
Restudy of the Late Oligocene dormice from northern Junggar Basin
2016-03-29 03:07:19WUWenYuMENGJin2YEJieNIXiJunBIShunDong
古脊椎动物学报(中英文) 2016年1期
WU Wen-YuMENG Jin,2YE JieNI Xi-JunBI Shun-Dong,3
(1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044, China wuwenyu@ivpp.ac.cn)
(2Division of Paleontology,American Museum of Natural HistoryNew York NY 10024, USA)
(3Department of Biology,Indiana University of PennsylvaniaIndiana PA 15705, USA)
Restudy of the Late Oligocene dormice from northern Junggar Basin
WU Wen-Yu1MENG Jin1,2YE Jie1NI Xi-Jun1BI Shun-Dong1,3
(1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044, China wuwenyu@ivpp.ac.cn)
(2Division of Paleontology,American Museum of Natural HistoryNew York NY 10024, USA)
(3Department of Biology,Indiana University of PennsylvaniaIndiana PA 15705, USA)
A new glirid genus and species,Gliruloides zhoui, is named based on specimens from the Late Oligocene Tieersihabahe Mammal Assemblage Zone I (Tie-I zone) of the northern Junggar Basin, Xinjiang. The new genus is diagnosed by the following features: middle-sized dormouse; occlusal surface of cheek teeth concave; upper and lower cheek teeth dominantly with nine transverse ridges; the anterotrope(id) and posterotrope(id) well developed and extending almost full length of corresponding valleys; transverse ridges of upper cheek teeth usually free-ended labially; M1/2 with V- or narrow U-shaped trigon; the precentroloph not connected to the endoloph that is incomplete or nearly complete; the endolophid in lower cheek teeth discontinuous or continuous; labial end of the anterolophid curving slightly distally but not connected with the protoconid; root number of p4, m1-3, P4 and M1-3 being 2, 2, 3 and 3 respectively. We discuss the differences ofGliruloidesfromGlirulusandVasseuromysand assign the AnatolianVasseuromys duplexandVasseuromysaff.V. duplexfrom the Early Miocene of Turkey toGliruloides. It is posited thatGliruloidesandGlirulusmay share a common ancestor similar toGlisguerbuezifrom the Lower Oligocene of Thrace, Turkey.Gliruloidesmight live in a relative wet and warm biotope.
northern Junggar Basin, Late Oligocene, Tieersihabahe Mammal Assemblage Zone I, dormouse
Our study on earliest Chinese dormice (Wu et al., 2000) was based on specimens collected in 1998 from the Late Oligocene Tieersihabahe Formation in northern Junggar Basin of Xinjiang. Only four teeth, among which three were assigned toGlirulussp., were available at the time. Additional 11 teeth were collected by screenwashing at the same level of thesame locality (XJ 98024) and other nearby localities (XJ 98035, XJ 200209 and XJ 20004) in four fi eld seasons from 1999 to 2002. All teeth but one right M3 (IVPP V 18113) are almost identical to the specimens described by Wu et al. (2000). Undoubtedly, they belong to the same taxon. Since the work of Wu et al. (2000), we have realized that these teeth show high similarities toVasseuromysduplexfrom the Early Miocene of Anatolia (Ünay, 1994), and therefore citedVasseuromyssp., instead ofGlirulussp., in fauna lists of our successive papers (Meng et al., 2001, 2006; Ye et al., 2001a,b; 2003a,b) but without further explanation. The present paper restudies the Late Oligocene Junggar dormouse based on all material available to us and reinterprets its taxonomic position.
Our study reveals that the specimens of the Late Oligocene Junggar dormouse are more similar toVasseuromysduplexthan to any other dormouse and thatV. duplexdiffers considerably from other EuropeanVasseuromysspecies andGlirulus. Thus, we name a new genus,Gliruloides, for the Junggar dormouse and assign the Anatolian species,Vasseuromys duplexandVasseuromysaff.V. duplex, to this genus.
In describing the tooth morphology, we use a modified terminology derived from de Bruijn (1966) and Freudenthal (2004), which is convenient and unambiguous for description of the extra ridges of the dormice cheek teeth (Fig. 1). The SEM photographs were taken by Meng Jin, using the Hitach S-4700 scanning electron microscope at the American Museum of Natural History in 2005.

Fig. 1 Dental terminology of Gliridae Modi fi ed from de Bruijn (1966) and Freudenthal (2004)
Family Gliridae Thomas, 1897
Subfamily Dryomyinae de Bruijn, 1967
GenusGliruloidesgen. nov.
Etymologyglirulus+ oides,indicates that the dormouse is morphologically similar to genusGlirulusbut should not beGlirulus.
Type speciesGliruloides zhouigen. et sp. nov.
Diagnosis Medium-sized dormouse with concave occlusal surface of the cheek teeth. The upper and lower cheek teeth possess dominantly nine transversal ridges. The anterotrope(id) and posterotrope(id) of the upper and lower cheek teeth are well developed and extend almost full length of the corresponding positioned valleys. The trigon of M1-2 is V-or narrow U-shaped, and the endoloph of upper cheek teeth is incomplete or nearly complete. The main ridges of the upper cheek teeth are labially free-ended. The endolophid of the lower cheek teeth is discontinous or continuous. The labial end of the anterolophid of the lower molars slightly curves distally, but not connected to the protoconid. Lower cheek teeth are two-rooted.
Differentiate diagnoses 1)Gliruloidesdiffers fromGlirulusin having a V-shaped or narrow U-shaped trigon on M1-2, the endoloph incomplete or nearly complete on upper cheek teeth, the lingual end of the precentroloph not connected to the endoloph, and main ridges in the upper cheek teeth free-ended labially. In contrast, the upper molars ofGlirulushave wide U-shaped trigon, complete endoloph, and the precentroloph that connects lingually to the endoloph (except for geologically older representatives in which the endoloph may be incomplete and the precentroloph unconnected to the endoloph, refer Ünay, 1994). InGlirulusthe main tooth ridges usually connect with each other labially and form the anterior and posterior loops (see Fig. 4D).
2)Gliruloidesdiffers fromVasseuromysmainly in having nine ridges on most upper and lower molars, in having the developed anterotrope and posterotrope on the upper cheek teeth, and the usually regular and continuous extra ridges. InVasseuromys, however, the anterotrope and posterotrope outside the trigon of the upper cheek teeth are absent or weak, and the extra ridges of the cheek teeth are usually irregular and interrupted (see Fig. 4B).
Included speciesGliruloides duplex(Ünay, 1994) from Early Miocene (MN2) of Anatolia, localities Harami 1-3, Turkey.Gliruloidesaff.G. duplex(Ünay, 1994) from Early Miocene (MN1) of Anatolia, localities Kilçak 0, Kilçak 0" and Kilçak 3A-3B.
Gliruloides zhouigen. et sp. nov.
Glirulussp., Wu et al., 2000
Vasseuromyssp., Ye et al., 2001a, b, 2003a, b; Meng et al., 2001, 2006
Etymology In honor of the late Dr. Zhou Mingzhen (Minchen Chow), an academician of the Chinese Academy of Sciences and the pioneer paleomammalogist in China.
Holotype Left M2, IVPP V 18110.1 (Fig. 2C).
Paratype One left M3 (IVPP V 18110.2), one right m1 and one left m1 (V 18110.3-4), and one right m3 (V 18110.5) (Fig. 2D, H, J, K).
Type locality and horizon Locality XJ 98035, Tieersihabahe of northern Junggar Basin, Xinjiang; Tieersihabahe Mammal Assemblage Zone I; Late Oligocene.
Referred specimens One left P4 (IVPP V 18111.1, Fig. 2A), two right p4 (V 18111.2-3, Fig. 2F-G), one right m1 (V 18111.4, Fig. 2I) from XJ 98024. One left M1/2 (V 18112, Fig.2B) from XJ 200209.
The specimens described in 2000 include one right M2 (IVPP V 11812.1), one left M3 (V 11812.2) and one left m1 (V 11812.3) from XJ 98024 (Wu et al. 2000: pl. 1, fi gs. 1-3, 5).
All specimens listed above were from Tieersihabahe of northern Junggar Basin;Tieersihabahe Mammal Assemblage Zone I; Late Oligocene.
One right M3 (V 18113, Fig. 2E), collected from XJ 20004 at Saerduoyila of northern Junggar Basin. The stratum producing the specimen is correlative to that of Tieersihabahe.
Diagnoses The p4 possesses eight transverse ridges. The anterotropid on the lower cheek teeth is usually single. The endoloph on upper cheek teeth is nearly complete, whereas the endolophid of lower cheek teeth is interrupted. The protoconid and mesoconid of the lower molars are hook-like. The metalophid extends slightly distally to the labial side and then turns abruptly mesiolabially.
Differentiate diagnosesGliruloides zhouidiffers from the Anatolian speciesG. duplexandGliruloidesaff.G. duplexin having 1) nearly complete endoloph on M1/2, 2) hook-like protoconid and mesoconid of the lower molars, 3) the metalophid almost always extends slightly distolabially and then turns abruptly mesiolabially and 4) possibly the rare presence of doubled-anterotropid on the lower molars. In contrast,G. duplexandGliruloidesaff.G. duplexnormally have incomplete endoloph on M1/2 and rarely have hook-like protoconid and mesoconid. In both forms, the metalophid usually extends mesiolabially and a doubledanterotropid is common on the lower molars.
Measurements (length × width in mm) P4 (V 18111.1) 0.87 × 1.05; M2 (V 18110.1) 1.05 × 1.25; M1/2 (V 18112) 1.03 × 1.09; M3 (V 18110.2) 0.92 × 1.05; M3 (V 18113) 0.77 × 0.98; p4 (V 18111.2) 0.86 × 0.68; p4 (V 18111.3) 0.84 × 0.67; m1 (V 18110.3) 1.17 × 1.05; m1 (V 18111.4) 1.16 × 1.09; m1 (V 18110.4) 1.13 × 1.06; m3 (V 18110.5) 1.11 × 1.00 (Except P4 is larger and m3 is slightly longer than inG. duplex, the other teeth are approximate to those ofG. duplexin size.
Description The occlusal surface of all cheek teeth is concave. All the upper cheek teeth have usually nine transverse ridges, including the six main ridges (anteroloph, protoloph, metaloph, posteroloph, precentroloph and postcentroloph) and three extra ones (anterotrope, prototrope and posterotrope). Both the anterotrope and posterotrope extend almost the full length of the valleys where they locate. The labial ends of the main ridges are free or lightly connected. The lingual wall of the endoloph on upper cheek teeth is well decorated. All upper cheek teeth have three roots (one major lingual and two minor labial ones).
P4 The left P4 (V 18111.1; Fig. 2A) is suboval-shaped in occlusal view. Its occlusal surface is strongly concave. The anteroloph has its labial end separated from the protoloph and the lingual end touched the protoloph but not the endoloph. The lingual end of the posteroloph is weakly in contact with the endoloph and labially connects to the metaloph. The protoloph is labially free and lingually connects to the endoloph; it is interrupted midway by the U-shaped anterotrope-prototrope connection. The anterotrope extends to the labial border and stays free, whereas the prototrope is half long the anterotrope. The precentroloph ends free labially, but lingually it bifurcates to join the protoloph and metaloph, respectively. The postcentroloph is short and situated in the middle of the valley, with both ends being free. The metaloph is convex distally and interrupted near its lingual end. The posterotrope is long, fi lling nearly thewhole length of the corresponding valley. The trigon is V-shaped.
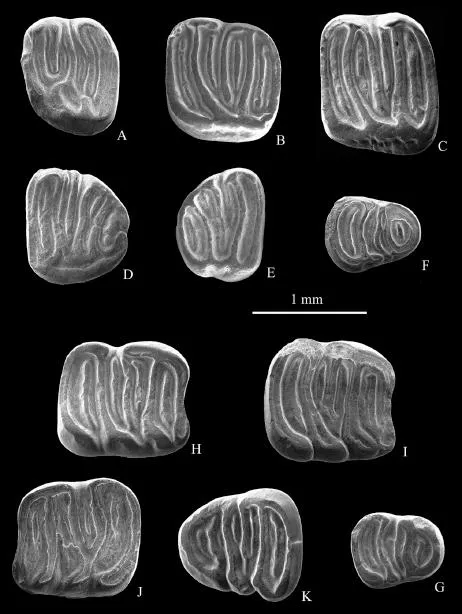
Fig. 2Gliruloides zhouigen. et sp. nov. A. P4 sin. IVPP V 18111.1; B. M1/2 sin. V 18112; C. M2 sin. V 18110.1, holotype; D. M3 sin. V 18110.2, paratype; E. M3 dex V 18113; F. p4 dex, V 18111.2; G. p4 dex, V 18111.3; H. m1 dex, V 18110.3, paratype; I. m1 dex, V 18111.4; J. m1 sin., V 18110.4, paratype; K. m3 dex., V 18110.5, paratype
M1/2 Two M2 and one M1/2 are in the collection. The identi fi cation of specimen V 18112 (Fig. 2B) as M1 or M2 cannot be certain so that we denote it as M1/2 (but is probably M1 because of its V-shaped trigon). The holotype M2 (V 18110.1) is moderately worn and almost identical to the specimen V 11812.1 reported by Wu et al. (2000: pl. 1, fi g. 1). The occlusal surface is wider than long. The paracone and metacone are prominent. The anteroloph is labially weakly connects to the paracone with a shallow furrow in between and lingually joins the endoloph. The paracone connects the labial end of the precentroloph loosely. The metacone is separated anteriorly from the postcentroloph and posteriorly fromthe posteroloph by a narrow and very shallow furrow, respectively. The precentroloph is long, extending to the point near the endoloph. The trigon is narrowly U-shaped and contains the long prototrope, precentroloph and postcentroloph. The lingual wall of the endoloph is well decorated. Specimen V 18112, a left M1/2 is slightly worn and closely similar to the holotype, but differs from the latter in being wider anteriorly than posteriorly, the V-shaped trigon, precentroloph being labially far separated from the paracone and curving backwards to the metacone, the anteroloph being labially free from the paracone, and loosely contacting with the protocone lingually to form a nearly complete endoloph. The anterotrope and posterotrope on all three specimens extend the whole length of the corresponding occupied valley.
M3 There are three M3 specimens (V 11812.2, V 18110.2 and V 18113), which are characterized by a trapezoid-shape, being much wider anteriorly than posteriorly. M3 has usually the same number of ridges as M1/2 but the precentroloph is much shorter than the postcentroloph. Specimen V 18110.2 is well worn. The labial side of the paracone was slightly damaged but it seems in connection with the anteroloph and protoloph. The precentroloph is much shorter than the postcentroloph and extends labially to the labial margin of the tooth and has no contact with both the paracone and metacone. The postcentroloph extends lingually and almost reaches the endoloph; its labial end is weakly connected with the metacone. The prototrope is absent, instead is a metatrope present inside the trigon. The metatrope is short, only a half width of the valley, and locates labially. Both metaloph and posteroloph are interrupted by a narrow gap at the midway. It seems that the endoloph is separated from the lingual end of the anteroloph before the tooth was worn. The specimen V 11812.2 (Wu et al., 2000: pl. 1, fi g. 2) is highly similar to V 18110.2. There is, in addition to the anterotrope, an extra ridge between the anterotrope and protoloph. Both prototrope and metatrope are present. The specimen V 18113 (Fig. 2E) from XJ 20004 is similar to specimen V 11812.2, but its ridges are more slender and the anterotrope is weaker and shorter. We tentatively assign this tooth to this species.
The p4 Both p4 are round trapezoid in shape, wider posteriorly than anteriorly (Fig. 2F, G). The anterolophid is convex mesially and connected distally to the metalophid to form a closed loop. The centrolophid is thin and long, connected lingually to the metaconid weakly, and extends to the labial border. The labial end of the mesolophid turns abruptly mesially along the labial border on one specimen (V 18111.2) but becomes well-swollen labially on the other (V 18111.3); the mesolophid connects lingually to the posterolophid at the entoconid, and labially connected to or separated from the posterolophid. The anterotropid and posterotropid are well developed, and the mesotropid is short or long. The metatropid is absent in both teeth so that p4 is eight-ridged.
The m1 Four specimens are in the collection, including the one described by Wu et al. (2000: pl. 1, fi g. 3). They are quite monotonous morphologically and generally trapezoidshaped with the anterior end being slightly narrower than the posterior end. The m1 usually has nine transverse ridges, including fi ve main ridges (anterolophid, metalophid, centrolophid,mesolophid and posterolophid) and four extra ridges (anterotropid, metatropid, mesotropid and posterotropid). All four extra ridges are well developed, of which the anterotropid and posterotropid extend nearly the entire length of the located valleys. However, specimen V 11812.3, described by Wu et al. in 2000, remains an exception because it has an additional secondary extra ridge between the anterotropid and metalophid. For all specimens of m1, the anterolophid is slightly concave mesially, labial end slightly curved distally and free-ended; its lingual end slightly curved distally too, and gradually merges into the metaconid lingually. The metalophid is lingually connected to the metaconid and extends slightly distolabially, with its labial end turning abruptly mesially to form a hook-like protoconid that is not connected to the anterolophid. The centrolophid is long and extends labially near the mesoconid but does not reach to the labial border of the tooth. Lingually the centrolophid is either separated from or connected with the metaconid. The mesolophid is slightly convex distally and extends mesiolabially where, like the metalophid, it turns abruptly mesially and forms a hook-like mesoconid that is separated from the protoconid mesially and from the hypoconid distally. The posterolophid joins the mesolophid lingually at the entoconid. An endolophid is absent.
The m3 This tooth is mesially much wider than distally. The anterolophid is slightly convex mesially. Except for the four extra ridges seen in m1, there are two more secondary extra ridges: one mesial and the other distal to the posterotropid, respectively. Furthermore, a very small and weak enamel bulge but not ridge is present between the anterolophid and anterotropid.
All lower cheek teeth have two roots, but the mesial root of m1 (V 11812.3) shows a trend of bifurcation at the end.
Several teeth display distinctive striations of abrasion in mesiolingual-distolabial direction on the occlusal surface (Fig. 3), which we record here. We think this information should be useful for further study of this animal in masticatory movement, dietary, and perhaps taxonomy.

Fig. 3 Occlusal view of the lower and upper cheek teeth ofGliruloides zhouiArrows indicate the direction of striations caused by wear on the concave crown surface
Comparisons and discussions As mentioned above, the Junggar glirid, represented by three specimens at the early time, was fi rst assigned toGlirulus(Wu et al., 2000) because it is similar toGlirulusin having usually nine ridges on both upper and lower cheek teeth, withwell-developed anterotrope and posterotrope in upper cheek teeth and the anterotropid and posterotropid in lower cheek teeth. However, additional specimens collected subsequently made us to reconsider the taxonomic assignment of the Junggar glirid. We have listed above several features that differ the Junggar form fromGlirulusin the differentiate diagnoses. At the same time, we noted that the Junggar form is quite similar to the Turkish Lower Miocene dormouse,Vasseuromys duplex, in both upper and lower molars. Thus, we considered that the Junggar glirid should not be assigned toGlirulusand have referred it toVasseuromysin several of our papers (Meng et al., 2001, 2006; Ye et al., 2001a, b; 2003a, b) without explaining the reason until this study.
Vasseuromyswas established by Baudelot and de Bonis (1966) based on the type speciesV. rugosusfrom the Lower Miocene of Laugnac, France. The only material of this species was a mandible with p4-m2. The original generic diagnosis states (translated from French by Daams and de Bruijn, 1995:50): “Medium-sized Gliridae. Cheek teeth with concave occlusal surface. Lower molars characterized by a centrolophid reaching the labial border and by a longitudinal prolongation of the labial cusps that form a nearly continuous ectolophid”. Based on additional specimens of upper cheek teeth from the type locality de Bonis (1973:54) emended the diagnosis of the genus as: “Vasseuromyscharacterized by multiplicity of the extra ridges and by the upper molars with a continuous endoloph (translated from French)”. The upper cheek teeth referred in the work of de Bonis (1973) consisted of one P4, one M1/2, and two M3. While working on the dormice from Austria and Spain, Daxner-Höck and de Bruijn (1981) and Alvarez Sierra et al. (1990) have independently visited the collections ofVasseuromys rugosushoused at the University of Utrecht; all the specimens are from the type locality. The two research teams noticed that there were several upper molars ofV. rugosuswith incomplete endoloph. Alvarez Sierra et al. (1990) wrote: “Although de Bonis (1973) characterizes this species as having a continuous endoloph (on the basis of one worn specimen), several M1,2 without endoloph are present in the collections of the University of Utrecht. The specimens from the Utrecht collections have a long anteroloph whose lingual end descends toward the base of the protocone.” Daxner-Höck and de Bruijn (1981) further provided fi gures of the upper and lower molars ofV. rugosus, which show the V-shaped or narrow U-shaped trigon and incomplete endoloph in the upper molars (Daxner-Höck and de Bruijn, 1981: fi g. 1-h,r,s,t,u; fi g. 2-m,n,o,p) (see Fig. 4B).
Up to now ten species have been included in this genus (ref. Daams and de Bruijn, 1995; Ruiz-Sánchez et al., 2012a, b; 2014):
Vasseuromys rugosusBaudelot & de Bonis, 1966; type locality and type level: Laugnac of France, MN2B, Early Miocene.
V. priscusde Bonis, 1973; type locality and type level: Moissac 1 of France, MN1, Early Miocene.
V. pannonicus(Kretzoi, 1978); type locality and type level: Budapest, Freshwater Limestone of the Széchenyi hill; MN10?, Late Miocene (Synonym:V. theniiDaxner-Höck &de Bruijn, 1981).
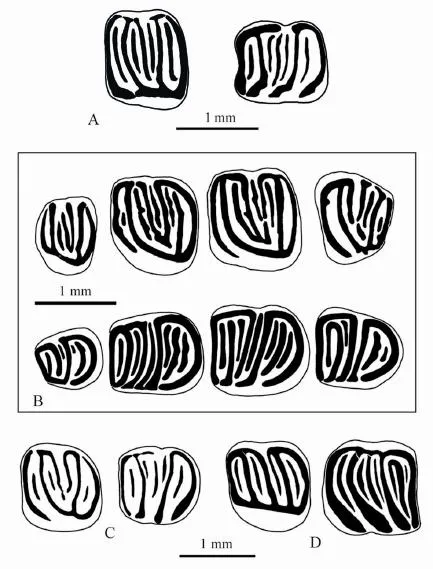
Fig. 4 The comparison of the upper and lower cheek teeth ofGliruloides zhoui,Vasseuromys rugosus,Glis guerbuezi, andGlirulus japonicusA.Gliruloides zhouigen. et sp. nov. from northern Junggar Basin, Late Oligocene (M2 and m2); B.Vasseuromys rugosusBaudelot & de Bonis, 1966 from the type locality Laugnac of France, MN2B, Early Miocene (after Daxner-Höck and de Bruijn, 1981) (P4-M3 and p4-m3); C.Glis guerbueziÜnay-Bayraktar, 1989 from Thrace of Turkey (modi fi ed from Ünay-Bayraktar, 1989) (M2 and m2); D.Glirulus japonicus(Schinz, 1845) from the upper part of Horizon M of Kannondo Cave Site of Japan (modi fi ed from Kawamura, 1989) (M1 and m2)
V. autolensis(Cuenca, 1985); type locality and type level: Autol, La Rioja, Spain; MN1, Early Miocene.
V. bacchius(Martínez-Salanova, 1987); type locality and type level: Fuenmayor 2, Autol 1, La Rioja, Spain; MN2B, Early Miocene.
V. elegansWu, 1993; type locality and type level: Stubersheim 3, Germany, MN3, Early Miocene.
V. duplexÜnay, 1994; type locality and type level: Harami 1, Turkey; MN2, Early Miocene.
V. cristinaeRuiz-Sánchez et al., 2012; type locality and type level: Pico del Fraile 2, Ebro Basin, Spain; MN4/5, Miocene.
V. rambliensisRuiz-Sánchez et al., 2012; type locality and type level: Pico del Fraile 1, Ebro Basin, Spain; Upper Ramblian, MN3, zone A, Early Miocene
V. bergasensisRuiz-Sánchez et al., 2014; type locality and type level: Bergasa, Ebro Basin, Spain; MP30, Late Oligocene (ref. Lacomba, 1988).
Apart from the above listed species,Nievella mayriDaams, 1976 from the Early Miocene of Cetina de Aragón, the early Late MioceneRamys multicresatus(de Bruijn, 1966) andMyolidusmay belong toVasseuromys, as so suggested by Agusti et al. (2011).
The taxonomy, phylogeny and biostratigraphy of the known species ofVasseuromysare beyond the scope of this study. Our focus is on whether the Junggar form belongs toVasseuromys, a potential assignment we have indicated in our previous studies (Meng et al., 2001, 2006; Ye et al., 2001a, b; 2003a, b).
After intensive survey of the literatures aboutVasseuromys, we realized that, in general, all European species have features in common with the type speciesV. rugosusand differ from those of the TurkishV. duplexandVasseuromysaff.V. duplex. We recognize the fundamental difference between them as representing two dental patterns ofVasseuromyscheek teeth, primarily re fl ected in the upper cheek teeth, which are differentiated below:
In European species ofVasseuromys, the anterotrope and posterotrope outside the trigon are usually absent; if present, they are short and weak; the ridges are rugose and asymmetrically arranged.
InV. duplexandVasseuromysaff.V. duplexfrom Anatolia of Turkey, the anterotrope and posterotrope outside the trigon are always present and long; they extend almost the full length of the corresponding valleys they are in; the ridges are regular and symmetrically arranged.
In fact, Ünay (1994:470) already noted that “The striking features of TurkishVasseuromysare the ever present long extra ridges in the anterior and posterior valleys outside the trigon on the M1/2… different from all European species.” We found that the pattern of the anterotrope and posterotrope is present not only on M1/2 but also on P4 and M3 of TurkishVasseuromys(Ünay, 1994). Because of the distinct morphological differences between the Turkish and EuropeanVasseuromysspecies, we think thatV. duplexshould be inevitably assigned to a different genus. The Junggar and Turkish forms are also different fromGlirulus, as we discussed above.
In addition to the tooth crown structures, we have compared the root number of various species ofGlirulus(Paraglirulusincluded),GliruloidesandVasseuromys(Table 1). All upper cheek teeth of the three genera are triple-rooted except thatGlirulus(Paraglirulus)werenfelsihas a double-rooted P4. However, the available data show that the lower cheek teeth of allGlirulusspecies are double-rooted except that p4 ofGlirulus(P.)agelakifrom Aliveri issingle-rooted andGlirulus japonicushas double-rooted p4 and triple-rooted lower molars.Gliruloideshas also double-rooted lower cheek teeth except that the m3 ofGliruloidesaff.G. duplexis occasionally triple-rooted. Species ofVasseuromys, where the root condition is known, however, have a single-rooted p4 and double or triple-rooted lower molars. Table 1 shows that the Junggar form is similar toG. duplexnot only in their tooth crown structures but also in the root number. However, it should be noted that only the m3 root number (two or three roots) ofGliruloidesaff.G. duplexwas provided by Ünay (1994), whereas the root number of m1/2 was not provided. With the survey of the root number distributions among relevant species, we consider the root condition to be of taxonomic information for species of the three genera in question, although more data are needed to verify this consideration.
In short, given that the Junggar and Turkish forms share some distinctive dental features but differ from theVasseuromysandGlirulusin various features, we think they should beplaced in the same genus; thus we establish the new genusGliruloidesto include the Junggar and Turkish forms.
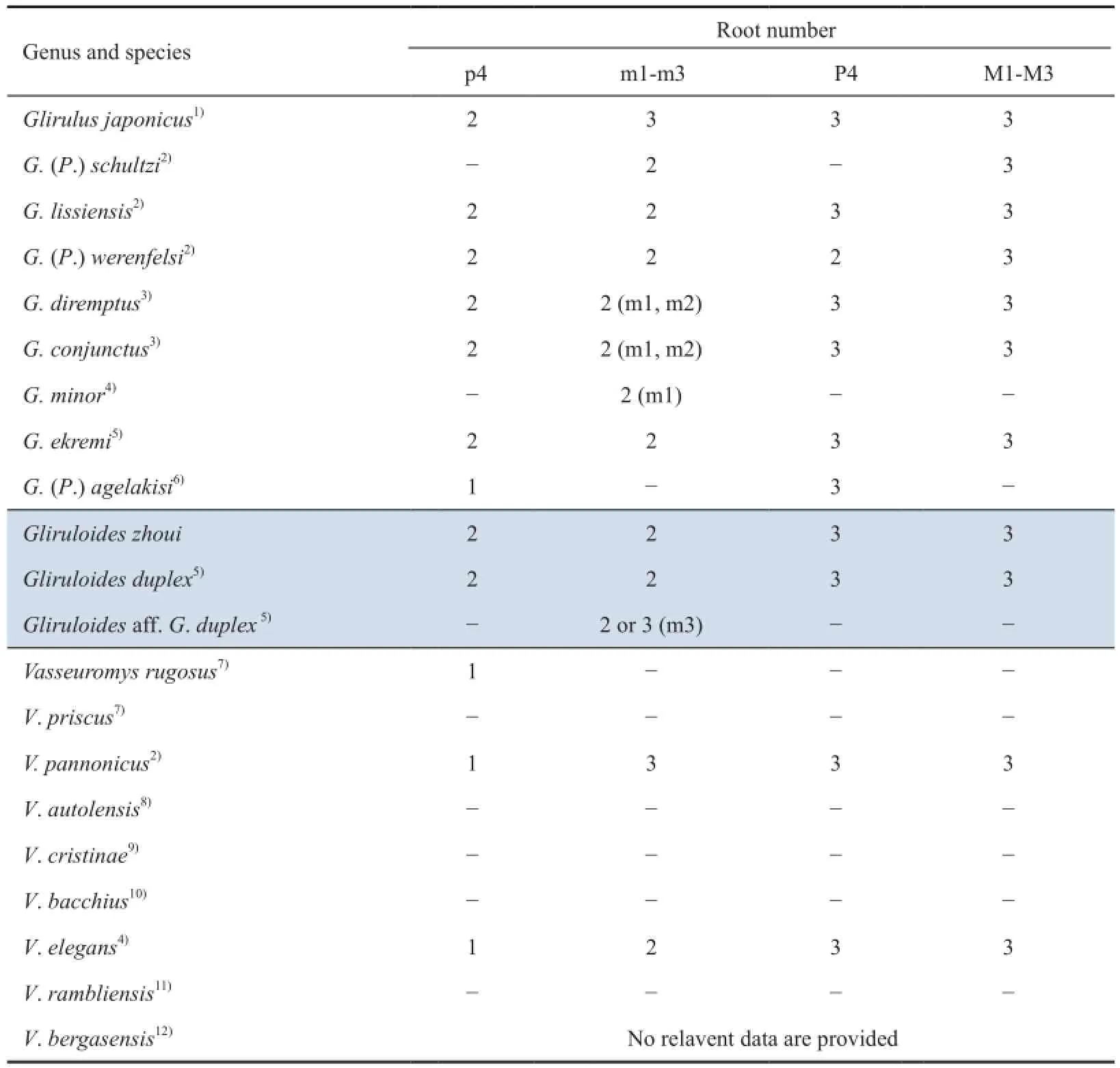
Table 1 Comparisons of cheek teeth root number ofGlirulus,GliruloidesandVasseuromys
Gliruloidescurrently includes three species:G. zhoui,G. duplexandGliruloidesaff.G. duplexand tentatively assigned to the subfamily Dryomyinae because of several diagnostic features, such as the concaved surface, basically symmetrical upper molar, and some teeth with nearly complete endoloph.
The Origin ofGliruloidesThe Early Oligocene (ca. MP25)Glis guerbuezifrom Thrace of Turkey (Ünay-Bayraktar, 1989) is, to our knowledge, the earliest known dormouse that displays considerable resemblance toGliruloidesin dental pattern.G. guerbuezihas at most nine transverse ridges in either upper or lower molars (see Fig. 4C). On certain M1/2s ofG. guerbuezi, besides four main ridges, there are precentroloph, postcentroloph and prototrope inside the trigon, and anterotrope and posterotrope outside the trigon. In addition to four main ridges and centrolophid, the anterotropid and posterotropid are developed in m1 and m2, with incipient metatropid and mesotropid present in m2. M1/2 ofG. guerbuezialso has V-shaped or narrow U-shaped trigon and an incomplete endoloph, and the anterolophid is labially isolated in lower molars. It is probable thatGliruloideswas derived from aGlis guerbuezilike ancestor, although we are not able to illustrate their detailed evolutionary process because of the limited knowledge available to us. Together withGliruloidesduplexandGliruloidesaff.G. duplex, Ünay (1994) also described two species ofGlirulus:G. ekremiof MN3 (from Keseköy) andGlirulusaff.G. ekremiof MN2 (from Harami 1, associated withV. duplex)) from Anatolia. We have noticed that the TurkishG. ekremihas an incomplete endoloph in 27.50% of M1/2 specimens and a precentroloph connected to the endoloph in only 19.2% of M1/2s, and the lower molars ofG. ekremiare morphologically very similar to those of the Turkish “Vasseuromys”. Thus, identi fi cation of those teeth is not always unquestionable. It is also reasonable to infer thatGlirulusmay also be derived from the sameGlis guerbuezi-like ancestor so thatGliruloidesandGlirulusshare a most recent common ancestor but evolved in different directions as two lineages. The fact that the root number of the mostGlirulusspecies is same as that ofGliruloides, as shown in Table 1, could be another evidence for their common origin.
Ecology According to Walker (1975: volume 2:979) the livingGlirulus japonicusinhabits mountain forests from about 400 to 1800 meters in elevation, with the highest recorded elevation being 2900 meters. This animal’s diet includes fruits, seeds, insects and bird’s eggs. All fossil specimens ofGliruloidesduplex,Gliruloidesaff.G. duplexand two species ofGlirulusÜnay (1994) described were from Lower Miocene lignite containing sections (de Bruijn and Saraç, 1991) of Anatolia (from localities Keseköy, Harami 1-3 and Kilçak 0, 0", 3A, 3B). The morphological similarity of teeth inGliruloidesandGlirulusfrom Turkey could be attributed to a similar ecological environment (ecotope) and diet. The presence of lignite in the sections probably indicates wet biotopes (de Bruijn and Saraç, 1991; Ünay, 1994).Gliruloides zhouiis a member of the Late Oligocene Tieersihabahe-Izone fauna, collected from the same localities (XJ 20004 and XJ 98024) as the giant rhinoParaceratherium sui(Ye et al., 2003a) but was discovered from the level immediately above the level where rhinoceros located. Three rhino localities were found in Ulungur River area, which arrange in a west-east extending line with the largest distance of ca. 57 kilometers.Paraceratherium suiwas discovered from the fl uvial sediments at the base of Tieersihabahe Formation. Ye et al. (2012:1530) inferred that the presence of this giant rhino, at the time of ca. 25 Ma, is an implication of a temperature rise in the course of global cooling and aridi fi cation that begins from the Early Oligocene, which is coincided with the time of “Late Oligocene warming” (Zachos et al., 2001: fig. 2).Gliruloides zhouimight still lived in a relatively wet and warm transitional time somewhat later thanParaceratherium sui. We have mentioned above thatGliruloidescould be derived from theGlis guerbuezi-like form. The latter came from the Turkish late Early Oligocene localities situated in the Lignite-Sandstone Formation (Ünay-Bayraktar, 1989:10). The sediments could also indicate that wet and forest environment existed during that period in this regional area.
Acknowledgements We thank Dr. Ruiz-Sánchez from Department of Geology, Division of Paleontology, University of Valencia of Spain for providing us important literatures about spanishVasseuromys. Thanks are also due to the American Museum of Natural History, New York for permission to use the scanning electron microscope. We are grateful to Su Jianfen and Wu Shaoyuan for their hard work in the fi eld and to Yue Qiwan for sorting the miniscule teeth from the concentration of screenwashed samples. We also thank many local peoples who helped us in collecting, transporting and screenwashing dirt samples under the baking sun of the desert. This research has been supported by the Strategic Priority Research Program of Chinese Academy of Sciences (CAS, XDB03020501) and National Basic Research Program of China (2012CB821904).
Agustí J, Pérez-Rivarés F J, Cabrera L et al., 2011. The Ramblian-Aragonian boundry and its signi fi cance for the European Neogene continental chronology. Contributions from the Ebro Basin record (NE Spain). Geobios, 44(2-3): 121-134
Alvarez-Sierra M A, Daams R, Lacomba J I et al., 1990. Paleontology and biostratigraphy (micromammals) of the continental Oligocene-Miocene deposits of the North-Central Ebro Basin (Huesca, Spain). Scripta Geol, 94: 1-77
Baudelot S, Bonis L de, 1966. Nouveaux Gliridés (Rodentia) de l’Aquitanien du basin d’Aquitaine. C R Soc Géol France, 8: 303-304
Bonis L de, 1973. Contribution a l’etude des mammifères de l’Aquitanien de l’Agenais: Rongeurs-Carnivores-Perissodactyles. Mém Mus Natl Hist Nat, Ser C, 28: 1-192
Bruijn H de, 1966. On the mammalian fauna of theHipparion-beds in the Calatayud-Teruel Basin (Prov. Zaragoza, Spain). The Gliridae. Proc K Ned Akad Wet, Ser B, 69(3): 367-387
Bruijn H de, Saraç G, 1991. Early Miocene rodent faunas from the eastern Mediterranean area, Part I. The genusEumyarion. Proc K Ned Akad Wet, Ser B, 94(1): 1-36
Cuenca G, 1985. Los Roedores (Mammalia) del Mioceno Inferior de Autol (La Rioja). Cienc Tierra, 2: 1-96
Daams R, 1976. Miocene rodents (Mammalia) from Cetína de Aragon (Prov. Zaragoza) and Buñol (Prov. Valencia), Spain. Proc K Ned Akad Wet, Ser B, 79(3): 152-182
Daams R, Bruijn H de, 1995. A classi fi cation of the Gliridae (Rodentia) on the basis of dental morphology. Histryx, 6(1-2): 3-50
Daxner-Höck G, Bruijn H de, 1981. Gliridae (Rodentia, Mammalia) des Eichkogels bei Mödleng (Niederösterreich). Paläont Z, 55(2): 157-172
Daxner-Höck G, Höck E, 2009. New data on Eomyidae and Gliridae (Rodentia, Mammalia) from Late Miocene of Austria. Ann Naturhist Mus Wien, 111A: 375-444
Freudenthal M, 2004. Gliridae (Rodentia, Mammalia) from the Eocene and Oligocene of the Sierra Palomera (Teruel, Spain). Treb Mus Geol Barcelona, 12: 97-173
Kawamura Y, 1989. Quaternary rodent faunas in the Japanese Islands (part 2). Mem Fac Sci, Kyoto Univ, Ser Geol Mineral, 54(1-2): 125-157
Kretzoi M, 1978. Wichtigere Streufunde in der Wirbeltierpaläontologischen Sammlung der Ungarischen Geologischen Anstalt. Áll Földt Int Évi Jel, 1978: 348-358
Lacomba J I, 1988. Estudio de las faunas de micromamiferos del Oligoceno superior y mioceno inferior en las cuencas deLoranca, Ebro riojano y Ebro aragonés. Aspectos paleoecológicos. Doctoral thesis. Madrid: Madrid Universidad Complu tense de Madrid. 1-389
Martínez-Salanova J, 1987. Estudio paleontológico de los Micromamiferos del Mioceno inferior de Fuenmayor (La Rioja). Cienc Tierra, 10: 1-99
Mayr H, 1979. Gebissmorphologische Urtersuchungen an miozaenen Gliridae (Mammalia, Rodentia) Süddeutshlands. Inaugural Dissertation. München: Ludwig-Maximilians-Universität. 1-380
Meng J, Ye J, Wu W Y et al., 2001. Two petrosals of gliriform mammals from Late Oligocene of Tieersihabahe, Xinjiang Uygur Autonomous Region, China. Vert PalAsiat, 39(1): 43-53
Meng J, Ye J, Wu W Y et al., 2006. A recommended boundary stratotype section for Xiejia Stage from northern Junggar Basin: implications to related bio-chronostratigraphy and environmental changes. Vert PalAsiat, 44(3): 205-236
Meulen A J van der, Bruijn H de, 1982. The mammal from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 2, The Gliridae. Proc K Ned Akad Wet, Ser B, 85(4): 485-524
Ruiz-Sánchez F J, Murelaga X, Freudenthal M et al., 2012a. A new species of glirid rodentVasseuromys(Gliridae, Rodentia) from the Aragonian (Miocene) of the Ebro Basin (north-eastern Spain). Acta Palaeont Pol, 57(2): 225-239
Ruiz-Sánchez F J, Murelaga X, Freudenthal M et al., 2012b.Vasseuromysrambliensissp. nov. (Gliridae, Mammalia) from the Ramblian (Lower Miocene) of the Tudela Formation (Ebro Basin, Spain). Paleont Electron, 15(1), 4A: 1-16
Ruiz-Sánchez F J, Lacomba-Andueza J I, Freudenthal M et al., 2014. A new species ofVasseuromys(Gliridae, Mammalia) from the Upper Oligocene of the Ebro Basin (Spain). Paläont Z, 88(1): 73-84
Ünay E, 1994. Early Miocene rodent faunas from the eastern Mediterranean area. Part IV. The Gliridae. Proc K Ned Akad Wet, Ser B, 97(4): 445-490
Ünay-Bayraktar E, 1989. Rodents from the Middle Oligocene of Turkish Thrace. Utrecht Micropal Bull Spec Publ, 5: 1-119
Walker E P, 1975. Mammals of the World. 3rd ed. Baltimore and London: The Johns Hopkins University Press. 1-1500
Wu W Y, 1993. Neue Gliridae (Rodentia, Mammalia) aus untermiozanen (orleanischen) Spaltenfüllungen Süddeutschlands. Doc Nat, 81: 1-157
Wu W Y, Ye J, Bi S D et al., 2000. The discovery of Late Oligocene dormice from China. Vert PalAsiat, 38(1): 36-42
Ye J, Meng J, Wu W Y, 2003a. Discovery ofParaceratheriumin the northern Junggar Basin of Xinjiang. Vert PalAsiat, 41(3): 220-229
Ye J, Meng J, Wu W Y, 2003b. Oligocene/Miocene beds and faunas from Tieersihabahe in the northern Junggar Basin of Xinjiang. Bull Am Mus Nat Hist, 279: 568-585
Ye J, Wu W Y, Meng J, 2001a. Tertiary stratigraphy in the Ulungur River Area of the northern Junggar Basin of Xinjiang. J Stratigr, 25(3): 193-200
Ye J, Wu W Y, Meng J, 2001b. The age of Tertiary strata and mammal faunas in Ulungur River Area of Xinjiang. J Stratigr, 25(4): 283-287
Ye J, Wu W Y, Ni X J et al. 2012. The Duolebulejin section of northern Junggar Basin and its stratigraphic and environmental implication. Sci Sin Terrae, 42: 1523-1532
Zachos J, Pagani M, Sloan L et al., 2001. Trends, rhythms and aberrations in global climate 65 Ma to Present. Science, 292: 686-693
新疆准噶尔盆地北缘晚渐新世睡鼠再研究
吴文裕1孟 津1,2叶 捷1倪喜军1毕顺东1,3
(1 中国科学院古脊椎动物与古人类研究所,中国科学院脊椎动物演化与人类起源重点实验室 北京 100044)
(2 美国自然历史博物馆古生物学部 纽约 10024)
(3 美国宾夕法尼亚州印第安纳大学生物系 印第安纳 PA 15705)
周氏似日本睡鼠Gliruloides zhoui是发现于新疆准噶尔盆地北缘晚渐新世铁尔斯哈巴合哺乳动物组合I带的一个化石新属种。新属似日本睡鼠Gliruloides的属征为:中等大小;颊齿咀嚼面凹;上、下颊齿通常具9条主要横脊,有时具次级附脊;上颊齿的前边附脊和后边附脊以及下颊齿的下前边附脊和下后边附脊都很发育,几乎占据了其所在齿谷的整个长度。上颊齿的横脊唇端趋于游离。M1和M2具V形或窄U形三角座,内脊不完整或近于完整,前中央脊不与内脊相连。下颊齿的下内脊通常不连续;下臼齿的下前边脊在唇端稍向后弯,但不与原尖相连。p4, m1-m3, P4, M1-M3的齿根数分别为2, 2, 3, 3。新属与Glirulus在形态上相似,但Glirulus的上颊齿的三角座均为宽U型,具有完整的内脊,前中央脊通常与内脊相交,横脊唇端通常不游离。新属与Vasseuromys属的最主要形态差异在于上颊齿具有很发育的前边附脊和后边附脊,而后者上颊齿的前边附脊和后边附脊通常缺失或很不发育。归入该属的种还有土耳其早中新世的Vasseuromysduplex和Vasseuromysaff.V. duplex。土耳其Thrace早渐新世的Glis guerbuezi很可能是Gliruloides和Glirulus的共同祖先类型。Gliruloides可能生活于温湿的生态环境。
准噶尔盆地北缘,晚渐新世,铁尔斯哈巴合哺乳动物组合I带,睡鼠
Q915.873
A
1000-3118(2016)01-0036-15
2015-07-22
Wu W Y, Meng J, Ye J et al., 2016. Restudy of the Late Oligocene dormice from northern Junggar Basin. Vertebrata PalAsiatica, 54(1): 36-50
中国科学院战略性先导科技专项(编号:XDB03020501)和国家重点基础研究发展计划项目(编号:2012CB821904)资助。
猜你喜欢
百花园(2022年6期)2022-12-28 04:49:34
房地产导刊(2022年5期)2022-06-01 06:19:24
药学实践杂志(2021年4期)2021-07-28 07:19:04
西南石油大学学报(自然科学版)(2018年5期)2018-11-06 06:46:06
民族音乐(2018年2期)2018-05-26 03:04:34
新疆地质(2016年4期)2016-02-28 19:18:43
石油知识(2016年2期)2016-02-28 16:19:48
新疆地质(2015年3期)2015-12-10 05:08:27
天然产物研究与开发(2015年11期)2015-01-11 04:39:34
首都经济贸易大学学报(2013年4期)2013-03-11 18:05:36

- 古脊椎动物学报(中英文)的其它文章
- A new type of dinosaur eggs from Early Cretaceous of Gansu Province, China
- A new hadrosauroid dinosaur from the Late Cretaceous of Tianzhen, Shanxi Province, China
- Female preference promotes asynchronous sex evolution in Elephantiformes
- New record of a haplocyonine amphicyonid in early Miocene of Nei Mongol fi lls a long-suspected geographic hiatus
- Morphology and taxonomy of Gazella (Bovidae, Artiodactyla) from the Late Miocene Bahe Formation, Lantian, Shaanxi Province, China