
江淮稻区杂交粳稻骨干亲本产量性状配合力的SSR标记位点鉴定
2016-03-03 09:01党小景刘二宝曾思远洪德林南京农业大学作物遗传与种质创新国家重点实验室江苏南京0095国家粳稻工程技术研究中心天津300457
作物学报 2016年3期
谢 辉 党小景 刘二宝 曾思远 洪德林,*南京农业大学 / 作物遗传与种质创新国家重点实验室, 江苏南京 0095;国家粳稻工程技术研究中心, 天津 300457
江淮稻区杂交粳稻骨干亲本产量性状配合力的SSR标记位点鉴定
谢辉1,2党小景1刘二宝1曾思远1洪德林1,*
1南京农业大学 / 作物遗传与种质创新国家重点实验室, 江苏南京 210095;2国家粳稻工程技术研究中心, 天津 300457
摘要:鉴定杂交粳稻亲本产量性状配合力的标记位点有助于利用分子标记辅助选择技术改良亲本配合力、提高杂交粳稻竞争优势水平。利用9个粳稻BT型不育系和10个恢复系, 按照北卡罗林那设计II (North Carolina Design II)配制90个F1组合,在南京和盱眙两个环境下种植, 测定了各亲本产量性状的配合力和SSR分子标记基因型; 结合二者数据鉴定了6个产量性状配合力的标记位点。结果表明, 在2个环境下综合评价配合力较优的不育系是BT-18A和武羌A, 恢复系是C418。与亲本单株有效穗数、每穗总粒数、每穗实粒数、结实率、千粒重和单株日产量性状配合力显著相关的SSR标记位点, 南京环境下分别检测到8、13、11、6、6和2个; 盱眙环境下分别检测到12、21、8、15、10和7个; 2个环境下都检测到的分别有4、11、5、3、5和1个。标记位点杂合基因型显示正向优势的, 南京环境下占74% (34/46); 盱眙环境下占53% (39/73)。2个环境下都检测到的标记位点中, 有3个各自与3个产量性状配合力共相关; 另有3个各自与2个产量性状配合力共相关; 其余的只与单个产量性状配合力显著相关。数据库检索发现两环境下都检测到的标记位点中, 有10个其附近存在控制相应性状的基因/QTL。讨论了利用鉴定出的标记位点改良粳稻恢复系产量性状配合力的策略。
关键词:杂交粳稻; 骨干亲本; 产量性状配合力; 标记位点基因型
本研究由国家高技术研究发展计划项目(2010AA101301), 高等学校博士学科点专项科研基金(20110097110038, 20130097110001)和中央高校基本科研业务费专项资金项目(KYZ201202-9)资助。
This study was supported by China National High-tech Research and Development Program (2010AA101301), the Doctoral Found of Educational Ministry (20110097110038, 20130097110001) and a grant from Educational Ministry of China for Basic Scientific Research of National Universities (KYZ201202-9).
第一作者联系方式: E-mail: xiehui_2005@126.com
近十年来, 中国杂交粳稻年种植面积34万公顷左右, 占粳稻总面积800万公顷的4%, 与同期全国杂交籼稻年种植面积1650万公顷、占籼稻总面积2200万公顷的75%相比, 还有很大的发展空间[1-4]。长江中下游稻区和黄淮稻区是我国目前最主要的杂交粳稻种植区域, 常年种植面积24万公顷左右。由于高产优质多抗纯系粳稻新品种的不断推出[5], 杂交粳稻的竞争优势百分率逐渐减小, 推广面积难以扩大。造成杂交粳稻发展缓慢的原因主要有竞争优势不强、米质性状欠佳、F1种子纯度波动较大等[6-8],其中竞争优势不强是最主要限制因素, 其主要原因是杂交粳稻育种中缺乏高配合力的恢复系[9]。
国内外大量测恢研究证实粳稻自然品种群体中缺乏有效的育性恢复基因[10-13]。籼稻恢复系中存在两个主效恢复基因位点Rf3和Rf4[14]。杨振玉等[9,15]通过“籼粳架桥”将籼稻中的恢复基因导入粳稻, 解决了三系杂交粳稻恢复系的恢复力问题。但是恢复系的配合力问题一直没有得到很好的解决。由于配合力这一特殊性状需要亲本配组通过F1才能表现出来, 以往的大量关于恢复系配合力研究仅能起到鉴别和指导亲本配组的作用, 但却难以有效直接指导恢复系配合力的改良。分子标记辅助选择育种改变了这一现状。刘小川等最早利用SSR分子标记技术和半双列杂交方法鉴定出籼稻不育系和恢复系亲本间4个有利等位基因和6个杂合等位基因对F1籽粒产量的优势有显著贡献, 6个有利等位基因和6个杂合等位基因显著减少F1籽粒产量优势, 提出把有利等位基因聚合进亲本系、把不利等位基因剔除出亲本系从而培育高产杂种组合的策略[16]。在之后的3项研究中, Liu等[17]鉴定出对籽粒产量优势有利的AFLP标记11个, 不利的8个; 与米质性状亲本配合力显著相关的SSR标记18个[18]; 利用鉴定出的SSR标记辅助改良籼稻恢复系明恢63获得单株谷粒产量性状配合力提高的新恢复系MGH44和MGH45[19]。在杂交粳稻领域, 梁奎等[20]研究了杂交晚粳18个亲本产量及其构成性状优异配合力SSR标记基因型; 黄殿成等[21-22]鉴定了杂交晚粳18个亲本穗长和枝梗数以及10个米质性状优异配合力SSR标记基因型。刘二宝等[23]和刘洋等[24]检测了包含两系亲本在内的20个杂交粳稻亲本农艺性状和品质性状优异配合力SSR标记基因型。为探明杂交粳稻亲本产量相关性状优异配合力标记基因型在不同熟期组合亲本之间的规律,本研究选用目前江淮稻区生产上应用的主要杂交中粳的骨干亲本, 按照NCII (North Carolina Design II)遗传交配设计配制90个F1组合, 种植在两个环境下,调查F1及其亲本产量相关性状, 计算亲本配合力;根据各亲本SSR分子标记基因型和配合力数据, 检测与骨干亲本产量性状配合力显著相关的标记位点,为利用分子标记辅助选择改良江淮稻区杂交粳稻恢复系配合力提供SSR标记信息。
1 材料与方法
1.1供试材料
供试材料为粳稻10个恢复系、9个Boro II细胞质(又称BT型细胞质)的雄性不育系和9个相应的保持系、以及用不育系作母本恢复系作父本按NCII交配设计配制的90个F1杂种。10个恢复系分别是3726R-21、武育粳R-39、157TR-68、C4115、秀水04R、C657、C418、K25、C堡和宁恢8号。9个BT型不育系和9个相应保持系分别是武3A、武3B; 863A、863B; 6427A、6427B; 9201A、9201B; 六千辛A、六千辛B; 武羌A、武羌B; 镇稻88A、镇稻88B; 泗稻8A、泗稻8B; 18A、18B。
1.2组合配制和田间种植及性状调查
2009年正季在南京农业大学江浦试验站水稻田种植10个恢复系、9个不育系和9个保持系。抽穗期对9个不育系人工剪颖与10个恢复系配制90个F1杂交组合。2010年正季将90个杂交组合的F1种子及其对应的恢复系和保持系(因不育系不结实, 用保持系代替)种子分成两份, 一份种植在南京农业大学江浦试验站水稻田(环境1, E1), 另一份种植在国家粳稻工程技术研究中心江苏省盱眙县水稻育种基地(环境2, E2)。两个环境下均于5月10日播种, 6 月13日移栽。每个组合的F1及对应的恢复系和保持系秧苗各种成1个小区, 每个小区种植4行, 每行8穴, 每穴1株; 株距17 cm, 行距20 cm。田间小区
按照完全随机区组设计布置。2次重复。栽培管理同大田。
稻谷成熟后, 先在田间调查单株有效穗数(实粒数在5粒以上的稻穗均记为有效穗)。调查每个小区中间2行共10个单株。然后在室内考查每穗总粒数和每穗实粒数。从调查过单株有效穗数的植株中随机取5株, 计数每株主茎穗(最高穗)的总粒数和实粒数。结实率等于每穗实粒数除以每穗总粒数乘以100。将每个小区去边行收获的所有植株的稻粒风干后, 测小区籽粒产量。以小区籽粒产量除以小区实收株数得单株平均谷粒产量,再除以全生育期天数得单株平均日产量。用风干后的稻粒称千粒重, 对每个田间重复的稻粒样品测定2次。
1.3DNA提取和PCR扩增及扩增产物电泳
2010年7月上旬(水稻分蘖期)剪取9个不育系和10个恢复系的叶片, 按照程宝山等[25]所述的SDS法提取总DNA。按照文献[20]进行PCR扩增, 之后进行电泳、银染和记录。
1.4数据分析
以小区的平均值为单位, 按p×q交配模式进行亲本性状配合力分析[26-27]。用总秩次排序方法[28]综合评价亲本配合力, 即先计算出各亲本产量构成性状的一般配合力(GCA)效应和特殊配合力方差(VSCA), 然后根据各值大小给以不同秩次。6个产量相关性状GCA效应的秩次和VSCA的秩次均按从大到小排序, 值愈大秩次愈小, 越好的亲本其秩次愈小。最后以6个性状的秩次之和(总秩次)作为亲本配合力的评价指标。
采用文献[20]提出的方法筛选亲本性状配合力的标记位点, 即把90个F1组合中单标记位点杂合带型的组合归为一组(杂合带型组), 纯合带型组合归为另一组(纯合带型组), 测验两组之间性状平均数的差异显著性(t测验)。差异显著表示该标记位点与性状的配合力有关。本研究6个产量性状均是越大越好, 标记位点杂合基因型导致性状正向优势的,为优异配合力标记基因型; 导致负向优势的则为不利配合力标记基因型。
标记位点是指能在所用亲本中扩增出多态性产物的引物。标记位点基因型是指同一个引物在不育系中扩增出的条带与恢复系中扩增出的条带的组合。同一标记位点在NCII设计的双列杂交F1群体中有多种标记基因型。假定SSR引物RM1在19个亲本(9个不育系和10个恢复系)之间扩增出100 bp、 200 bp和300 bp 3个条带, 则双列杂交F1群体在该位点就有RM1-100/200、RM1-100/300、RM1-200/300三种杂合的标记基因型和RM100/100、RM1-200/200 和RM1-300/300三种纯合的标记基因型。根据上一段描述的原理, 经过计算确定影响性状配合力的标记位点。参见文献[20], 运用MATLAB语言编制的程序CAScreen1.0在计算机上完成计算过程。
2 结果与分析
2.1两个环境下90个F1组合6个产量性状表现
南京试验点90个F1组合中, 每穗总粒数最多的为352粒, 最少的为145粒, 平均为252粒, 变异系数为16.6%, 表明组合间每穗总粒数有较大的变异。在研究的6个产量性状中, 变异系数最大的是单株有效穗数, 达21.8%; 其次为每穗实粒数(20.4%)。变异系数最小的性状为千粒重, 为5.9%。盱眙试验点90 个F1组合6个性状的最大值和最小值, 除结实率外,均大于南京点相应性状的最大值和最小值。变异系数的变化与南京试验点相似。表明这90个F1组合除结实率外, 其余5个性状在盱眙试验点的表现好于在南京试验点的表现(表1)。
2.210个恢复系和9个不育系6个产量相关性状一般配合力解析
分别对种植在南京和盱眙两个环境下的90个F1组合6个性状表型数据进行方差分析, F测验结果表明6个性状杂交组合间差异均达到显著水平。说明亲本之间各性状配合力存在着真实差异, 可以对这些性状进行亲本一般配合力效应的估算和显著性测验。
在南京环境下, 恢复系中, 单株有效穗数一般配合力(General Combining Ability, GCA)效应最大的是武育粳R-39, 显著大于3726R-21以外的8个恢复系; 每穗总粒数GCA效应最大的是C4115, 显著大于K25和C418以外的7个恢复系; 每穗实粒数GCA效应最大的也是C4115, 显著大于C418以外的恢复系; 结实率GCA效应最大的是157TR-68和武育粳R-39, 显著大于C657、K25、C堡、秀水04R和3726R-21; 千粒重GCA效应最大的是K25, 显著大于C657以外的恢复系; 单株日产量GCA效应最大是C418, 显著大于其他9个恢复系(表2)。不育系中, 单株有效穗数GCA效应最大的是武羌A, 显著大于其余8个不育系; 每穗总粒数GCA效应最大的是18A,显著大于武羌A、镇稻88A和泗稻8A; 每穗实粒数GCA效应最大的是863A, 显著大于镇稻88A和泗稻
8A; 结实率GCA效应最大的是武3A, 显著大于6427A、18A、9201A、镇稻88A和泗稻8A; 千粒重GCA效应最大的是泗稻8A, 显著大于其余8个不育系; 单株日产量GCA效应最大的是18A, 显著大于其余8个不育系(表2)。根据6个产量相关性状的总排序(表2最后1列)和亲本利用价值的评价标准[28], 武3A是最优不育系, 18A、武羌A是较优不育系。恢复系中, C418是最优恢复系, C堡和C4115是较优的恢复系。

表1 两个环境下90个F1组合6个产量相关性状的表现Table 1 Performance of six yield-related traits of 90 F1in two environments
在江苏省盱眙县环境下, 恢复系中, 单株有效穗数GCA效应最大的是武育粳R-39, 显著大于3726R-21以外的8个恢复系; 每穗总粒数GCA效应最大的是C4115, 显著大于其他恢复系; 每穗实粒数GCA效应最大的是C4115, 显著大于C418以外的8个恢复系; 结实率GCA效应最大的是宁恢8号和武育粳R-39, 显著大于其他恢复系(表2)。不育系中, 每穗总粒数GCA效应最大的是18A, 显著大于镇稻88A和泗稻8A以外的不育系; 每穗实粒数GCA效应最大的是6427A, 显著大于六千辛A、武3A和9201A; 结实率GCA效应最大的是武羌A (表2)。综合6个性状加以评判(评判标准同南京环境),盱眙环境下泗稻8A是最优不育系, 18A和武羌A是较优不育系; 武育粳R-39是最优恢复系, 157TR-68、C418和K25是较优恢复系。
2.3两个环境下亲本6个产量性状配合力的标记位点
对10个恢复系和9个不育系亲本的总DNA, 用 115对SSR引物扩增显示, 78对引物在19个亲本之间扩增出多态性产物。用这78个SSR标记位点的亲本基因型数据和F1植株各性状表型数据, 检测与亲本6个性状配合力显著相关的SSR标记位点。与亲本单株有效穗数、每穗总粒数、每穗实粒数、结实率、千粒重和单株日产量性状配合力显著相关的SSR标记位点, 南京环境下分别检测到8、13、11、6、6和2个(表3), 盱眙环境下分别检测到12、21、8、15、10和7个(表4)。标记位点杂合基因型显示正向优势的, 南京环境占74% (34个/46个); 盱眙环境占53% (39个/73个)。
2.3.1南京环境下检测到的与亲本6个产量性状配合力显著相关的标记位点与亲本单株有效穗数配合力显著相关的标记位点鉴定到8个, 涉及第3、第6、第8、第11和第12共5条染色体(表3)。8个标记位点杂合基因型中有3个, 即RM144-230/260、RM247-170/180和RM167-150/160, 显示正向优势(即优异配合力标记基因型), 优势分别为18.6%、26.3%和19.1%。
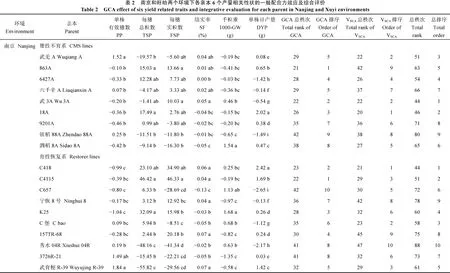


表3 南京环境下检测到的与亲本单个性状配合力显著相关的SSR标记位点Table 3 Marker loci significantly related to combining ability for each trait in parents in Nanjing environment
与亲本每穗总粒数配合力显著相关的标记位点鉴定到13个, 涉及第1、第2、第3、第6、第7、第8和第11共7条染色体(表3)。13个标记位点杂合基因型中11个显示正向优势, 优势范围在13.2%~ 22.0%之间, 优势最大的标记位点杂合基因型是RM345-180/190 (表3)。与亲本每穗实粒数配合力显著相关的标记位点鉴定到11个, 涉及第1、第2、第3、第6、第7、第8和第9共7条染色体(表3)。
11个标记位点杂合基因型中9个显示正向优势, 优势范围在15.2%~24.3%之间。其中, 位点杂合基因型RM341-170/195优势为24.3% (表3)。与亲本结实率配合力显著相关的标记位点鉴定到6个, 涉及第1、第2、第5、第6、第7和第10共6条染色体(表3)。6个标记位点杂合基因型中4个显示正向优势, 优势范围在10.1%~11.9%之间。RM406-145/160可以增加F1结实率优势11.9% (表3)。
与亲本千粒重配合力显著相关的标记位点鉴定到6个, 涉及第2、第3、第7和第8共4条染色体(表3)。6个标记位点杂合基因型中5个显示正向优势, 其中RM10-170/180优势为5.1% (表3)。与亲本单株日产量配合力显著相关的标记位点筛选到2个, 均位于第2染色体。RM208-180/190和RM406-145/160的位点杂合基因型优势分别为17.8%和16.2%。
2.3.2盱眙环境下鉴定出的与亲本6个产量性状配合力显著相关的标记位点鉴定出12个标记位点与亲本单株有效穗数配合力显著相关, 涉及第1、第2、第3、第6、第7、第8、第9、第10、第11和第12共10条染色体。12个标记位点杂合基因型中3个显示正向优势, 范围在15.1% (RM263-175/180)~ 21.1% (RM418-320/360)之间(表4)。

(续表3)
与亲本每穗总粒数配合力显著相关的标记位点鉴定出21个, 涉及第1、第2、第3、第5、第6、第7、第8、第10、第11和第12共10条染色体。21个标记位点杂合基因型中18个显示正向优势, 范围在13.6% (RM163-140/150)~23.7% (RM167-140/ 150)之间(表4)。鉴定出8个标记位点与亲本每穗实粒数配合力显著相关, 涉及第1、第2、第3、第6、第7和第11共6条染色体。8个标记位点杂合基因型中6个显示正向优势, 范围在12.5% (RM259-170/ 180)~24.2% (RM476B-140/150)(表4)。鉴定出15个标记位点与亲本结实率配合力显著相关, 涉及第1、第2、第3、第5、第6、第7、第8、第9和第10 共9条染色体。15个标记位点杂合基因型中2个显示正向优势, 优势值分别为13.4% (RM258-150/165) 和14.5% (RM406-145/160)(表4)。
与亲本千粒重配合力显著相关的标记位点鉴定出10个, 涉及第3、第6、第7、第8、第9和第10 共6条染色体。10个标记位点杂合基因型中9个显示正向优势, 范围在4.0% (RM5786-185/190)~7.5% (RM510-110/130)之间(表4)。鉴定出7个标记位点与亲本单株日产量配合力显著相关, 涉及第1、第2、第6和第7共4条染色体。7个标记位点杂合基因型中只有1个, 即RM406-145/160, 显示正向优势18.3% (表4)。

表4 盱眙环境下检测到的与亲本单个性状配合力相关的标记位点Table 4 Marker loci significantly related to combining ability for each trait in parents in Xuyi environment

(续表4)
2.3.3两个环境下都检测到的与亲本6个产量性状配合力显著相关的标记位点从表5可以看出, 两个环境下都检测到的影响亲本单株有效穗数、每穗总粒数、每穗实粒数、结实率、千粒重和单株日产量配合力的标记位点数量分别为4、11、5、3、5和1个。这29个标记位点涉及第1、第2、第3、第5、第6、第7、第8、第10、第11和第12共10条染色体。29个标记位点杂合基因型中, 20个显示正向优势, 9个显示负向优势。对于同一性状, 标记位点杂合基因型的优势方向, 在两个环境下是相同的, 而且优势值也很相
近。如RM345-180/190可使F1每穗总粒数优势增加22.0% (南京)和23.2% (盱眙); RM208-180/185可使F1每穗总粒数优势减少14.6% (南京)和15.3% (盱眙)。
两个环境下都检测到的标记位点中, 有3个各自与3个产量性状亲本配合力共相关, 分别是第2染色体上的RM406与结实率、千粒重和单株日产量配合力共相关, 标记位点杂合基因型RM406-145/160优势方向分别为正、负、正; 第3染色体上的RM16与单株有效穗数、每穗总粒数和千粒重配合力共相关, 标记位点杂合基因型RM16-180/190优势方向分别为负、正、负; 第6染色体上的RM340与单株有效穗数、每穗总粒数和每穗实粒数配合力共相关, RM340-110/160优势方向分别为负、正、正(表5)。另3个标记位点各自与2个产量性状亲本配合力共相关, 分别是第2染色体上的RM208与每穗总粒数和每穗实粒数配合力共相关, RM208-180/185优势方向均为负; 第7染色体上的RM8263与每穗总粒数和每穗实粒数配合力共相关, RM8263-190/195优势方向均为正; 第8染色体上的RM6863与单株有效穗数和千粒重配合力共相关, RM6863-185/195优势方向分别为负和正。其余标记位点只与1个产量性状配合力相关(表5)。

表5 两个环境下都检测到的与亲本性状配合力显著相关的标记位点Table 5 Marker loci related to combining ability for traits in parents detected across the two environments
3 讨论
配合力性状不同于一般的农艺性状, 它不能从试验材料本身直接观测到, 必须通过特定遗传设计的杂交获得的F1组合群体才能测定。配合力改良是杂交种育种的重要环节[19]。异花授粉作物的玉米自交系的配合力改良, 长期以来采用杂种优势亲本群间配组和轮回选择的方法[29-30]。水稻是自花授粉作物, 品种(品系)的一般配合力是在长期育种过程中自然形成的, 未曾刻意选择改良, 传统育种研究可以测定已然品种的配合力, 但却难以对它定向改良。近10年来, 陆续出现水稻杂种优势亲本群划分和利用分子标记技术改良亲本配合力的报道[19,31-32]。本课题组近年3项独立的杂交粳稻亲本产量性状和品质性状优异配合力的标记基因型鉴定研究[20-24]发现, 产量构成性状中每穗总粒数优异配合力的标记基因型数目最多, 单株有效穗数则相反, 不利配合力的标记基因型数目最多。这与科学研究和生产实际中观察到的杂交稻大穗少蘖出现频率很高的现象[6,33]相吻合。另外, 同一性状优异配合力标记基因型相同的数目较少。本研究南京点显示与梁奎等[20]报道的结果相同(相近)的配合力标记基因型共有8个。其中, RM340-110/160显示为单株有效穗数性状不利配合力的标记基因型; RM1211-170/180、RM208-180/185、RM340-110/160和RM345-180/190显示为每穗总粒数性状优异配合力标记基因型; 对每穗实粒数性状, RM341-170/195和RM340-110/160显示为优异配合力标记基因型, 而RM208-180/185显示为不利配合力的标记基因型。盱眙点结果与南京点结果相似。本研究两地点研究结果与刘二宝等[23]报道的结果相同的配合力标记基因型只有1个, 即与每穗总粒数、每穗实粒数配合力显著相关的RM208-180/185。但在两项研究中的效应相反, 本研究均为负值。推测以上3项研究结果异同的原因, 一是控制产量及其构成性状配合力的位点可能很多。位点间非等位基因之间的组合方式更多, 这为配合力改良提供了很大潜力。二是取材侧重点不同。梁奎等[20]的研究侧重于长江中下游的早熟晚粳类型组合; 本研究侧重于江淮稻区迟熟中粳类型组合。两项研究所用不育系有4个相同(863A、武羌A、18A和6427A), 恢复系有6个相同(宁恢8号、C堡、157TR-68、秀水04R、3726R-21、武育粳R-39), 共检测到8个相同的性状配合力标记基因型。刘二宝等[23]的研究材料来自“杂交水稻产业联盟中国杂交粳稻协作组”, 地理范围更广。本研究仅2个不育系(863A和18A)和1个恢复系(C4115)与之相同, 结果仅检测到1个与每穗总粒数、每穗实粒数配合力相关的标记基因型RM208-180/185。这反映了各性状能够检测到的优异标记基因型数目与亲本生态型有一定的关系, 分生态区鉴定杂交粳稻亲本性状优异配合力的标记基因型有其必要性。
本研究在两个环境下鉴定出BT-18A和武羌A是综合评价较优的不育系, C418是较优恢复系。BT-18A 是2013年审定的杂交粳稻新组合18优75的母本。18优75目前正在生产上推广应用。C418是多个杂交粳稻品种的父本, 配组的9优418、泗优418、3优18等杂交中粳品种在江淮地区累计推广面积达67万公顷以上。9优418已推广十多年, 目前仍是江淮稻区的主推杂交粳稻品种[1]。这说明田间配合力鉴定结果与生产应用状况具有较好的一致性。
检索数据库(http://agri-trait.dna.affrc.go.jp/)发现,两环境下都检测到的标记位点中, 有10个在其附近存在18个控制性状的基因/QTL。RM16附近存在控制每穗颖花数的spp3.1、qSNP3b和控制粒重的gw3a。RM340附近存在控制每穗颖花数的qSPN-6和控制每穗实粒数的gp6。RM406附近存在控制粒重的gw2.1和控制谷粒产量的yld2.1。RM208附近存在控制每穗颖花数的sn2.1和控制每穗实粒数的gn2.1。RM8263附近存在控制每穗颖花数的qSSP7和控制每穗实粒数的gp7a。RM3453附近存在控制每穗颖花数的qTNSP-1-1和sp1a。RM263附近存在控制每穗颖花数的sn2.1。RM570附近存在控制每穗颖花数的spp3.1和qSNP3b。RM152附近存在控制穗长的qPL8。RM259附近存在控制每穗实粒数的gp1。这一事实提供了通过分子标记辅助选择改良性状配合力的遗传基础。Huang等[34]通过SNP标记与杂种F1性状之间的全基因组关联分析, 揭示了正向显性稀有优异等位基因的集积对杂种优势有重要贡献。本研究还有一个与水稻杂交种实际表现相当一致的信息, 就是在上述18个控制产量相关性状的基因/QTL中, 15个是控制每穗总粒数和每穗实粒数的。这部分解释了为什么绝大多数杂交稻产量优势都表现在大穗性状上这一普遍存在的客观事实。Birchler[35]认为优良杂种表现的最佳配合力涉及来自整个基因组的诸多基因的协调效应。全基因组诸多基因如何协调产生这一普遍现象的遗传机制(代
谢通路), 有待进一步研究。
一个标记位点只与一个性状配合力相关的, 可直接根据标记位点杂合基因型信息进行辅助选择改良亲本该性状的配合力。如表5中RM247-170/180、RM3453-155/160、RM259-170/180、RM258-150/165、RM10-170/180信息分别可用于改良亲本单株有效穗数、每穗总粒数、每穗实粒数、结实率和千粒重性状的配合力。一个标记位点与亲本多个性状配合力相关时, 要考虑性状之间配合力效应的方向。方向一致时, 可同时改良多个性状的配合力。如表5 中RM8263-190/195可用于同时改良亲本每穗总粒数和每穗实粒数的配合力。举例说明改良策略, 若进一步提高杂交粳稻恢复系宁恢8号产量性状的配合力, 改良后的恢复系仍与863A配组, 则可以将C418或C堡中的RM8263-190 bp等位基因导入宁恢8号改良其每穗总粒数、每穗实粒数的配合力; 将3726R-21或武育粳R-39中的RM247-180 bp等位基因导入宁恢8号改良其单株有效穗的配合力; 将C657或K25中的RM10-170 bp等位基因导入宁恢8号改良其千粒重的配合力。这里除改良位点外其遗传背景保持不变, 再与863A配组的F1的产量水平就有可能高于原来组合86优8号。
4 结论
结合江淮稻区杂交粳稻骨干亲本产量性状的配合力和SSR分子标记基因型数据, 鉴定出一批影响产量性状配合力的标记位点; 一部分标记位点杂合基因型显示正向优势(优异配合力), 另一部分显示负向优势, 揭示了亲本性状配合力改良的遗传基础; 通过分子标记鉴定并聚合(或替换)更多的杂合基因型正向优势的标记等位基因到亲本中, 可作为杂交粳稻亲本(特别是恢复系)性状配合力改良的途径之一。
References
[1] 万建民. 中国水稻遗传育种与品种系谱. 北京: 中国农业出版社, 2010. pp 548–604
Wan J M. Genetics and Breeding, Cultivar Pedigree of Rice in China. Beijing: China Agriculture Press, 2010. pp 548–604 (in Chinese)
[2] Ma G H, Yuan L P. Hybrid rice achievements and development in China. In: Virmani S S, Mao C X, Hardy B, eds. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Protection. Los Baños (Philippines): International Rice Research Institute, 2003. pp 247–256
[3] 斯华敏, 刘文真, 付亚萍, 孙宗修, 胡国成. 我国两系杂交水稻发展的现状和建议. 中国水稻科学, 2011, 25: 544–552
Si H M, Liu W Z, Fu Y P, Sun Z X, Hu G C. Current situation and suggestions for development of two-line hybrid rice in China. Chin J Rice Sci, 2011, 25: 544–552 (in Chinese with English abstract)
[4] 邓华凤. 中国杂交粳稻. 北京: 中国农业出版社, 2008. pp 13–23
Deng H F. Japonica Hybrid Rice in China. Beijing: China Agriculture Press, 2008. pp 13–23 (in Chinese)
[5] 杜永林, 张巫军, 吴晓燃, 李刚华, 王绍华, 刘正辉, 唐设, 丁艳锋. 江苏省水稻产量时空变化特征. 南京农业大学学报,
2014, 37(5): 7–12. DOI: 10.7685/j.issn.1000-2030.2014.05.002 Du Y L, Zhang W J, Wu X R, Li G H, Wang S H, Liu Z H, Tang S, Ding Y F. The characteristics of spatial and temporal change of rice yield in Jiangsu Province. J Nanjing Agric Univ, 2014, 37(5): 7–12 (in Chinese with English abstract)
[6] 邓华凤, 何强, 舒服, 张武汉, 杨飞, 荆彦辉, 东丽, 谢辉. 中国杂交粳稻研究现状与对策. 杂交水稻, 2006, 21(1): 1–6
Deng H F, He Q, Shu F, Zhang W H, Yang F, Jing Y H, Dong L, Xie H. Status and technical strategy on development of japonica hybrid rice in China. Hybrid Rice, 2006, 21(1): 1–6 (in Chinese with English abstract)
[7] 汤述翥, 张宏根, 梁国华, 严长杰, 刘巧泉, 顾铭洪. 三系杂交粳稻发展缓慢的原因及对策. 杂交水稻, 2008, 23(1): 1–5
Tang S Z, Zhang H G, Liang G H, Yan C J, Liu Q Q, Gu M H. Reasons and countermeasures of slow development on three-line japonica hybrid rice. Hybrid Rice, 2008, 23(1): 1–5 (in Chinese with English abstract)
[8] 王才林. 江苏省杂交粳稻育种的现状、问题与对策. 西南农业学报, 2009, 24: 1165–1169
Wang C L. Current status and prospects of japonica hybrid rice breeding in Jiangsu, China. Southwest China J Agric Sci, 2009, 24: 1165–1169 (in Chinese with English abstract)
[9] 杨振玉. 我国杂交粳稻的发展及其技术策略. 杂交水稻, 2005, 20(2): 5–6
Yang Z Y. Development and technical strategy of japonica hybrid rice in China. Hybrid Rice, 2005, 20(2): 5–6 (in Chinese with English abstract)
[10] 洪德林, 汤玉庚. 粳稻雄性不育恢复基因研究: I. 粳稻雄性不育恢复基因的地理分布. 江苏农业学报, 1985, 1(4): 1–5
Hong D L, Tang Y G. Studies on fertility restoring genes for CMS Oryza sativa SUBSP. KENG in hybrid breeding: I. Geographical distribution of fertility restoring genes for CMS Oryza sativa SUBSP.KENG. J Agric Sci, 1985, 1(4): 1–5 (in Chinese with English abstract)
[11] 中国农业科学院, 湖南省农业科学院. 中国杂交水稻的发展.北京: 农业出版社, 1991. pp 28–40
Chinese Academy of Agricultural Sciences and Hunan Academy of Agricultural Sciences. Development of Hybrid Rice in China. Beijing: Agriculture Press, 1991. pp 28–40 (in Chinese)
[12] Gontcharova I K, Gontcharov S V. Hybrid rice breeding in Russia. In: Virmani S S, Mao C X, Hardy B, eds. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Protection. Los Baños (Philippines): International Rice Research Institute, 2003. pp 321–327
[13] Bastawisi A O, El-Mowafi H F, Abo Yousef M I, Draz A E, Aidy I R, Maximos M R, Badawi A T. Hybrid research and development in Egypt. In: Virmani S S, Mao C X, Hardy B, eds. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Pro-
tection. Los Baños (Philippines): International Rice Research Institute, 2003. pp 257–263
[14] 蔡健, 范海燕, 廖秋平, 马同富, 张桂权. 水稻恢复基因Rf3和Rf4聚合效应分析. 南京农业大学学报, 2014, 37(3): 20–26
Cai J, Fan H Y, Liao Q P, Ma T F, Zhang G Q. Analysis of pyramiding effect of the Rf3 and Rf4 genes on fertility restoration in rice with cytoplasmic male sterility. J Nanjing Agric Univ, 2014, 37(3): 20–26 (in Chinese with English abstract)
[15] 杨振玉, 张宗旭, 魏耀林, 赵迎春, 高勇. 粳型特异亲和恢复系C418的选育及其特性. 杂交水稻, 1998, 13(3): 31–32
Yang Z Y, Zhang Z X, Wei Y L, Zhao Y C, Gao Y. Breeding and characteristics of japonica type wide compatibility line C418. Hybrid Rice, 1998, 13(3): 31–32 (in Chinese with English abstract)
[16] Liu X C, Wu J L. SSR heterogenic patterns of parents for marking and predicting heterosis in rice breeding. Mol Breed, 1998, 4: 263–268
[17] Liu X C, Ishiki K, Wang W X. Identification of favorable AFLP markers to heterosis in hybrid rice. Breed Sci, 2002, 52: 201–206
[18] 刘小川, 王渭霞, 陈深广, 余柳青. 杂交稻米质性状的亲本配合力分子标记鉴定. 中国水稻科学, 2005, 19: 25–28
Liu X C, Wang W X, Chen G S, Yu L Q. Identification of molecular markers referred to the grain quality characters of rice hybrids based on the combining ability of their parental lines. Chin J Rice Sci, 2005, 19: 25–28 (in Chinese with English abstract)
[19] Liu X C, Chen S C, Chen J S, Ishiki K, Wang W X, Yu L Q. Improvement of combining ability for restorer lines with identified SSR marker in hybrid rice breeding. Breed Sci, 2004, 54: 341–346
[20] 梁奎, 黄殿成, 赵凯铭, 阮方松, 谢辉, 马文霞, 洪德林. 杂交粳稻亲本产量性状优异配合力的标记基因型筛选. 作物学报, 2010, 36: 1270−1279
Liang K, Huang D C, Zhao K M, Ruan F S, Xie H, Ma W X, Hong D L. Screening marker genotypes with elite combining ability for yield traits in parents of hybrid japonica rice (Oryza sativa L.). Acta Agron Sin, 2010, 36: 1270−1279 (in Chinese with English abstract)
[21] 黄殿成, 梁奎, 孙程, 江建华, 石明亮, 戴剑, 谢辉, 赵凯铭,阮方松, 马文霞, 洪德林. 杂交粳稻亲本米质性状优异配合力的标记基因型鉴定. 作物学报, 2011, 37: 405−414
Huang D C, Liang K, Sun C, Jiang J H, Shi M L, Dai J, Xie H, Zhao K M, Ruan F S, Ma W X, Hong D L. Identification of marker genotypes associated with elite combining ability for quality traits in parents of hybrid japonica rice (Oryza sativa L.). Acta Agron Sin, 2011, 37: 405−414 (in Chinese with English abstract)
[22] 黄殿成, 阮方松, 刘健, 谢辉, 赵凯铭, 梁奎, 江建华, 蔡勇林,洪德林. 杂交粳稻亲本穗长和枝梗数性状优异配合力的标记基因型筛选. 南京农业大学学报, 2011, 34(2): 1−7
Huang D C, Nguyen P T, Liu J, Xie H, Zhao K M, Liang K, Jiang J H, Cai Y L, Hong D L. Screening marker genotypes with elite combining ability for panicle length and branch numbers in parents of hybrid japonica rice (Oryza sativa L.). J Nanjing Agric Univ, 2011, 34(2): 1−7 (in Chinese with English abstract)
[23] 刘二宝, 刘洋, 刘晓丽, 刘强明, 赵凯铭, Edzesi W M, 洪德林.杂交粳稻亲本产量性状优异配合力的标记基因型检测. 中国水稻科学, 2013, 27: 473−481
Liu E B, Liu Y, Liu X L, Liu Q M, Zhao K M, Edzesi W M, Hong D L. Detecting marker genotypes with elite combining ability for yield traits in parents of hybrid japonica rice. Chin J Rice Sci, 2013, 27: 473−481 (in Chinese with English abstract)
[24] 刘洋, 刘二宝, 曾思远, 刘强明, 普伟, 梁利君, 刘佩睿, 张琪梦, 洪德林. 杂交粳稻亲本米质性状配合力的分子标记区段检测. 中国水稻科学, 2015, 29: 443−451
Liu Y, Liu E B, Zeng S Y, Liu Q M, Pu W, Liang L J, Liu P R, Zhang Q M, Hong D L. Identification of molecular marker fragments associated with combining ability for quality traits in parents of hybrid japonica rice (Oryza sativa L.). Chin J Rice Sci, 2015, 29: 443−451 (in Chinese with English abstract)
[25] 程宝山, 万志兵, 洪德林. 35个粳稻品种SSR指纹图谱的构建及遗传相似性分析. 南京农业大学学报, 2007, 30(3): 1–8
Cheng B S, Wan Z B, Hong D L. Establishment of SSR finger print map and analysis of genetic similarity among 35 varieties in japonica rice (Oryza sativa L.). J Nanjing Agric Univ, 2007, 30(3): 1−8 (in Chinese with English abstract)
[26] 莫惠栋. p×q交配模式的配合力分析. 江苏农学院学报, 1982, 3(3): 51–57
Mo H D. The analysis of combining ability in p×q mating pattern (continued). J Jiangsu Agric Coll, 1982, 3(3): 51–57 (in Chinese)
[27] 莫惠栋. p×q交配模式的配合力分析(续). 江苏农学院学报, 1982, 3(4): 53–57
Mo H D. The analysis of combining ability in p×q mating pattern (continued). J Jiangsu Agric Coll, 1982, 3(4): 53–57 (in Chinese)
[28] 徐辰武. 稻米品质性状的遗传研究. 南京农业大学博士学位论文, 江苏南京, 1998
Xu C W. Inheritance of Quality Characters of Rice Grain. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 1998 (in Chinese with English abstract)
[29] Russell W A. Genetic improvement of maize yield. Adv Agron, 1991, 46: 245–298
[30] Dudly J W, Saghai M A, Rufener G K. Molecular marker and grouping of parents in maize breeding programs. Crop Sci, 1991, 31: 718–723
[31] 王胜军, 万建民, 陆作楣. 利用SSR分子标记划分杂交籼稻亲本群的研究. 作物学报, 2006, 32: 1437–1443
Wang S J, Wan J M, Lu Z M. Parental cluster analysis in indica hybrid rice (Oryza sativa L.) by SSR analysis. Acta Agron Sin, 2006, 32: 1437–1443 (in English with Chinese abstract)
[32] 王胜军, 陆作楣. 我国常用杂交籼稻亲本杂种优势群的初步研究. 南京农业大学学报, 2007, 30(1): 14–18
Wang S J, Lu Z M. Study on heterosis groups of key parental lines in indica hybrid rice (Oryza sativa L.) in China. J Nanjing Agric Univ, 2007, 30(1): 14–18 (in Chinese with English abstract)
[33] Virmani S S, Aquino R C, Khush G S. Heterosis breeding in rice (Oryza sativa L.). Theor Appl Genet, 1982, 63: 373–380
[34] Huang X H, Yang S H, Gong J Y, Zhao Y, Feng Q, Gong H, Li W J, Zhan Q L, Cheng B Y, Xia J H, Chen N, Hao Z N, Liu K Y, Zhu C R, Huang T, Zhao Q, Zhang L, Fan D L, Zhou C C, Lu Y Q, Weng Q J, Wang Z X, Li J Y, Han, B. Genomic analysis of hybrid rice varieties reveals numerous superior alleles that contribute to heterosis. Nat Commun, 2015, 6: 6258, DOI: 10.1038/ ncomms7258
[35] Birchler J A. The genetic basis of hybrid vigour. Nat Plants, 2015, 1: 15020, DOI: 10.1038/nplants.2015.20
URL: http://www.cnki.net/kcms/detail/11.1809.S.20151218.0915.004.html
Identifying SSR Marker Locus Genotypes with Elite Combining Ability for Yield Traits in Backbone Parents of Japonica Hybrid Rice (Oryza sativa L.) in Jianghuai Area
XIE Hui1,2, DANG Xiao-Jing1, LIU Er-Bao1, ZENG Si-Yuan1, and HONG De-Lin1,*1State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China;2China National Japonica Rice Research and Development Center, Tianjin 300457, China
Abstract:Identifying marker loci related to combining ability (CA) for yield trait in parents of japonica hybrid rice facilitates improving CA of parents and enhancing standard heterosis degree of japonica rice by using molecular marker-assisted selection techniques. F1seeds of 90 combinations were made by hand-crossed nine CMS lines with ten restorer lines using North Carolina Design II. The F1populations were planted in Nanjing and Xuyi environments, and six yield traits were investigated. CA of the 19 parental lines was analyzed for six yield traits respectively using the data of 90 F1’s. Combining the data of CA and SSR marker genotypes of the 19 parental lines, SSR marker loci related to CA for six yield traits were detected. Results showed that BT-18A and Wuqiang A were elite CMS lines, and C418 was elite restorer lines in both environments. Number of detected SSR marker loci related to CA for effective panicles per plant, spikelets per panicle, filled grains per panicle, seed setting rate, 1000-grain weight and daily yield per plant were 8, 13, 11, 6, 6, and 2, respectively in Nanjing, 12, 21, 8, 15, 10, and 7, respectively in Xuyi, and 4, 11, 5, 3, 5, and 1, respectively in both environments. Heterozygous genotype marker loci showing positive heterosis accounted for 74% (34/46) in Nanjing, and 53% (39/73) in Xuyi. Among the SSR marker loci detected in both environments, three
were each co-associated with CA for three yield traits, and another three for two yield traits. The remaining 14 marker loci were each associated with CA for one yield trait. Through data-base searching, genes/QTLs for the corresponding traits were found nearly ten of the marker loci detected in both environments. Strategies of enhancing CA for yield traits of restorer lines in japonica rice using the marker loci identified in this study were discussed.
Keywords:Hybrid Japonica rice; Backbone parents; Combining ability for yield traits; Marker locus genotypes
收稿日期Received(): 2015-07-29; Accepted(接受日期): 2015-11-20; Published online(网络出版日期): 2015-12-18.
通讯作者*(Corresponding author): 洪德林, E-mail: delinhong@njau.edu.cn, Tel: 025-84396626
DOI:10.3724/SP.J.1006.2016.00330
