
大黄属(蓼科)植物ndhF基因的适应性进化
2016-03-21 05:54:35李景剑刘合霞毛世忠黄仕训
广西植物 2016年1期
李景剑,刘合霞,毛世忠,赵 博,3,黄仕训*
( 1.华南农业大学林学与风景园林学院,广州510642; 2.广西壮族自治区广西植物研究所,中国科学院广西桂林541006; 3.中国中医科学院中药研究所,北京100700 )
大黄属(蓼科)植物ndhF基因的适应性进化
李景剑1,2,刘合霞2,毛世忠2,赵博2,3,黄仕训2*
( 1.华南农业大学林学与风景园林学院,广州510642; 2.广西壮族自治区广西植物研究所,
中国科学院广西桂林541006; 3.中国中医科学院中药研究所,北京100700 )
摘要:大黄属( Rheum L.)是蓼科( Polygonaceae)中一个高度分化的大属,广泛分布在亚洲和欧洲的高山和沙漠地区,全世界约60种,其中在青藏高原及其邻近地区发现了约40种。该属种的高度分化曾被推测是第三纪末青藏高原的快速隆升以及第四纪气候的反复变化所引发的适应性辐射导致。为进一步了解大黄属植物辐射式物种分化的分子适应机制,该研究选取34个形态上多样化的大黄属物种,利用系统发育分析软件,在时间框架下采用位点模型和分支模型对大黄属的叶绿体ndhF基因进行了适应性进化分析。结果表明:大黄属植物的分
李景剑,刘合霞,毛世忠,等.大黄属(蓼科)植物ndhF基因的适应性进化[J].广西植物,2016,36( 1) : 101-106
LI JJ,LIU HX,MAO SZ,et al.Adaptive evolution of the ndhF gene in the genus Rheum ( Polygonaceae)[J].Guihaia,2016,36( 1) : 101-106
关键词:大黄属(蓼科),ndhF基因,分支模型,位点模型,正选择位点
The genus Rheum ( Polygonaceae) with about 60 species,primarily distributed in mountainous and desert regions of the Qinghai-Tibetan Plateau and adjacent areas ( Kao&Cheng,1975; Li,1998).The distribution and the ancestral area reconstruction analyses consistently suggested that rapid radiations of Rheum have occurred,and may have been caused by the extensive uplifts of the Qinghai-Tibetan Plateau ( Wang et al,2005; Wan et al,2011; Sun et al,2012).To adapt to the new alterations of habitat,morphological traits of this genus are highly diversified.Some species have evolved into dwarf plants with coriaceous basal leaves or drooping bracts to defense them from freeze injury and could distribute up to snow line at altitude of 5 400 m,for example R.Nobile ( Xie,2000).For other species,stem leaves are have degenerated and basal leaves are covered with verruca or indumentum to reduce water transpiration,to avoid the burning from high temperature and to avoid damage by strong winds,therefore,these species can grow in the Gobi Desert at altitude 700 m.R.palaestinum in particular,has broad,rigid leaves,with a waxy surface,and channels cut into them that funnel any water that drops onto them toward its root,with enough force to cause deep soil penetration ( Lev-Yadun et al,2009).These changes in morphology and physiology might be resulted from the adaptive evolution of some genes which encode functional proteins,such as chloroplast ndhF gene that related to photosynthesis and photorespiration ( Zapata et al,2005).
Chloroplast is thought to be a very conservative part of plant genome but little is known about the evolution of this plastome promoters.Previous study showed that the alignment of sequences upstream ndhF suggested that promoters of this gene underwent comparatively rapid evolution in flowering plants ( Seliverstov et al,2009).The ndhF gene is located in a small single-copy region of the chloroplast genome that rarely underwent substantial rearrangements in terrestrial plants ( Hiratsuka et al,1989).Its nucleotide sequence predicts a hydrophobic protein of 664 amino acids with a calculated mass of 72.9 kDa ( Schluchter et al,1993).The ndhF gene encodes NADH dehydrogenase F subunit of the plastid NDH complex which regulated the activity of NDH complexes by its phosphorylation.The plastid NDH complex in chloroplast thylakoid membranes is involved in photosystem I cyclic and chlororespiratory electron transport in photosynthetic regulation of higher plants ( Lascano et al,2003).
Considering the adaptability of Rheum to extreme habitats,the sequences of ndhF gene from 34 species of Rheum and 2 species of Oxyria in Polygonaceae were retrieved from the National Center for Biotechnology Information ( NCBI) for adaptive evolution testing in this study.Our finding may provide new molecular evidence for the rapid putative radiations of Rheum triggered by the recent uplifts of the Qinghai-Tibetan Plateau.
1 Materials and Methods
Thirty-four species of Rheum and two species of Oxyria in Polygonaceae used in this study were listed in Table 1.Sequences of ndhF gene were downloaded from NCBI ( http: / /www.ncbi.nlm.nih.gov/guide/).Sequence alignments were conducted using the software CLUSTAL W ver.1.83 ( Thompson et al,1994) and adjusted manually in BioEdit 5.0.9.1 ( Hall,1999).Oxyria digyna and O.sinensis were used as outgroup.Maximum Parsimony ( MP) analysis was conducted using PAUP 4.0b10 ( Swofford,2003),Heuristic searches were conducted 1 000 times with random taxon-addition sequences,with tree-bisection-reconnection ( TBR) branch swapping,and with the options MULPARS in effect and STEEPEST DE-SCENT off.Support for internal nodes was estimated with bootstrap values ( Felsenstein,1985).

Table 1 Species names and accession numbers of ndhF sequences
Based on the MP tree,the analysis of adaptive evolution of ndhF gene was implemented in the program of CODEML from PAML package version 4 ( Yang,2007).The lnL values under one-ratio model as well as free ratio model were calculated,and the Likelihood Ratio Test ( LRT) was conducted to test whether there were different ratios for each lineage.Site-specific models,which allowed the ω ratio to vary among sites but fixed a single ω ratio in all branches,were used to detect positive selection and to identify positively selected sites.Three pairs of site-specific models were calculated to test for recurrent,diversifying,selection: M0 ( one ratio) and M3 ( Discrete),M1 ( Neutral) and M2 ( Selection),and M7 ( Beta) and M8 ( Beta&ω).( Yang&Nielsen,2002; Yang et al,2005).Log likelihoods of models ( M1 vs.M2; M0 vs.M3; M7 vs.M8) were compared using LRT.
For the spatial analysis of the codon site under positive selection,the PSIPRED server ( http: / /bioinf.cs.ucl.ac.uk/psipred/) was used to analysis the secondary structure of NDHF subunit for Rheum palaestinum.
2 Results and Analysis
The ndhF dataset had an aligned length of 1 944 characters in the dataset,of which,1 751 characters were constant,111 were variable and parsimony-uninformative,and 82 were parsimony-informative.Maximum Parsimony analysis yielded 84 equally parsimonious trees,and a strict consensus tree of these trees was shown in Fig.1.The topology of MP tree was consistent with the molecular phylogenies published to date ( Wang et al,2005; Sun et al,2012).The consensus tree revealed three major clades ( A,B,C) within the genus Rheum,all species of Rheum comprised a well-supported lineage,with a sister relationship to Oxyria.
To analyze the possibility that positive selection acts on ndhF genes,we used the maximum-likelihood codon model from the CODEML program in the PAML4 package.The topology of the MP tree mentioned above was modified for all CODEML analyses.All calculations and tests are listed in Tables 2 and 3.Under the one-ratio model which allowed for only a single ω ratio across all sites of the gene phylogeny and the same ω ratio for all branches in the phylogenic tree ( Fig.1),the loglikelihood value was ω= 0.296 5,lower than 1 ( Table 2).In the branch-specific analysis,the LRT statistic for the comparison of the one-ratio model vs.the free-ratio model was 2Δ=77.467 0 with P<0.05 and df=54,suggesting that there had different ratios for each lineage of Rheum species ( Table 3).But no sites with a Bayesian posterior probability of positive selection larger than 0.95 in one or more cases were found when analyzed by Bayes.
In site-specific models,models M2,M3 and M8 allowed sites with ω>1.The LRT statistic of M0-M3,M1-M2 and M7-M8 comparison all with P<0.05,so models M3,M2 and M8 was significantly better than M0,M1 and M7.Under both M2 and M8 models,three sites were under positive selection with ω>1 and identified three NDHF residues ( 188H,465H,551L) with a Bayesian posterior probability of positive selection larger than 0.95 in one or more cases when analyzed by Empirical Bayes ( Table 2).

Fig.1 Strict consensus tree from maximum-parsimony ( MP) analysis based on ndhF sequences of Rheum Numbers above the branches indicate bootstrap values by MP analysis.Numbers with bootstrap values>50% are shown.Shadows on the right indicatefive clades of Rheum displaying rapid radiation.
With no obvious sequence similarity to structures present in PDB,the secondary structure of NDHF subunit for Rheum palaestinum was predicted by PSIPRED server ( Fig.2).The test results showed the 188th amino acid which located in the α-helix was histidine ( H) in R.palaestinum,while asparagine ( N) was also found in other species of Rheum.The amino acid encoded by the 465th codon was histidine ( H) in R.palaestinum,while asparagine ( N) and tyrosine ( Y) was found in other species of Rheum.In addition,the 551th codon encoded was leucine ( L) in R.palaestinum,while phenylalanine ( F) was found in other species of Rheum.
3 Discussion
The distribution and the ancestral area reconstruction analyses suggests that rapid putative radiations of Rheum might have been triggered by the recent uplifts of the Qinghai-Tibetan Plateau and the Quaternary climate oscillations.Geological evidence indicates that at least four different periods at the early Miocene ( i.e.,22,15 -13,8-7,and 3.5-1.6 Ma) occurred during recent extensive uplifting of the Qinghai-Tibetan Plateau ( Shi et al,1998; Sun et al,2012),and new habitats may have been created while old ones became fragmented within each period.The new alterations of habitat of Rheum species are various,from snow line at altitude 5 400 m to Gobi Desert at altitude 700 m.Through the adaptive evolution of ndhF gene involved in photosynthesis pathways,some species of Rheum could adapt the extreme habitats.
By comparing Models M1a/M2a and M7/M8,threeamino acid sites ( 188H,465H,551L) were identified under positive selection.The secondary structure of NDHF subunit showed thatthe positively selected sites ( 465H and 551L) were on the loops.The 188th amino acid which located in the α-helix was histidine ( H) in R.palaestinum ( Fig.2),while asparagine ( N) was found in other species.The ndhF gene encoded a subunit of the plastid NDH complex,and this complex assembly might be regulated on the pos-transcriptional level in a way that the quantity of whole NDH complexescould be determined by the quantity of one of its subunits,e.g.NDHF.The activity of NDH complexes was also regulated by phosphorylation of the NDHF polypeptide.Our study found that the ndhF gene was at high expression level under stress conditions,and those stress factors from environment might be the selective pressure to lead the adaptive evolution of ndhF gene.Our results indicated that the change of spatial structure may have a relationship with the adaptation of Rheum to the environment.For example,R.palaestinum with histidine mutation on the 188th codon site is the rare Rheum plant growing in mountainous desert areas ( receiving an average annual rainfall of ca.75 mm) in the world,and it has a single deep main vertical root ( Zohary,1966).Previous studies released that histidine is one of the essential amino acids for plant growth and survival,especially for root meristem maintenance,the higher histidine content in plant,the faster root growth and better adaptability to the environments ( Mo,2006; Malki&Jaeobs,2001).So the adaptive evolution of ndhF subunit might be important for Rheum species to adapt various habitats.
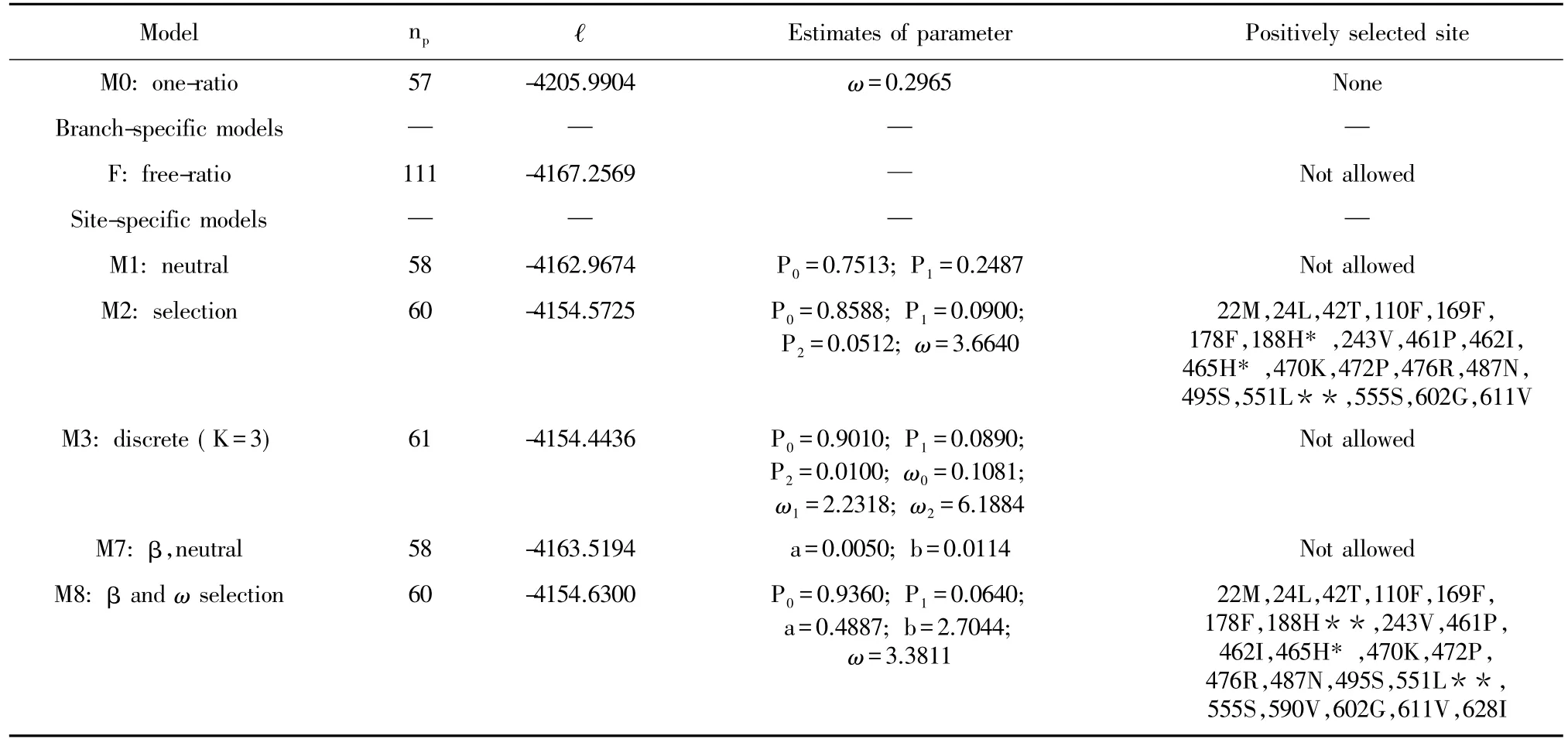
Table 2 Maximum likelihood parameter estimates for ndhF gene

Fig.2 The secondary structures of NDHF protein for Rheum palaestinum The sites ( 188H,465H,551L) under adaptive evolution are marked with red boxes.

Table 3 Likelihood ratio test ( LRTs) of the variable ω ratios under different models for ndhF gene
AcknowledgementsWe are grateful to LIU Lei for his help with data analysis and Qian Guo for language editing support.
References:
FELSENSTEIN J,1985.Confidence limits on phylogenies: an approach using the bootstrap[J].Evolution,39: 783-791.
HALL TA,1999.BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT[M].Nucl Acids Symp Ser,41: 95-98.
HIRATSUKA J,SHIMADA H,WHITTIER R,et al,1989.The complete sequence of the rice ( Oryza sativa) chloroplast genome: Intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals[J].Mol Gen Genet,217: 185-194.
KAO TC,CHENG CY,1975.Synopsis of the Chinese Rheum[J].Acta Phytotax Sin,13: 69-82.
LASCANO HR,CASANO LM,MARTIN M,et al,2003.The activity of the chloroplastic ndh complex is regulated by phosphorylation of the NDH-F subunit[J].Plant Physiol,13: 2 256-2 262.
LEV-YADUN S,KATZIR G,NEEMAN G,2009.Rheum palaestinum ( desert rhubarb),a self-irrigating desert plant[J].Beijing: Naturwissenschaften,96: 393-397.
LI AR,1998.Flora republicae popularis sinicae[M].Beijing: Beijing Science Press.
MALKI FE,JAEOBS M,2001.Molecular characterization and expression study of a histidine auxotorphic mutant ( hisl-) of Nicotiana plumbaginifolia[J].Plant Mol Biol,45: 191-199.
MO XR,2006.The functional analysis of Arabidopsis HPA1 gene [D].Hangzhou: Zhejiang University.
SELIVERSTOV AV,LYSENKO EA,LYUBETSKY VA,2009.Rapid evolution of promoters for the plastome Gene ndhF in flowering plants[J].Russ J Plant Physl,56( 6) : 838-845.
SCHLUCHTER WM,ZHAO JD,BRYANT DA,1993.Isolation and characterization of the ndhF gene of Synechococcus sp.Strain PCC 7002 and initial characterization of an interposon mutant [J].J Bacteriol,175( 11) : 3 343-3 352.
SHI YF,LI JJ,LI BY,1998.Uplift and environmental changes of Qinghai-Tibetan Plateau in the Late Cenozoic[M].Guangzhou: Guangdong Science and Technology Press.
SUN Y,WANG A,WAN D,et al,2012.Rapid radiation of Rheum ( Polygonaceae) and parallel evolution of morphological traits [J].Mol Phylogenet Evol,63: 150-158.
SWOFFORD DL,2003.PAUP* : Phylogenetic analysis using parsimony ( * and other methods)[M].Sunderlande,MA: Sinauer Associates.
THOMPSON JD,GIBSON TJ,PLEWNIAK F,et al,1997.The ClustalX Windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools[M].Nucleic Acids Res,24: 4 876-4 882.
WAN D,WANG A,ZHANG X,et al,2011.Gene duplication and adaptive evolution of the CHS-like genes within the genus Rheum ( Polygonaceae)[J].Biochem Syst Ecol,39: 651-659.
WANG AL,YANG MH,LIU JQ,2005.Molecular phylogeny,recent radiation and evolution of gross morphology of the Rhubarb genus Rheum Polygonaceae inferred from chloroplast DNA trnL-F sequences[J].Ann Bot,96: 489-498.
XIE ZQ,2000.Eco-geographical distribution of the species from Rheum L.Polygonaceae in China.China’s Biodiversity conservation toward the 21stCentury[M].Beijing: China Forest Press: 230-236.
YANG Z,NIELSEN R,2002.Codon-substitution models for detecting molecular adaptation at individual sites along specific lineages[J].Mol Biol Evol,19: 908-917.
YANG Z,WENDY SWW,RASMUS N,2005.Bayes empirical bayes inference of amino acid sites under positive selection[J].Mol Biol Evol,22: 1 107-1 118.
YANG Z,2007.PAML 4: a program package for phylogenetic analysis by maximum likelihood[J].Mol Biol Evol,24: 1 586-1 591.
ZAPATA JM,GUERA A,ESTEBAN-CARRASCO A,et al,2005.Chloroplasts regulate leaf senescence: delayed senescence in transgenic ndhF-defective tobacco[J].Cell Death Diff,12: 1 277-1 284.
ZOHARY M,1966.Flora Palaestina.Vol.I[M].Jerusalem: The Israel Academy of Sciences and Humanities.
*通讯作者:黄仕训,研究员,主要从事濒危植物保护研究,( E-mail) hsx@ gxib.cn。book=102,ebook=107子进化系统树呈现短而平行的辐射式分支式样,显示出典型的物种快速辐射多样化特征;用位点模型检验ndhF基因是否存在经受正向选择(ω>1)时,在氨基酸水平上共鉴定出3个NDHF亚基的正选择位点( 188H,465H,551L),对NDHF亚基的二级结构进行分析后发现编码的188H氨基酸位于α螺旋上。大黄属植物可能通过这些结构域的适应性进化,适应青藏高原的快速隆升以及第四纪气候的反复变化而引发的陆地生态系统改变。该研究结果可为今后对该属植物的实验分析提供首选位点。
作者简介:李景剑( 1983-),男,广东廉江人,博士研究生,主要从事植物分子生物学研究,( E-mail) calljone@163.com。
基金项目:国家科技基础性工作专项( 2009FY120200) ;广西自然科学基金( 2012GXNSFBA053075) ;广西植物研究所基本业务费(桂植业14003) [Supported by the Special Program for Basic Research of Science and Technology of China( 2009FY120200) ; the Natural Science Foundation of Guangxi ( 2012GXNSFBA053075) ; the Science Research Foundation of Guangxi Institute of Botany ( Guizhiye14003)]。
收稿日期:2015-08-24修回日期: 2015-12-02
DOI:10.11931/guihaia.gxzw201508017
