来物种入侵的影响越小。短距离的扩散过程中,生境面积越少越分散,种子落入破坏生境的概率越大。图3描述了弱竞争力的外来物种的入侵动态,相比而言要复杂很多。从生境破碎度的角度来看,当生境破碎度D=0.25(图3中蓝色曲线)时,外来入侵物种的多度随干扰度和繁殖力的变化而变化。H=1时,外来入侵物种的多度随繁殖能力增强而增多,但当种子数达到4000时,外来入侵物种的多度反而下降。H=0.5时,外来入侵物种的多度在繁殖力为3000时突然下降,繁殖力在4000时外来入侵物种多度增多。H=0时,外来入侵物种的多度相比其他两种干扰度下整体下降,但随着种子数变化呈上升趋势。当生境破碎度增大为50%时(图3中红色曲线),随着干扰度增加,外来入侵物种多度基本呈上升趋势。当生境破碎度继续增大到75%(图3中黑色曲线)时,随着干扰度增强,外来入侵物种多度显著上升。生境破碎和干扰程度增强有利于弱竞争力的外来物种入侵。从干扰程度来看,当H=1(图3中红色、蓝色、黑色点线)时,繁殖力对外来物种的入侵影响不大,物种间的竞争作用占优势。土著种凭借优势的竞争力和初始生境占有率在高集聚度的生境中快速抢占大量空间,弱竞争力的外来种很难利用繁殖力进行抗衡。当H=0.5(图3b中红色、蓝色、黑色实线)时,繁殖力变化对外来物种的入侵影响比较复杂。当H=0(图3b中红色、蓝色、黑色虚线)时,繁殖力增大则外来物种入侵增强,此时物种在离散随机的生境中很难相遇,竞争作用退居次要作用。
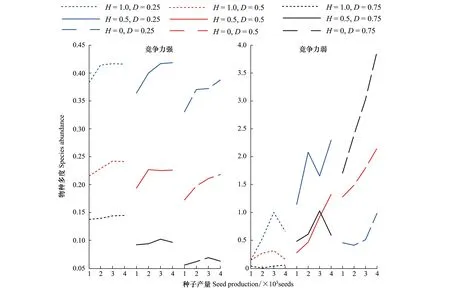
图3 种子产量对外来物种入侵的影响(竞争力强和竞争力弱)Fig.3 Effect of seed production on exotic species invasions (Strong competitor and poor competitor)
Marco等[8,29]通过对外来入侵物种Gleditsiatriacanthos在阿根廷中部山地森林群落的入侵模拟发现,外来入侵物种的传播速度随着种子产量的变化呈对数曲线变化。下图给出了强竞争力下外来入侵物种在不同景观格局下与繁殖力(种子产量)之间的关系并进行了曲线拟合(图4),拟合的相关系数均达到90%以上。从图中可以看到大部分情况下外来入侵物种的多度与种子产量之间是对数拟合关系,但在H=1,D=0.75;H=0.5,D=0.5;H=0.5,D=0.25和H=0,D=0.25情况下对数拟合关系相关系数较低,二次多项式能更好的反映拟合关系。
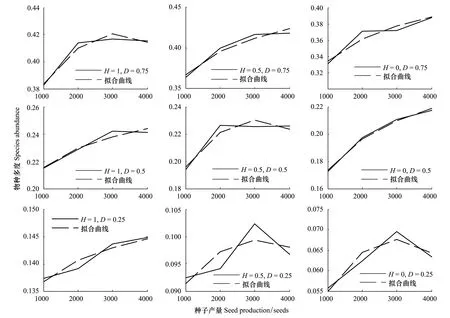
图4 种子产量与外来物种入侵的拟合关系Fig.4 Fitting curve of seed production and exotic species invasions
2.3外来物种平均扩散距离对其入侵的影响
平均扩散距离变化对外来物种入侵的影响比较有规律性,对强竞争力的外来入侵物种来说,当生境毁坏相同时(图5,蓝色曲线系列,红色曲线系列和黑色曲线系列),随着干扰程度变强,平均扩散距离变大对外来物种入侵的影响越强烈,可利用资源越多,平均扩散距离对外来物种入侵的影响越显著。当干扰度相同时(图5a,点线系列,实线系列和虚线系列),可利用资源越丰富,对平均扩散距离的响应越显著。对弱竞争力的外来入侵物种来说,平均扩散距离的影响较强竞争力的外来入侵物种更剧烈。强竞争力的外来入侵物种随着平均扩散距离的增大呈曲线凸形缓慢上升,而弱竞争力的外来入侵物种则表现为曲线凹形快速升高。平均扩散距离对弱竞争力的外来物种来说更有利于其入侵,特别是对生境破坏严重且分散的景观格局来说(H=0,D=0.75),更有利于外来物种的入侵传播(图5)。

图5 平均扩散距离对外来物种入侵的影响(竞争力强和竞争力弱)Fig.5 Impact of mean dispersal distance on exotic species invasion (Strong competitor and poor competitor)
本文主要研究短距离扩散,即α=2的情况,主要表现为平均扩散距离r等于1—4个元胞空间的距离,在聚集度高的景观格局中,即使4个元胞的扩散距离在可利用生境面积大的情况下在很大程度上也会落在适宜的生境中,而对于可利用生境面积较小的景观中,落入毁坏元胞中的概率大很多。对弱竞争力的外来入侵物种来说,平均扩散距离越大越有利于其逃脱土著种的强竞争力。
2.4外来物种繁殖年龄对其入侵的影响
随着繁殖年龄的增大,外来入侵物种多度基本上都表现为迅速降低的状态。强竞争力的外来物种入侵表现的规律性较强(图6),随着繁殖年龄的增加和可利用生境的减少迅速降低。对于弱竞争力的外来入侵物种(图6),当繁殖年龄增大到4龄时,外来入侵物种根本无法定殖成功,因此在图中本文只选取了繁殖年龄从1龄到3龄的变化。外来入侵物种竞争力弱更加剧了繁殖年龄增大带来的繁殖体压力,外来入侵物种的多度随着繁殖年龄变化的更加尖锐。

图6 繁殖年龄变化对外来种入侵的影响(强竞争力和弱竞争力)Fig.6 Effect of average reproduction age on exotic species invasions (Strong competitor and poor competitor)
3讨论
可利用生境面积大小对外来入侵物种的传播动态影响很大,能够调节景观格局对传播的影响。很多研究者发现[19,30- 32],当可利用生境低于某一临界阈值,外来入侵物种的传播能力会显著降低。在模拟中发现,对于强竞争力的外来入侵物种来说,在各种集聚度水平上随着可利用生境面积的减少,外来入侵物种多度下降明显。而且,集聚度越小,干扰度越大,下降幅度越大(图7a)。生境越聚集,可利用生境面积越大,对竞争力强的外来入侵物种越有利。换言之,在短距离扩散条件下,竞争力强可以更方便、更有效的利用整块的高集聚度的资源。但弱竞争力的外来入侵物种在不同的景观格局下随着可利用生境的变化却表现出不同的特征。当聚集度相对较大时(H=1,0.5),随着可利用生境的减少,外来入侵物种的多度稍微上扬后快速减少;在随机分布的景观格局中(H=0,聚集度减小,干扰度增强),可利用生境越小,扩散相对来说会有所增强(图7b)。竞争力弱的外来入侵物种为了逃避土著种的竞争往往会趋向于分布在分散的小生境中。随机分布的、可利用生境面积小的景观格局在一定程度上削弱了土著种的强竞争力,增加了外来入侵物种的生存空间。
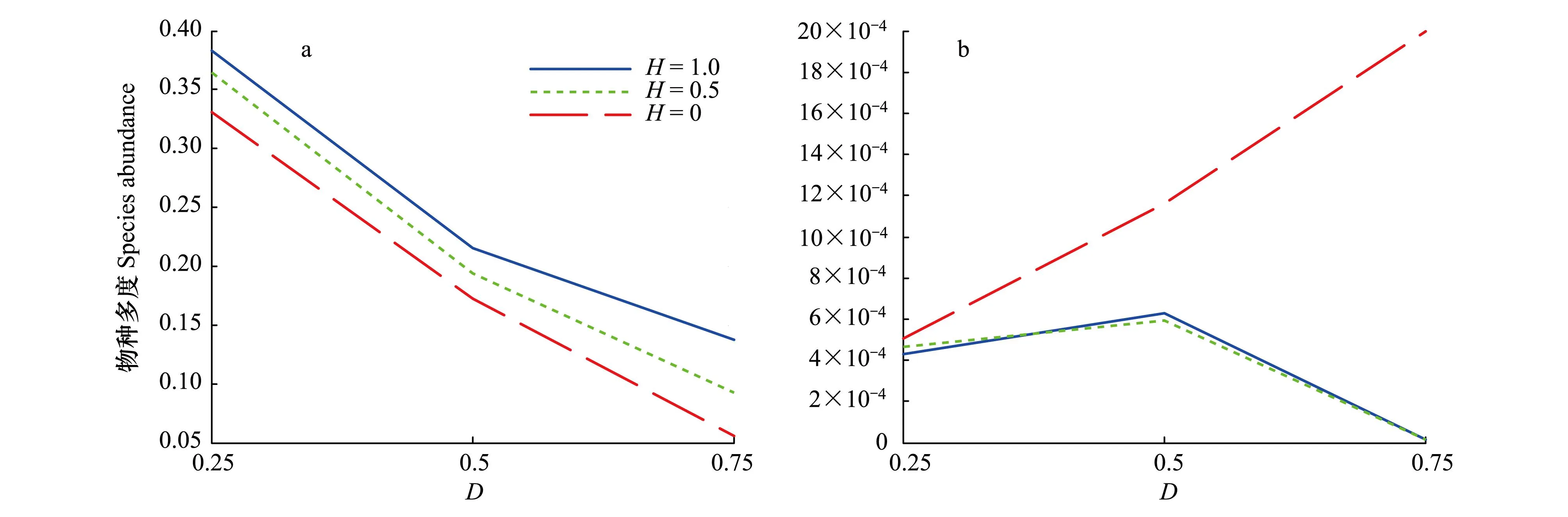
图7 外来入侵物种对集聚度的响应Fig.7 Responses of exotic species to the clumping degree of landscape
集聚度是影响外来物种入侵的另一个重要因子。当外来入侵物种竞争力强,可利用生境面积固定时(图2),外来入侵物种的多度随时间变化逐渐增加,集聚度越大,外来入侵物种的多度越大,传播越迅速。当外来入侵物种竞争力较弱时,外来入侵物种随着集聚度的变化表现较复杂。从图2中可以看出,随时间变化,外来入侵物种的多度基本上表现出较有规律的变化,在前两年的时间内快速增加,之后经过大致2—3a的弛豫时间,平行发展,然后快速下降。当可利用生境面积为75%时,在前两年的时间外来种快速上升,经过2—3a的弛豫时间外来入侵物种的多度表现为基本平行(H=1)或上扬(H=0)或下降(H=0.5)的状态,之后快速下降。当可利用面积为50%时,同样在前两年的时间外来入侵物种的多度快速上升。集聚度H=1时,经过大约1年的弛豫时间,外来入侵物种的多度即快速降低。当集聚度H=0.5时,外来入侵物种没有发生任何迟滞时间即快速下降。当集聚度H=0时,外来入侵物种经过两年多的弛豫时间后快速下降。当可利用面积为25%时,外来入侵物种在最初两年的稍微增长后几乎没有经过任何弛豫时间即缓慢下降(H=1,0.5);当H=0时,外来入侵物种经过1年的弛豫时间后缓慢上升然后缓慢下降。整体来看,在前两年的时间内由于可利用生境足够多,外来入侵物种能够自由扩散。之后H=0时,外来入侵物种的多度缓慢壮大但最终难以抵抗土著物种的强竞争力迅速下降;当H=1和0.5时,外来入侵物种的多度在与土著物种交锋过程中缓慢下降后再突然减小。可利用生境面积大于50%的情形下,集聚度越小,弛豫转换时间越长。
高繁殖力是影响外来物种入侵的关键因素,Marco等[8,29]认为外来入侵物种的传播速度与种子产量呈对数曲线变化。本文分析了强、弱竞争能力的外来入侵物种在不同景观格局下与繁殖力(种子产量)之间的关系并进行了曲线拟合(图4),外来入侵物种竞争力强时,大部分情况下外来入侵物种的多度与种子产量之间是对数拟合关系,但在H=1,D=0.75;H=0.5,D=0.5;H=0.5,D=0.25和H=0,D=0.25情况下对数拟合关系相关系数较低,二次多项式能更好的反映拟合关系。外来入侵物种竞争力弱时(由于篇幅所限,不再给出图形),两者的关系更为复杂(图3)。高繁殖力是外来物种能够成功入侵的一个重要特征,Marco等人的研究主要限于特定的物种和生境特征,并不具有普适性。繁殖力对外来物种入侵的影响还要受到景观格局、竞争力等的影响,它们之间的关系只能说存在非线性关系。繁殖年龄是影响外来物种入侵的另一个重要参数,从本文研究中可以看出,繁殖年龄与传播能力之间呈现负相关关系,而且随着繁殖年龄的增大,外来入侵物种的多度变化更为剧烈。在对外来入侵物种风险控制的实际操作中,很少考虑增大繁殖年龄的方式控制外来物种入侵,这主要基于投资和回报的考虑。从物种本身的能量平衡来讲,繁殖年龄增大会削弱其对后代的投资,增强其自身竞争能力,即从r-对策者转变为K-对策者,但这还与繁殖体压力等关系密切。

图8 种子产量和平均扩散距离对不同景观格局的响应 Fig.8 Responses of seed production and mean dispersal distance to various landscape patterns
控制外来入侵物种传播的方式一般采取两种方式,通过减弱其生长力(r)或者通过改变生境结构如建立廊道等减弱其在空间上的扩散潜力。但是,需要注意的是,对于弱竞争力的外来入侵物种来说,增加廊道,增加景观格局破碎度并不一定是一个好的管理方法。比较了种子产量和平均扩散距离变化对不同景观格局变化的响应(图8),当种子数n=3000,平均扩散距离r=3时,在景观破碎度为75%的景观格局中变为种子数为n=2000,1000,平均扩散距离r=2,景观破碎度不变,但景观干扰度变为0.5和0时,外来入侵物种的多度反而增加,即减弱生长力和增加破碎度并不能抑制扩散反而加快了外来入侵物种的传播。Marvier等[13]认为保护栖息地对抵御外来物种入侵十分有效。根据模拟结果,保护栖息地还需要具体情况具体处理,不同的景观格局、竞争力、种群特征和保护目标,需要不同的考虑。与此同时,通过对不同种子产量、繁殖年龄及平均迁移距离的组合模拟,发现迁移能力对外来入侵物种的传播影响最大,在管理过程中要特别注意。
本文研究了外来入侵物种和土著物种在不同竞争能力对比下的入侵动态,并且分析了景观格局和种群特征等对外来入侵物种传播的影响,对外来入侵物种传播控制与管理提供了一定的理论指导。当然,本文对种子的扩散方式只考虑了短距离扩散对外来入侵物种种群动态的影响,在以后的工作中还要继续深入研究外来入侵物种在长距离扩散条件下的种群动态变化。
参考文献(References):
[1]Caswell H, Lensink R, Neubert M G. Demography and dispersal: life table response experiments for invasion speed. Ecology, 2003, 84(8): 1968- 1978.
[2]Barlow N D, Kean J M. Resource abundance and invasiveness: a simple model. Biological Invasions, 2004, 6(3): 261- 268.
[3]王雅男, 万方浩, 沈文君. 外来入侵物种的风险评估定量模型及应用. 昆虫学报, 2007, 50(5): 512- 520.
[4]Fisher R A. The wave of advance of advantageous genes. Annals of Eugenics, 1937, 7(4): 355- 369.
[5]Skellam J G. Random dispersal in theoretical populations. Biometrika, 1951, 38(1/2): 196- 218.
[6]Clark J S, Lewis M, Horvath L. Invasion by extremes: population spread with Variation in dispersal and reproduction. The American Naturalist, 2001, 157(5): 537- 554.
[7]Fagan W F, Lewis M A, Neubert M G, van den Driessche P. Invasion theory and biological control. Ecology Letters, 2002, 5(1): 148- 157.
[8]Marco D E, Páez S A, Cannas S A. Species invasiveness in biological invasions: a modelling approach. Biological Invasions, 2002, 4(1/2): 193- 205.
[9]Cannas S A, Marco D E, Páez S A. Modelling biological invasions: species traits, species interactions, and habitat heterogeneity. Mathematical Biosciences, 2003, 183(1): 93- 110.
[10]Arim M, Abades S R, Neill P E, Lima M, Marquet P A. Spread dynamics of invasive species. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(2): 374- 378.
[11]Dauer J T, Mortensen D A, Vangessel M J. Temporal and spatial dynamics of long-distance Conyza Canadensis seed dispersal. Journal of Applied Ecology, 2007, 44(1): 105- 144.
[12]Brown R G, James A F, Pitchford J W, Plank M J. Habitat fragmentation: simple models for local persistence and the spread of invasive species. Journal of Theoretical Biology, 2012, 310: 231- 238.
[13]Marvier M, Kareiva P, Neubert M G. Habitat destruction, fragmentation, and disturbance promote invasion by habitat generalists in a multispecies metapopulation. Risk Analysis, 2004, 24(4): 869- 878.
[14]With K A. Assessing the risk of invasive spread in fragmented landscapes. Risk Analysis, 2004, 24(4): 803- 815.
[15]Arii K, Parrott L. Examining the colonization process of exotic species varying in competitive abilities using a cellular automaton model. Ecological Modelling, 2006, 199(3): 219- 228.
[16]刘会玉, 林振山, 齐相贞, 沈竟. 基于景观中性模型的不同竞争力外来种入侵对栖息地毁坏的响应. 生态环境学报, 2011, 20(11): 1638- 1646.
[17]Thébault A, Stoll P, Buttler A. Complex interactions between spatial pattern of resident species and invasiveness of newly arriving species affect invasibility. Oecologia, 2012, 170(4): 1133- 1142.
[18]李乐, 骆争荣, 李琼, 胡志勇, 丁炳扬. 温州地区黑荆树入侵群落的竞争与动态. 生态学报, 2009, 29(12): 6622- 6629.
[19]With K A. The landscape ecology of invasive spread. Conservation Biology, 2002, 16(5): 1192- 1203.
[20]Kinezaki N, Kawasaki K, Shigesada N. Spatial dynamics of invasion in sinusoidally varying environments. Population Ecology, 2006, 48(4): 263- 270.
[21]Andrew M E, Ustin S L. The effects of temporally variable dispersal and landscape structure on invasive species spread. Ecological Applications, 2010, 20(3): 593- 608.
[22]Marco D E, Montemurro M A, Cannas S A. Comparing short and long-distance dispersal: modelling and field case studies. Ecography, 2011, 34(4): 671- 682.
[23]Coutts S R, van Klinken R D, Yokomizo H, Buckley Y M. What are the key drivers of spread in invasive plants: dispersal, demography or landscape: and how can we use this knowledge to aid management?. Biological Invasions, 2011, 13(7): 1649- 1661.
[24]Weiner J. Neighbourhood interference amongstPinusrigidaindividuals. Journal of Ecology, 1984, 72(1): 183- 195.
[25]Gaudet C L, Keddy P A. A comparative approach to predicting competitive ability from plant traits. Nature, 1988, 334(6179): 242- 243.
[26]Gardner R H. RULE: A program for the generation of random maps and the analysis of spatial patterns // Klopatek J M, Gardner R H, eds. Landscape Ecological Analysis: Issues and Applications. New York: Springer-Verlag, 1999.
[27]伍海峰, 李月辉, 李娜娜. 中性景观模型在景观生态学中的应用和发展. 生态学杂志, 2012, 31(12): 3241- 3246.
[28]陈玲玲, 林振山, 何亮. 风传草本植物种子空间传播新模型. 生态学报, 2010, 30(17): 4643- 4651.
[29]Marco D E, Páez S A. Invasion ofGleditsiatriacanthosinLithraeaternifoliamontane forests of central argentina. Environmental Management, 2000, 26(4): 409- 419.
[30]Collingham Y C, Huntley B. Impacts of habitat fragmentation and patch size upon migration rates. Ecological Applications, 2000, 10(1): 131- 144.
[31]King A W, With K A. Dispersal success on spatially structured landscapes: when do spatial pattern and dispersal behavior really matter?. Ecological Modelling, 2002, 147(1): 23- 39.
[32]Matlack G R, Monde J. Consequences of low mobility in spatially and temporally heterogeneous ecosystems. Journal of Ecology, 2004, 92(6): 1025- 1035.
Dynamic modeling of the interactive effects of competition and landscape patterns on the spread of exotic species
QI Xiangzhen1,2, LIN Zhenshan1,2,*, LIU Huiyu1,2
1SchoolofGeographyScience,NanjingNormalUniversity,Nanjing210023,China2JiangsuKeyLaboratoryofEnvironmentalChangeandEcologicalConstruction,Nanjing210023,China
Abstract:Invasive exotic species are increasingly recognized as important drivers of ecological change. Understanding the persistence and growth of natural populations in environments prone to disturbance is very important for both conservation of threatened species and control of invasive species. In this study, neutral landscape and spatially explicit cellular automata models are employed to numerically analyze the interactive effects of key demographic traits, competitive abilities, and landscape patterns on the spread dynamics of exotic species. Available habitat was shown to be of great importance in spread dynamics and mediating the effects of landscape structure. For ‘strong’ invasive species, spread and invasion are facilitated by large areas of highly clumped available habitat. In contrast, for ‘weak’ alien species, scattered small patches of habitat facilitate survival. Strong competitors can efficiently exploit the massive clumps of resources, while poor competitors tend to occupy disrupted niches. Aggregation is another critical factor affecting exotic species invasions, especially for weak nonnative competitors. Exotic species abundance increased quickly in the first two years, before plateauing for 2—3 years, after which it decreased rapidly. In a scenario where more than 50% of the habitat is available, the lower habitat aggregation leads to longer lag times in which weak exotic species adapt to the new environment. The spread of invasive exotic species was positively related to fecundity and negatively related to reproductive age. The best fitting curve for this relationship varies with landscape patterns and competitive ability. For strong competitors, spread is generally logarithmically related to fecundity or reproductive age, but in some cases, quadratic polynomial relationship can provide a better fit. For poor competitors, the relationships among exotic species spread, fecundity and reproductive age are complex. Mean dispersal distance is the key determinant of the spread of exotic species. Compared to the gradual increase in strong exotic species, mean dispersal distance has a greater effect on the spread of weak exotic species. The abundance of inferior competitors rapidly increased as mean dispersal distance increased. Preventing invasive alien plants from reaching new areas should be the most cost effective means of control. In general, two methods can be applied, targeting demographic traits such as fecundity, reproduction, and survival, or managing landscape structure, such as corridors for spread. However, increasing landscape fragmentation will not be an effective method for controlling invasion by weak exotic species. According to our simulation results, habitat conservation needs to be tailored to each specific situation. For example, the abundance of weak exotic species increases when landscape aggregation, fecundity, and mean dispersal distance decrease. Therefore, management strategies should be modified to suit different targets with different competitive and demographic characteristics, and specific landscape scenarios. In this study, we only considered the spread dynamics of invasive species with short-distance dispersal. Further studies on the effect of long-distance dispersal on the invasion and spread of exotic species are required.
Key Words:competition; landscape patterns; spread; dynamic modeling
DOI:10.5846/stxb201403310600
*通讯作者
Corresponding author.E-mail: linzhenshan@njnu.edu.cn
收稿日期:2014- 03- 31; 网络出版日期:2015- 06- 11
基金项目:国家自然科学基金项目(40901034, 31470519); 江苏高校优势学科建设工程资助项目(164320H101)
齐相贞, 林振山, 刘会玉.竞争和景观格局相互作用对外来入侵物种传播影响的动态模拟.生态学报,2016,36(3):569- 579.
Qi X Z, Lin Z S, Liu H Y.Dynamic modeling of the interactive effects of competition and landscape patterns on the spread of exotic species.Acta Ecologica Sinica,2016,36(3):569- 579.
猜你喜欢
现代企业文化·综合版(2017年5期)2017-06-14
环球人文地理·评论版(2016年5期)2017-01-03
中国记者(2016年6期)2016-08-26
湖北农业科学(2015年17期)2015-10-09
人生十六七(2015年26期)2015-08-22
小说月刊(2015年9期)2015-04-23
营销界(2015年22期)2015-02-28
绿色科技(2014年4期)2015-01-06
农业科技与装备(2014年3期)2014-11-28
