
油茶幼林复合经营系统中3种农作物的生长节律研究
2016-04-12 05:36陈志伟龙梦琪何小勇葛永金张日清
浙江林业科技 2016年6期
陈志伟,龙梦琪,何小勇,葛永金,张日清*
(1. 中南林业科技大学,湖南 长沙 410002;2. 浙江省丽水市林业科学研究院,浙江 丽水 323000)
油茶幼林复合经营系统中3种农作物的生长节律研究
陈志伟1,2,龙梦琪1,何小勇2,葛永金2,张日清1*
(1. 中南林业科技大学,湖南 长沙 410002;2. 浙江省丽水市林业科学研究院,浙江 丽水 323000)
2012年3月在浙江丽水市龙泉市兰巨乡仙仁村土地整理后种植1年生油茶(Camellia oleifera)嫁接苗,2015年5月分别套种落花生(Arachis hypogaea)、大豆(Glycine max)和稻(Oryza sativa)。7月起采用整株挖掘取样法和图像分析法,测定生长期套种农作物的生物量,分析其生长节律。结果表明,3种农作物高度最高为稻,收获时可达150.23 cm;生物量累积最高为大豆,收获时达769 g;单株根系长度、须根总长、根表面积最大值均为大豆,收获时分别为33.33 cm,875.04 cm,174.34 cm2。3种农作物中大豆具有最佳的根系以应对油茶幼林复合系统中潜在的根系竞争,且大豆根系腐化后可提高油茶幼林的耕性;3种作物生物量在器官中的积累规律趋于一致,茎和叶所占生物量最高,可达61.6%~68%;根所占生物量最低,约占总生物量的6.6%~14.7%。
油茶幼林;复合经营系统;生长节律;生物量
油茶(Camellia oleifera)属常绿树种,根系较发达,耐干旱瘠薄,抗低温冻害,且适地较广,是我国特有的木本油料树种之一,我国栽培油茶历史逾2000 a[1]。油茶是一种长寿的木本油料树种,种植后收益多年且收获期长。但在生产实践中,油茶幼苗在种植2~3 a后开始开花,5 a后有一定产量,8 a后才能进入丰产期[2],且油茶幼林的抚育管理需要大量人力及物力。因此,产生了油茶幼林管理粗放或荒芜,土壤肥力大量流失,油茶树体生长不佳,结果期推迟,结果率下降等一系列问题,严重制约了油茶产业的发展[3]。
丽水市是浙江油茶的主产区,新造油茶幼林大部分种植在新整理的土地上,有效耕作层薄,立地条件较差,遇暴雨天气冲刷极易造成水土、养分流失。在油茶幼林地与落花生(Arachis hypogaea)、大豆(Glycine max)、稻(Oryza sativa)等农作物进行农林复合经营,不仅符合生态经营的要求,而且能有效改善土壤结构,提高土壤养分含量,从而为油茶树体的生长发育提供充足的营养,同时还能够提高单位面积内土地利用率,增加油茶种植户的经济收入,达到以短养长、以耕代抚的目的。
目前对以油茶为主的农林复合经营系统的研究主要集中在油茶复合系统微环境、油茶树体生物量、土壤理化性状等方面,或仅研究复合系统中农作物产量比较其经济效益[4~16],极少涉及套种作物的生长节律、生物量及根系生长规律。本研究对丽水地区油茶幼林复合经营系统中的3种不同作物生物节律、生物量进行了比较研究,目的是探讨油茶幼林复合系统的能量流、物质流,调控生产结构及建立持续稳定的油茶幼林复合经营模式。
1 材料与方法
1.1 试验地概况
试验地设置在浙西南龙泉市兰巨乡油茶基地,处于浙江省和福建省结合部,27°25′ ~ 28°5′ N,118°41′ ~120°26′ E。中亚热带季风气候,中山、丘陵地貌,温暖湿润,年平均气温11.5~18.3℃,年均降水量1 838 mm,年均日照时数1 769 h,无霜期180~280 d。油茶幼林地为低丘缓坡开发地,砖红壤,土层厚度在60 cm。2012年3月种植‘长林40号’(良种编号:国S-SC-CO-011-2008)1年生油茶嫁接容器苗,购自浙江省定点苗圃,苗高8.133±1.325 cm,地径1.643±0.385 cm,株行距2.5 m×2.5 m,种植面积3.33 hm2。2012-2015年油茶幼林抚育管理依照丽水市地方标准《油茶山地栽培技术规程》(DB3311/T12-2013)执行。每年施肥3次,冬季分别施有机肥5~10 kg•株-1、复合肥1.0 kg /株;早春每株施尿素0.15 kg,磷钾肥1.0 kg;夏季在春梢停止生长后每株施过磷酸钙0.2 kg,氯化钾0.1 kg。2015年7月油茶幼林平均高度180±0.6 cm,平均地径4.7±0.4 cm,东西冠幅147±0.7 cm,南北冠幅153±0.5 cm,2015之前均摘除花芽控制结果,保持林分的营养生长。落花生、稻、大豆均为地方品种。共设置3个试验区,每个试验区分3个试验小区,每个试验小区面积为667 m2,分别种植落花生、大豆、稻,每穴播种3~4粒,株行距25 cm×25 cm。供试落花生、稻、大豆于2015年5月中旬播种,土壤、水分、肥料和病虫害等管理一致。
1.2 测定方法
在各试验小区内设置10 m×50 m的固定标准样地测定作物生物量。2015年7月20日起,每隔15 d(如遇特殊天气提前或延后)取样一次。落花生收获日期为2015年9月10日,大豆收获日期为9月28日,稻收获日期为10月13日。
作物生物量的测定。在标准样地1 m×1 m小样方内采取整株挖掘法将作物全株挖起。测量小样方内的作物株数、平均高度、根系长度。将作物置入85℃烘箱烘至恒重,进行地上部分、地下部分干重测定,测定结果换算成单位面积的数值。整株挖掘后的根系清水洗净用Microtek Scan Wizard EZ扫描仪获取根系扫描图像,并用万深LA-S根系分析系统进行落花生、大豆、稻的总根长、根径、根表面积分析。
1.3 数据分析
作物相对生长速率(RGR)和生长率(CGR)计算:
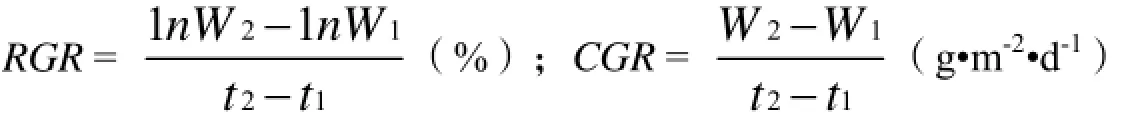
式中,t1,t2为测定日期, W1,W2分别为t1,t2时单位面积干物质量(g•m-2)。试验数据均采用Excel分析制图。
2 结果与分析
2.1 作物株高和地上生物量的变化规律
图1A反映了油茶幼林复合系统内作物高生长变化规律。与7月20日相比,作物采收时落花生、稻、大豆株高分别增长了57%,125%,56%。稻增长速度最快,落花生、大豆在整个生长周期内高生长缓慢且明显低于稻,采收时高度分别为51.17 cm,74.95 cm,处在油茶-农作物复合系统较低层,与油茶-稻复合系统相比,油茶-落花生、油茶-大豆复合系统内拥有良好的通风、光照条件,有利于油茶及落花生、大豆良好生长。稻生长迅速,采收时植株高度与油茶差不多,在营养和光照等方面可能会与油茶形成竞争关系,微环境通风不良和冠层内光照郁闭容易滋生杂草及病虫害。
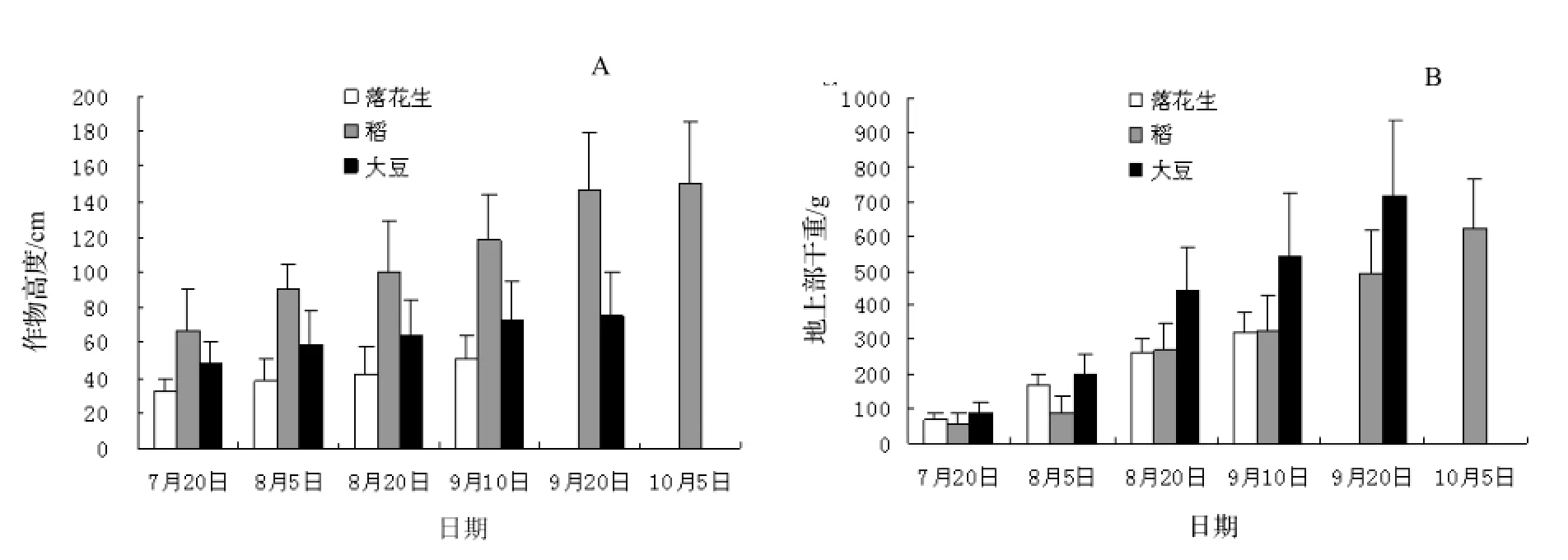
图1 3种农作物株高、地上生物量变化规律Figure 1 Variation of height and aboveground biomass of the tested crops
图1 B反映了落花生、稻、大豆地上生物量变化规律。随着时间的推移各农作物总生物量呈递增趋势。对3种作物采收时生物量比较发现,地上总生物量大小:大豆>稻>落花生。与7月20日相比,落花生、稻、大豆总生物量分别增长了599%,835%,405%。其中,大豆地上部分总生物量累积最大,为717.9 g,分别为采收时落花生、稻地上生物量的2.22倍、1.16倍。
2.2 农作物根系生长规律比较
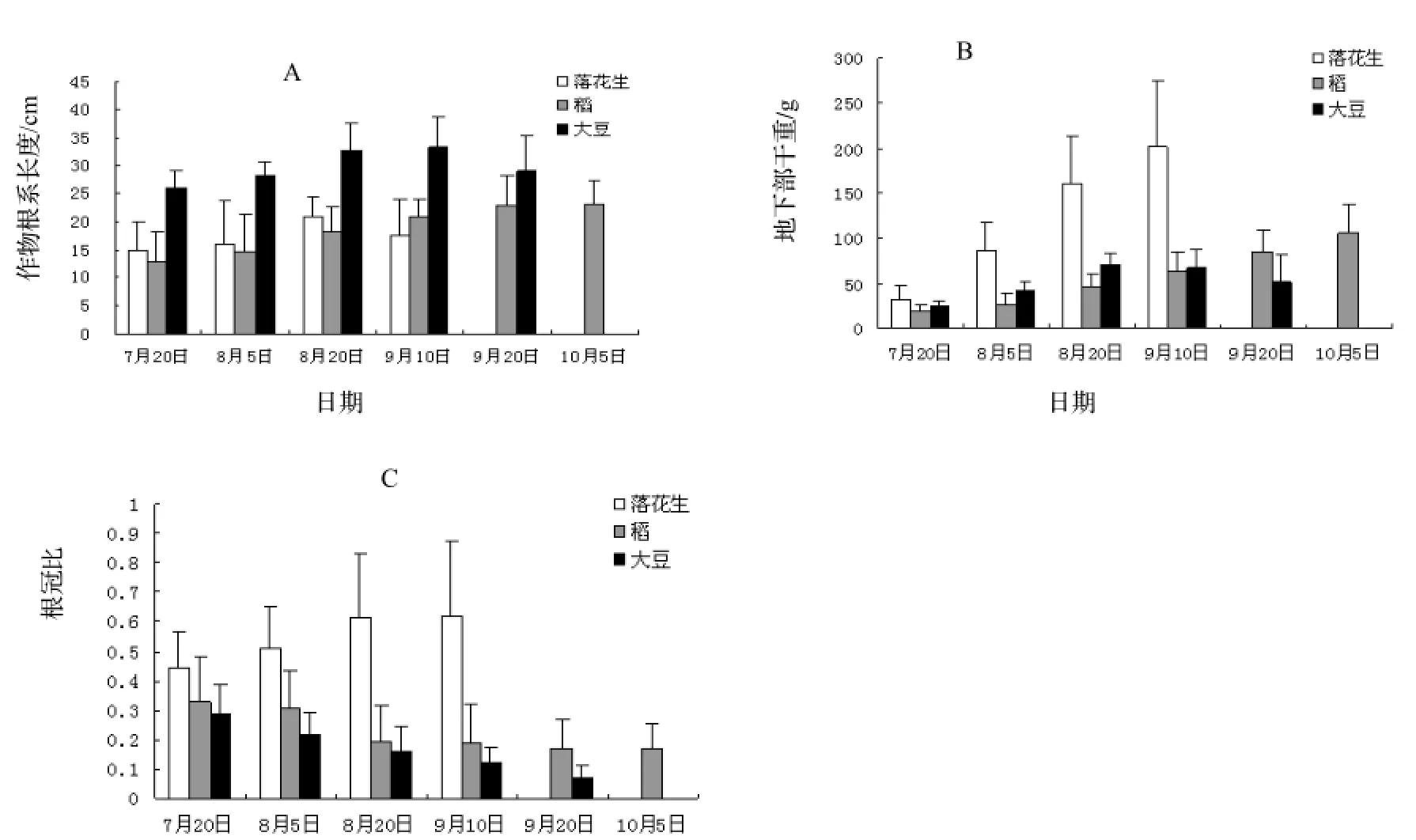
图2 3种农作物根系长度、生物量及根冠比变化规律Figure 2 Variation of root length, root biomass and root shoot ratio of the tested crops
由图2A可知油茶幼林复合系统内大豆在整个生长周期内根系长度显著高于稻、落花生,最长达33.33 cm。稻、落花生整个生长周期内根系基本分布于油茶幼林复合系统10 ~ 20 cm的耕作层。至采收时,落花生根系最短,为17.67 cm。稻根系生长呈现稳步增长趋势。而两种豆科作物落花生、大豆在生长期趋势一致,呈先增加后下降趋势,根长峰值均出现在采收前,在生长末期根系长度有一定下降,这可能是豆科作物成熟后根系腐烂所致。
图2B反映了稻、落花生地下部干重随生长期呈递增趋势,而大豆则呈先增加后下降的趋势。落花生、稻、大豆地下部干重较7月20日比,分别增长了527%,461%,110%。其中落花生因荚果处于地下因而地下部干重最大,达200.7 g,分别为稻、大豆的1.9倍、3.9倍。由图2C可知作物生长期内根冠比:落花生>稻>大豆,落花生因荚果处于地下因而根冠比最大,稻、大豆根冠比最大值出现在7月20日,分别为0.33,0.28,随着时间的推移,稻、大豆根冠比逐渐减小,至采收时稻、大豆根冠比分别降至为0.16、0.05。前期稻、大豆根冠比较大,说明幼苗根系生长速度高于地上部分,而生长后期根冠比逐渐见效,地上部生长速度高于根系,落花生则不同,其根冠比随着时间呈递增趋势。分析作物根冠比的生长变化规律,说明作物幼苗期需有充足的养分供根系生长。
由图3A可以看出,落花生、稻、大豆单株根系总长均随时间变化呈先增加后下降的趋势。三种作物收获时,落花生、稻、大豆根系总长与7月20日时相比,分别增长了120%,174%,107%,其中,大豆根系总长最长,为875.04 cm,落花生最短,仅为197.25 cm。整个生长周期内,根系总长度:大豆>落花生>稻,说明大豆较其他两种作物有着更发达的水分、养分吸收能力。作物籽粒成熟后,3种作物根系总长度均有一定程度减少,表明已有一部分根系腐烂还田。
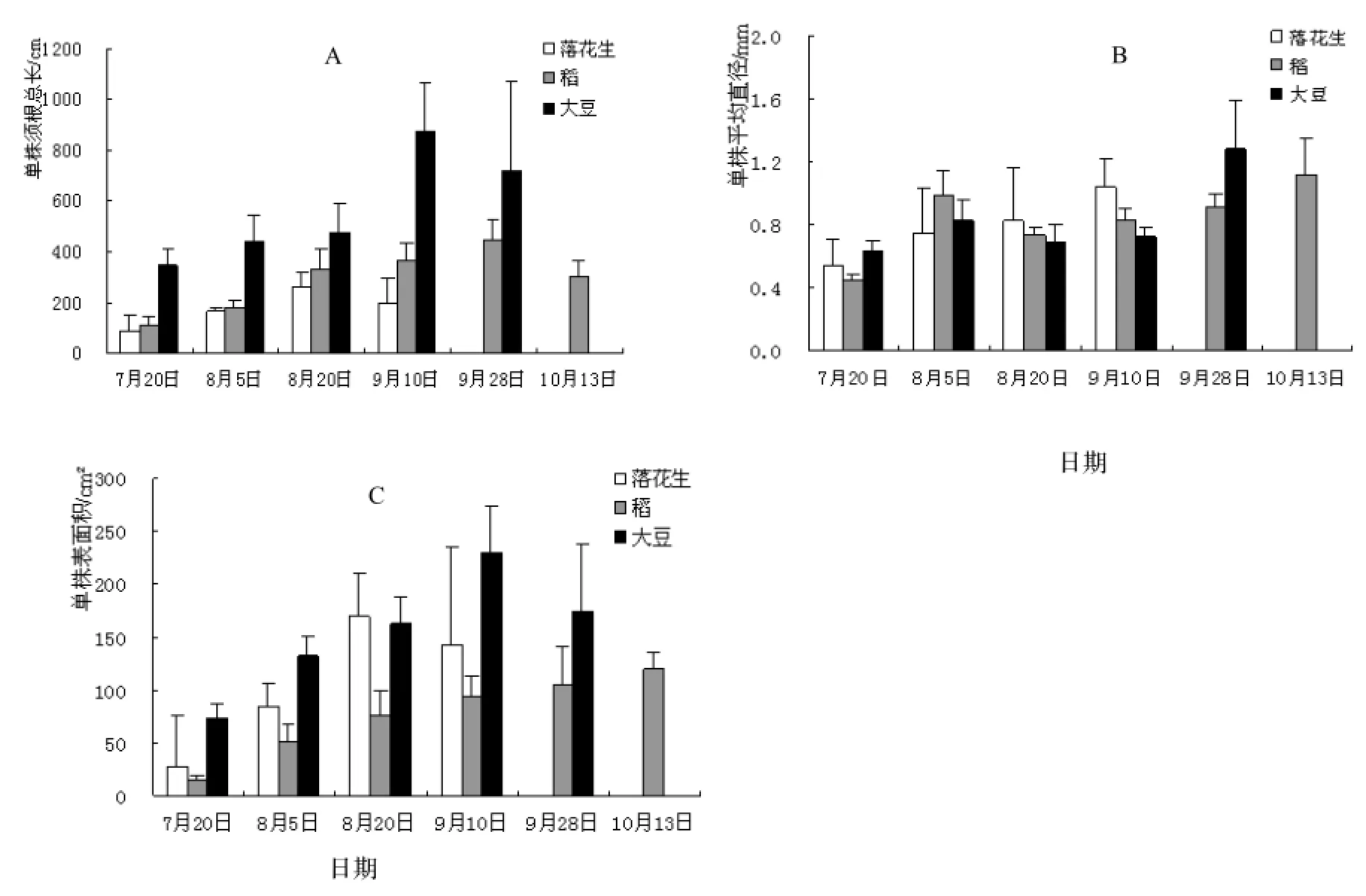
图3 3种作物根系总长、平均直径、表面积生长变化规律Figure 3 Variation of total root length, average diameter and superficial area of the tested crops
图3 B反映了3种农作物根系的平均直径均随着时间呈递增趋势,三种作物采收时,落花生、稻、大豆平均直径由7月20日0.53 mm,0.44 mm,0.63 mm增长至采收时1.04 mm,1.12 mm,1.28 mm,分别增长了96%,155%,103%。落花生、大豆在其生长后期细根腐烂还田,因而在收获期间平均直径略有增加。
由图3C可以看出,稻根系表面积随着时间一直稳步增长,落花生、大豆则在生长后期呈下降趋势。落花生、稻、大豆根系表面积采收时分别较7月20日增长了421%,704%,139%,稻根表面积增幅最大。作物收获时大豆根表面积最大,为174.34 cm2,稻根表面积最小,为119.78 cm2。整个生长周期内,根系表面积:大豆>落花生>稻。根系表面积被认为是衡量根系对水分和养分吸收能力的参数之一,根表面积的多少直接影响到根系吸收面积。落花生、大豆根表面积峰值均出现在采收之前的成熟期,表明落花生、大豆根表面积随着籽粒的成熟而增加,在籽粒完全成熟后表面积达到最大,对水分和养分的吸收也达到最大化。
2.3 农作物生长发育速度比较
以RGR和CGR为指标对油茶幼林复合系统三种农作物不同阶段的生长发育情况进行了比较。结果表明,三种农作物不同阶段生长有差别,同一作物的 RGR和CGR随生长期变化规律基本一致。落花生的RGR和CGR最大值均出现在7月20日至8月5日,且呈下降趋势。稻、大豆生长速率易受水热条件影响,其RGR和CGR呈升-降-升-降趋势。大豆的RGR和CGR最小值均出现在8月20日至9月10日,表明在此期间大豆植株生长较为缓慢。
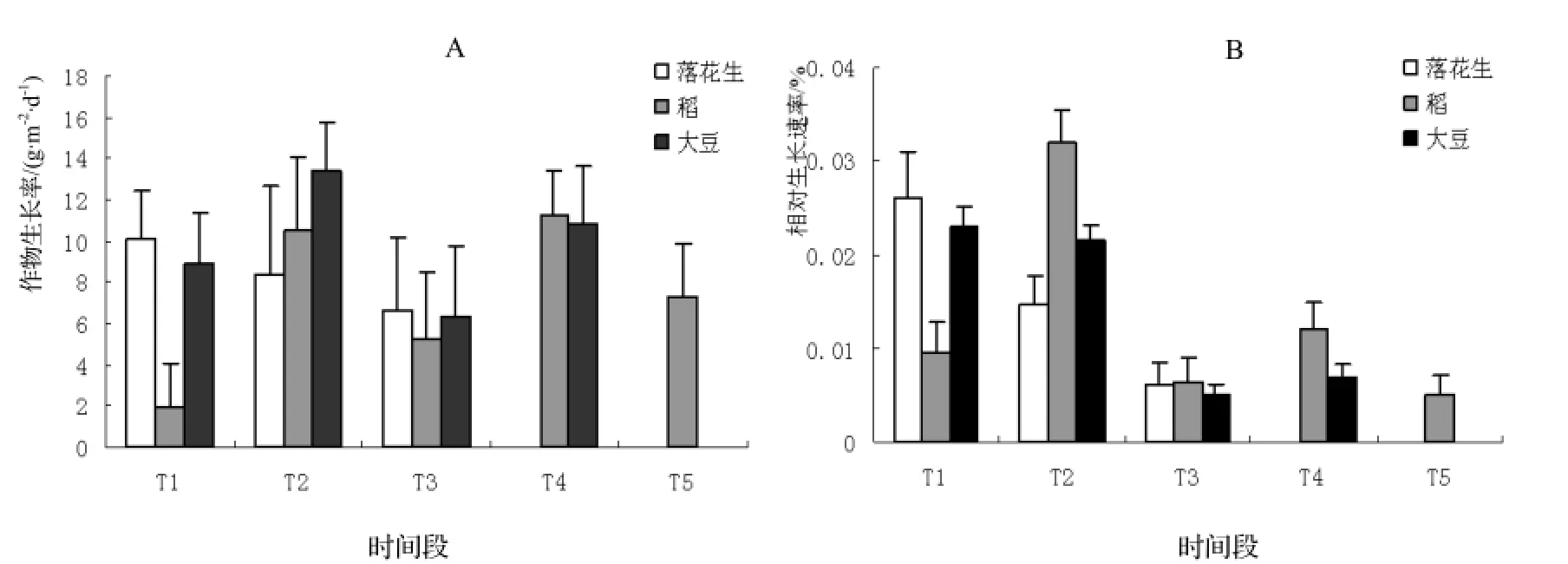
图4 3种作物生长率、相对生长速率变化Figure 4 Variation of GCR and RGR of the tested crops
2.4 农作物生物量比较
油茶幼林复合模式3种不同作物的生物量(以作物最终收获时的生物量作为研究对象)及其在根、茎叶、籽粒的分配情况如表1。不同结构的油茶幼林复合模式,由于其系统组成及所处微环境不同,从而影响群落中作物的生长,表现出生物量及其分配规律的差异。各油茶幼林复合模式中除籽粒(占总生物量的17.3% ~ 28%)作为完全的收获物外,其茎、叶、根均可以以各种方式还田,生物量归还率达到71.0%~82.7%。尤其是落花生、大豆2种绿肥作物,其根部根瘤菌固氮对增加土壤肥力以及维持油茶幼林复合系统养分平衡及生产稳定性十分有利。

表1 3种作物生物量及生产力在器官中的分配Table 1 The biomass and productivity’s distribution in organs of the three types of crops
油茶幼林3种复合作物收获时生物量累积见表1。模式内落花生、稻、大豆的累积生物量分别达到523g•m-2,706.5 g•m-2,769 g•m-2。油茶-稻和油茶-大豆模式生物量明显高于油茶-落花生复合模式,稻、大豆分别为花生生物量的1.4,1.5倍,表明油茶-稻、油茶-大豆复合模式较油茶-落花生模式相比更有利于作物生物量的积累。
对油茶幼林复合经营不同模式及根、茎、叶、籽粒等各组分生物量的研究表明,各模式中农作物的年生物量不同,作物年生物量排序:大豆>稻>落花生。大豆的生物量最高,达到769 g• m-2,落花生年生物量最低,为523 g•m-2。油茶幼林复合系统中农作物生物量在各器官的分配基本相似,以茎、叶生物量最高,约占作物总生物量61.6%~68%;以根的生物量最低,占6.6%~14.7%。
3 结论与讨论
(1)油茶幼林复合系统作物植株高度由高到低分别为稻>大豆>落花生,根系长度大豆最长,油茶新梢长度为油茶-落花生(19.2 cm)>油茶-大豆(18.2 cm)>油茶-稻(18 cm)>油茶纯林(15.2 cm)。试验结果表明,油茶作物复合经营模式中,油茶新梢生长均高于未套作模式。不同复合经营模式下油茶新梢生长的差异,与套种作物的根系长度和植株生长高度有关。落花生、稻根系分布在0~20 cm的浅耕作层,与油茶根系竞争不明显,另外由于稻植株较高,可能对油茶新梢生长发育造成影响。
(2)对油茶幼林复合系统内大豆单株根系总长、根表面积研究表明,较落花生、稻相比,其根长、根表面积以及根系扎入土壤的深度都更有利于复合系统中潜在的根系竞争。豆科作物根瘤菌固氮后腐烂还田也有助于提高耕作层的耕性,提高土壤肥力,加速油茶幼林复合系统土壤熟化。
(3)油茶幼林复合系统内作物生物量在器官中的累积都以茎、叶最大,以根为最小。生物量的多少反映了油茶幼林复合系统内复合强度的大小,大豆具有最高的生物量和还田率,因此油茶-大豆这种复合经营模式有助于加强作物秸杆循环利用,维持油茶幼林系统的稳定性、可持续性。
[1] 胡芳名,谭晓风,刘惠民,等. 中国主要经济树种栽培与利用[M]. 北京,中国林业出版社,2006.
[2] 庄瑞林.中国油茶(第二版)[M]. 北京:中国林业出版社,2000.
[3] 明延柏,李爱华,袁知雄,等. 油茶幼林不同套种模式与综合效益分析[J]. 技术开发,2012,04:98-101.
[4] 冯金玲,郑新娟,等. 5种栽培模式对油茶土壤微生物及酶活性的影响[J]. 西南林业大学学报,2016,36(2):10-16.
[5] 王玉娟,陈永忠,等. 覆草间种对油茶林土壤养分及生长量影响的主成分分析[J]. 中南林业科技大学学报,2010,30(6):43-49.
[6] 文亚雄,等. 不同间作模式对油茶幼林病虫害发生的影响[J]. 经济林研究,2016.34(1):129-134.
[7] 陈隆生,陈永忠,等. 不同间作模式对油茶幼林生长的影响[J]. 湖南林业科技,2012,37(1):15-16.
[8] Richardson, D.M. 1998. Forestry trees as invasive aliens. Conservation Biology 12:18-26.
[9] 王玉娟,陈永忠,等. 稻草覆盖对油茶幼林林地土壤温度及新梢的影响[J]. 经济林研究,2009,27(2):49-52.
[10] 腾维超,等. 油茶大豆间作对盆栽土壤化学和生物性质的影响[J]. 中南林业科技大学学报,2013,33(2):24-27.
[11] 刘丁林,张杨珠,聂军,等. 不同绿肥作物与油茶幼林套种对丘岗红壤pH值变化和有效养分含量的影响[J]. 湖南农业科学,2013,09:43-45,49.
[12] 钱萍. 套种对油茶幼林的土壤理化性状及其生长的影响[D]. 南昌:江西农业大学,2011.
[13] 高伟. 浙西南不同间作模式对油茶生境及光合作用的影响[D]. 长沙:中南林业科技大学,2014.
[14] 陈永忠,王玉娟,王湘南,等. 间种对油茶林地土壤理化性质及幼林生长量的影响[J]. 南京林业大学学报(自然科学版),2011,35(5):117-120.
[15] 王轶虹,王美艳,等. 2010年中国农作物净初级生产力及其空间分布格局[J]. 生态学报,2016,36(19):1-11.
[16] 腾维超. 油茶-农作物间作系统生理生态及经济效益评价[D]. 南京:南京林业大学,2013.
[17] 程鹏. 银杏复合经营生物生存力及生态效应研究[D]. 南京:南京林业大学,2010.
Growth Rhythm of Three Crops Interplanted in Young Camellia oleifera Stand
CHEN Zhi-wei1,2,LONG Meng-qi1,HE Xiao-yong2,GE Yong-jin2,ZHANG Ri-qing1*
(1. Central South University of Forestry and Technology, Changsha 410002, China; 2. Lishui Forestry Institute of Zhejiang, Lishui 323000, China)
∶ 1-year Camellia oleifera seedlings were planted at Lanju, Longquan of Zhejiang province in March of 2012, and interplanting of Arachis hypogaea, Glycine max and was carried out in May of 2015. Biomass of interplanted crops was determined from July to harvest in the current year. Results showed that O. sativa had the most height growth, topped to 150.23cm at the date of harvesting. G. max had the highest biomass accumulation, up to 769g, and had the highest root length of single plant, total fine root length and root surface area, with 33.33cm, 875.04cm and 174.34cm2respectively. The experiment demonstrated that G. max had the best root system for interplanting at C. oleifera stand. .
young plantation of Camellia oleifera; agroforestry system; growth rhythm; biomass
S753.53+6
A
1001-3776(2016)06-0052-06
2016-02-28;
2016-09-30
油茶新造幼林地生态修复关键技术研究(2014NZH07)与油茶基地建设及高效栽培技术示范推广(2012JDJS04)联合资助作者简介:陈志伟(1992-),男,湖南长沙人,硕士研究生,研究方向为经济林;*通讯作者。
猜你喜欢
南方农业·下旬(2022年4期)2022-05-24
金秋(2021年20期)2021-02-16
故事作文·低年级(2020年7期)2020-07-28
作文新天地(高中版)(2019年6期)2019-08-15
——许地山、老舍同题散文《落花生》比较赏析
作文新天地(2019年16期)2019-01-11
中国林业产业(2019年11期)2019-01-09
现代园艺(2018年2期)2018-03-15
海峡姐妹(2017年10期)2017-12-19
中国西部(2017年4期)2017-04-26
源流(2017年1期)2017-03-07
