
汉中地区不同黑稻品种查尔酮合成酶基因的遗传变异分析
2016-03-01 12:57李蕊清田蕾铭路宏朝张力瑞
贵州农业科学 2016年3期
李蕊清, 刘 欢, 田蕾铭, 路宏朝, 张力瑞, 张 辉, 张 涛*
(1.陕西理工学院 生物科学与工程学院, 陕西 汉中 723001; 2.陕西省南郑中学, 陕西 南郑 723100)
汉中地区不同黑稻品种查尔酮合成酶基因的遗传变异分析
李蕊清1, 刘 欢1, 田蕾铭1, 路宏朝1, 张力瑞2, 张 辉2, 张 涛1*
(1.陕西理工学院 生物科学与工程学院, 陕西 汉中 723001; 2.陕西省南郑中学, 陕西 南郑 723100)
为汉中黑稻起源及其花青苷合成机制提供分子遗传依据,通过分析汉中地区8个黑稻品种和其他植物查尔酮合成酶(chalcone synthase,CHS)基因序列,利用PCR扩增8个黑稻品种查尔酮合成酶基因并测序,使用DNAMAN和MEGA5.0软件分别对水稻的CHS基因外显子序列进行比对和不同物种间的聚类。结果表明:水稻种内存在4个多态位点,8种黑稻中存在3个多态位点,比对CHS氨基酸序列,仅黑宝品种存在1个氨基酸突变。经聚类分析,8种黑稻亲缘关系最近,与籼稻聚为一类,然后与粳稻聚在一起,禾本科植物CHS同源性很高。结论:CHS是一个古老的基因,可作为种属鉴定的参考基因,CHS虽是花青苷合成的限速酶基因,但其序列的变异可能与黑稻花青苷的合成无相关性。
黑稻; 查尔酮合酶; 序列分析; 遗传变异
水稻是稻属分支下栽培驯化最悠久的一个种,分为籼稻和粳稻2个变种,其中存在一些古老而珍贵的种质资源——黑稻,属于特种稻范畴。黑稻种皮花青苷是天然的色素,无毒性,既可作为食品着色剂,又具有提高机体非特异性免疫功能,增强抗病能力及抗过敏、抗氧化等多种保健功能[1-4]。花青苷类化合物种类多样,不同品种黑米花青苷化学结构存在一定差异,表明花青苷的合成途径相对较复杂[5-6],其合成的分子机理及相关功能基因的鉴定研究可为黑稻种质培育和遗传进化研究提供新的思路。
植物花青素生物合成途径可分为3个阶段:1) 苯丙氨酸经过一系列反应生成4-香豆酰CoA,此为多数植物次生代谢途径共有;2) 4-香豆酰CoA和丙二酰CoA反应生成二氢黄酮醇,此过程由查尔酮合酶(Chalcone synthase,CHS)、查尔酮异构酶(CHI)和黄烷酮3-羟化酶(F3H)等催化;3) 二氢黄酮醇通过二氢黄酮醇还原酶(DFR)将二氢黄酮醇转化成无色花青素,然后在花青素合成酶(ANS)的作用下经过氧化脱水形成花青素[7]。花青素很不稳定,需经糖基化、甲基化、酰基化和羟基化等一系列修饰作用才能形成稳定的花青苷。
在花青苷合成途径中,查尔酮合成酶是一个关键酶[8],研究其基因序列变异、表达调控和蛋白质结构等,有助于解析植物花青素积累的分子机理。因此,很多学者对不同植物的查尔酮合酶基因进行了大量研究,确定了植物CHS有2个外显子,1个内含子,外显子2编码几乎所有功能位点[9-12],但关于黑稻查尔酮合成酶基因完整的编码区序列变异分析尚未见详细报道。笔者选取陕西汉中地区主要种植的8个黑稻品种,通过对其查尔酮合成酶基因编码区的测序和比对,CHS的变异与黑稻花青苷合成的关系,并基于CHS基因序列进行了黑稻与其他种属植物的系统树构建,以期日后为黑稻的起源与遗传进化等科学问题提供理论基础。
1 材料与方法
1.1 材料
选取汉中地区种植的8个黑稻品种为材料,由陕西省水稻研究所提供,该8个品种全部为籼稻,分别为黑宝(HB),黑优粘(HYZ),黑丰(HF),黑帅(HS),黑米A(HMA),培811(P811),黑米B(HMB),云黑(YH)。
1.2 DNA提取
稻种在调温调湿箱中萌发,取水稻其嫩叶叶尖,按照改良的CTAB法提取基因组DNA[14]。
1.3 基因及引物序列信息
根据GenBank检索CHS基因序列(登陆号X89859)(表1),Primer 5.0软件设计PCR的引物(表2),由上海桑尼生物科技有限公司合成。

表1 不同植物CHS基因序列GeneBank登陆号

表2 CHS序列的PCR产物及相关信息
1.4 PCR扩增反应体系及程序
PCR反应总体系为25 μL:10×PCR缓冲液(含15 mmol/L Mg2+)2.5 μL,dNTP(100 mmol/L)0.5 μL,上下游引物(10 pmol)各1 μL,Taq DNA聚合酶(0.5U/μL)2.0 μL,DNA模板(50 ng/μL)1.0 μL,超纯水17.0 μL。PCR扩增条件:94℃预变性4 min;94℃变性45 s,52℃退火30 s,72℃延伸1 min,35个循环,然后72℃延伸10 min,4℃保存。
1.5 数据处理
采用使用DNAMAN和MEGA5.0软件对CHS基因序列进行比对和聚类分析,通过www.expasy.org网络软件对查尔酮合成酶的理化性质进行预测分析。
2 结果与分析
2.1 黑稻CHS基因的mRNA序列比对
黑稻CHS基因的mRNA序列共有2个外显子,外显子I 187 bp,外显子II 1 010 bp,共1 197 bp,编码398个氨基酸。
将获得的8个品种的CDS序列与NCBI公布的水稻CHS基因序列(No.X89859和No.AB058397)比对分析发现,汉中地区8个黑稻栽培品种CHS的CDS序列长度与GeneBank的序列一致,不存在插入突变和缺失突变。8个黑稻栽培籼稻品种,通过与NCBI公布的籼稻和粳稻的序列进行比对,汉中8个品种黑稻内存在3个多态位点,与公布的序列比较则存在4个多态位点(图1)。这4个位点的变异为编码区57位的A/G转换,473位的A/G转换,530位的C/G颠换(粳稻与籼稻之间),789位的G/A转换(图2)。
2.2 不同物种基于CHS的mRNA序列的聚类树
从图3看出,2种聚类结果基本一致,汉中8个黑稻品种与籼稻先聚为一类,亲缘关系较近,这与实际相吻合,然后与粳稻聚为一个稻属的大类。稻属植物与玉米、小麦等禾本科植物聚在一个分支。
2.3 不同植物在CHS基因位点的同源性和遗传距离
由表3可见,黑优粘与黑丰同源性为100%,黑米A与培811同源性为100%,黑帅、云黑与黑米B完全同源,黑米A与培811和粳稻的同源性为99.9%。8个品种与粳稻的同源性在99.7%以上,8个品种中有3个与籼稻同源性在99.7%,2个与籼稻同源性在99.8%,2个与籼稻同源性为99.9%,黑宝与籼稻的同源性在99.6%,低于与粳稻的99.7%。水稻CHS位点的序列同源性非常保守,均在99.6%以上。水稻与小麦的遗传距离为0.172~0.175,和玉米遗传距离为0.151~0.152,水稻与玉米的亲缘关系较小麦近一些。水稻与禾本科以外其他植物的同源性在73.1%~77.5%。说明,查尔酮合成酶基因是一个相对保守的基因,是所有植物生长发育必需的一个基因。

图1 籼稻、粳稻与8种黑稻的CHS基因外显子序列
Fig.1 The exon sequence alignment of indica, japonica and eight black rice varieties
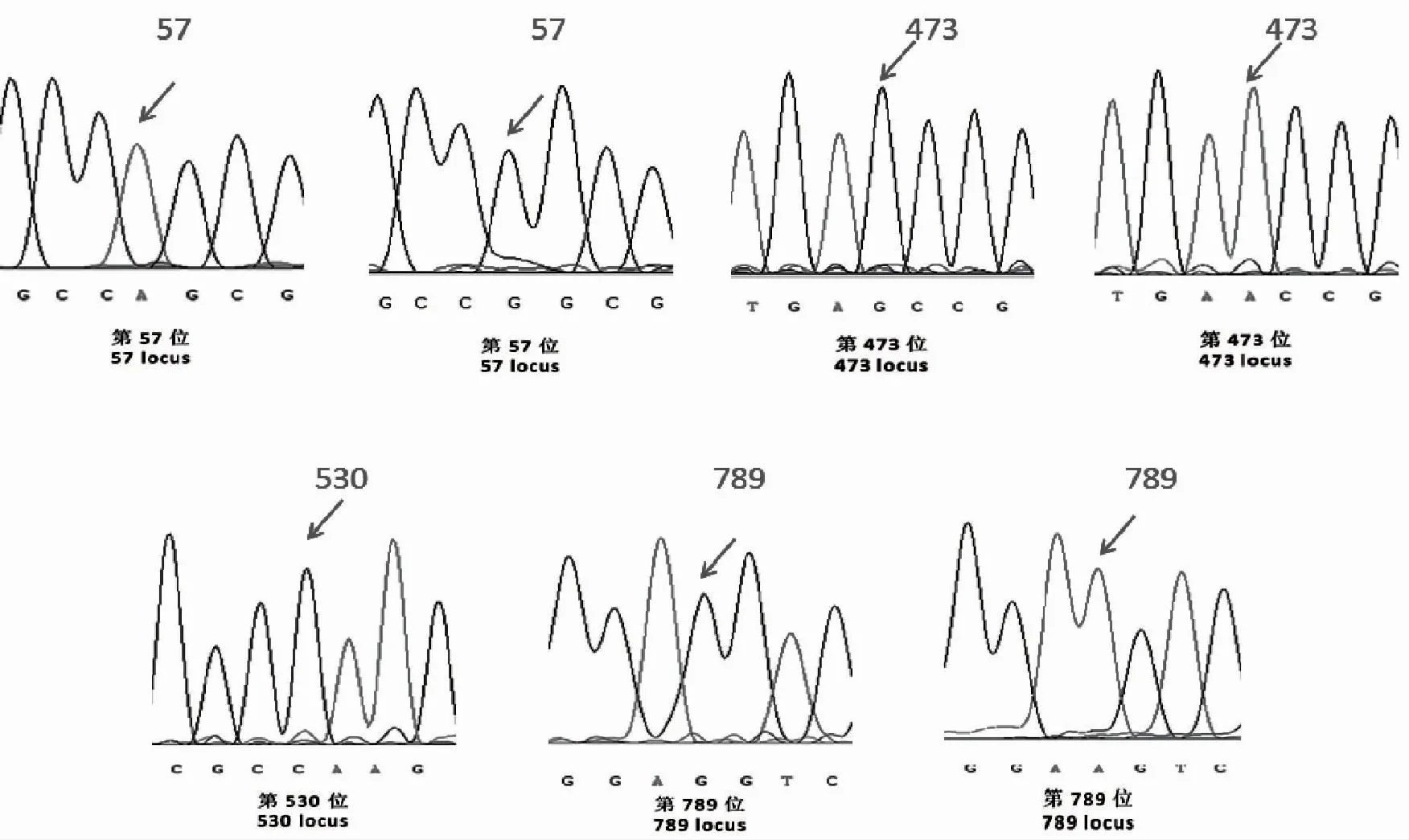
图2 8种黑稻的CHS基因外显子区SNP位点
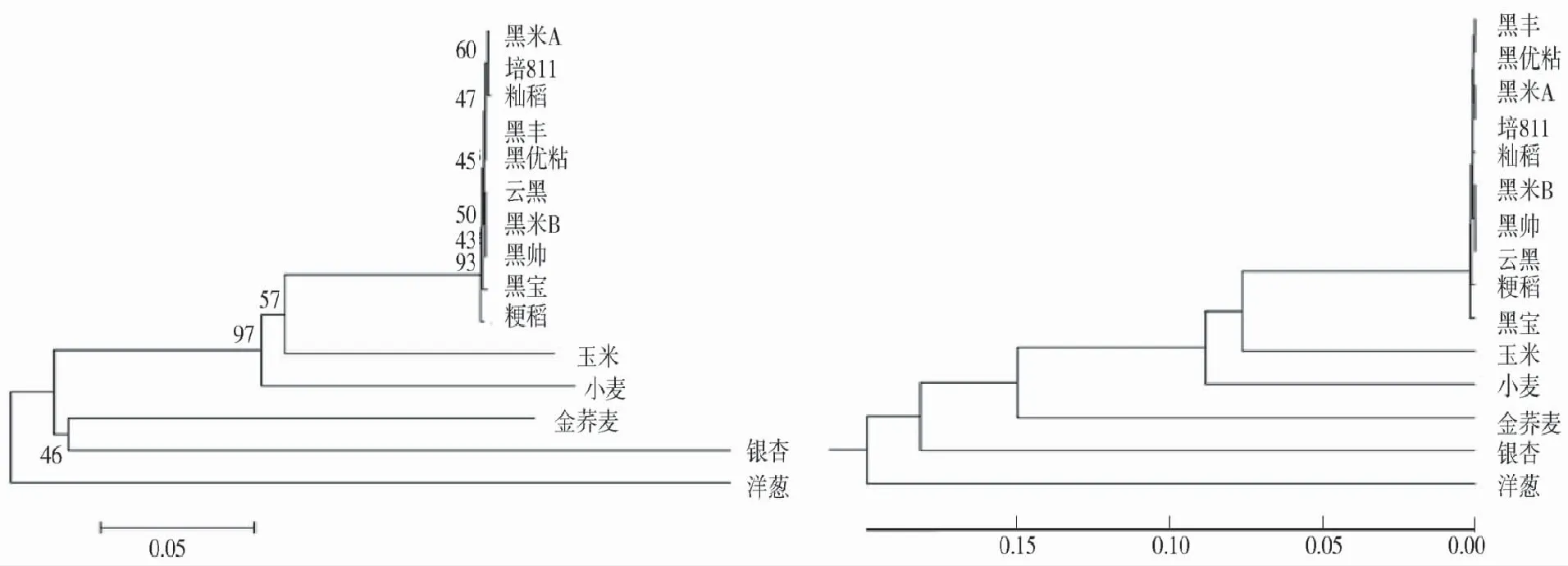
图3 基于CHS基因序列的聚类分析
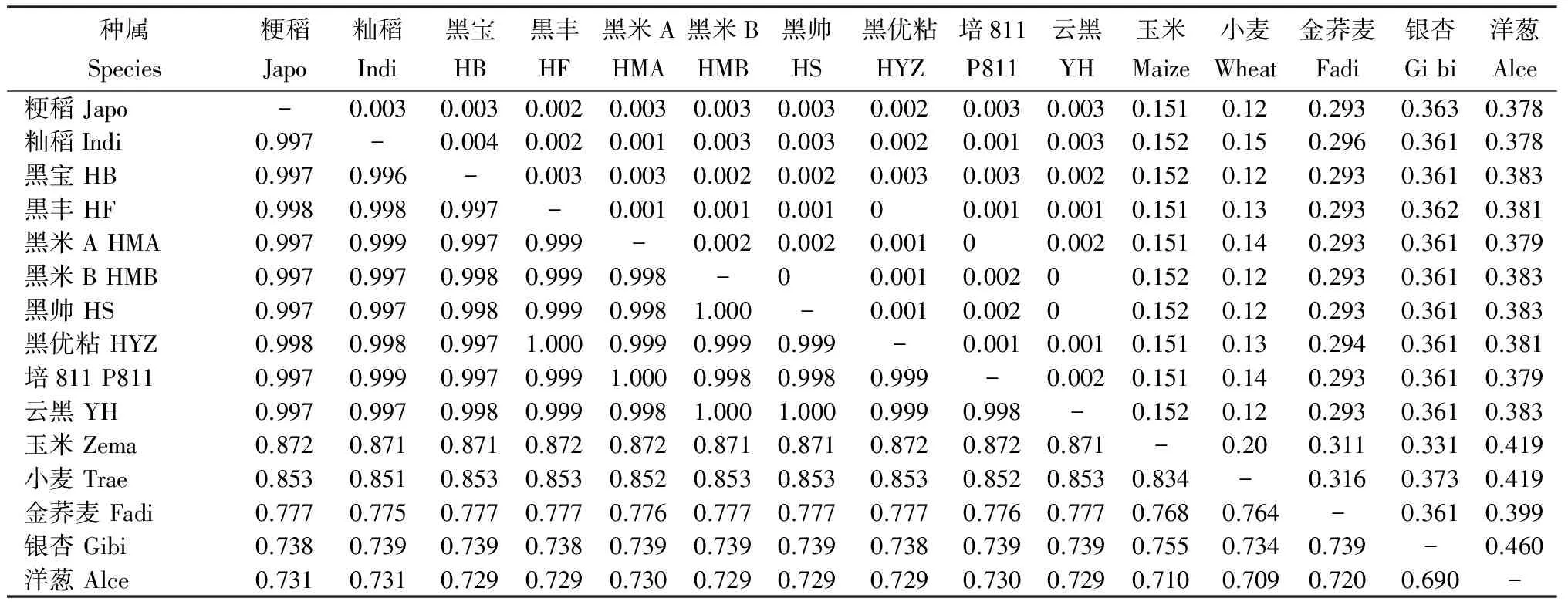
种属Species粳稻Japo籼稻Indi黑宝HB黒丰HF黑米AHMA黑米BHMB黑帅HS黑优粘HYZ培811P811云黑YH玉米Maize小麦Wheat金荞麦Fadi银杏Gibi洋葱Alce粳稻Japo-0.0030.0030.0020.0030.0030.0030.0020.0030.0030.1510.120.2930.3630.378籼稻Indi0.997-0.0040.0020.0010.0030.0030.0020.0010.0030.1520.150.2960.3610.378黑宝HB0.9970.996-0.0030.0030.0020.0020.0030.0030.0020.1520.120.2930.3610.383黒丰HF0.9980.9980.997-0.0010.0010.00100.0010.0010.1510.130.2930.3620.381黑米AHMA0.9970.9990.9970.999-0.0020.0020.00100.0020.1510.140.2930.3610.379黑米BHMB0.9970.9970.9980.9990.998-00.0010.00200.1520.120.2930.3610.383黑帅HS0.9970.9970.9980.9990.9981.000-0.0010.00200.1520.120.2930.3610.383黑优粘HYZ0.9980.9980.9971.0000.9990.9990.999-0.0010.0010.1510.130.2940.3610.381培811P8110.9970.9990.9970.9991.0000.9980.9980.999-0.0020.1510.140.2930.3610.379云黑YH0.9970.9970.9980.9990.9981.0001.0000.9990.998-0.1520.120.2930.3610.383玉米Zema0.8720.8710.8710.8720.8720.8710.8710.8720.8720.871-0.200.3110.3310.419小麦Trae0.8530.8510.8530.8530.8520.8530.8530.8530.8520.8530.834-0.3160.3730.419金荞麦Fadi0.7770.7750.7770.7770.7760.7770.7770.7770.7760.7770.7680.764-0.3610.399银杏Gibi0.7380.7390.7390.7380.7390.7390.7390.7380.7390.7390.7550.7340.739-0.460洋葱Alce0.7310.7310.7290.7290.7300.7290.7290.7290.7300.7290.7100.7090.7200.690-
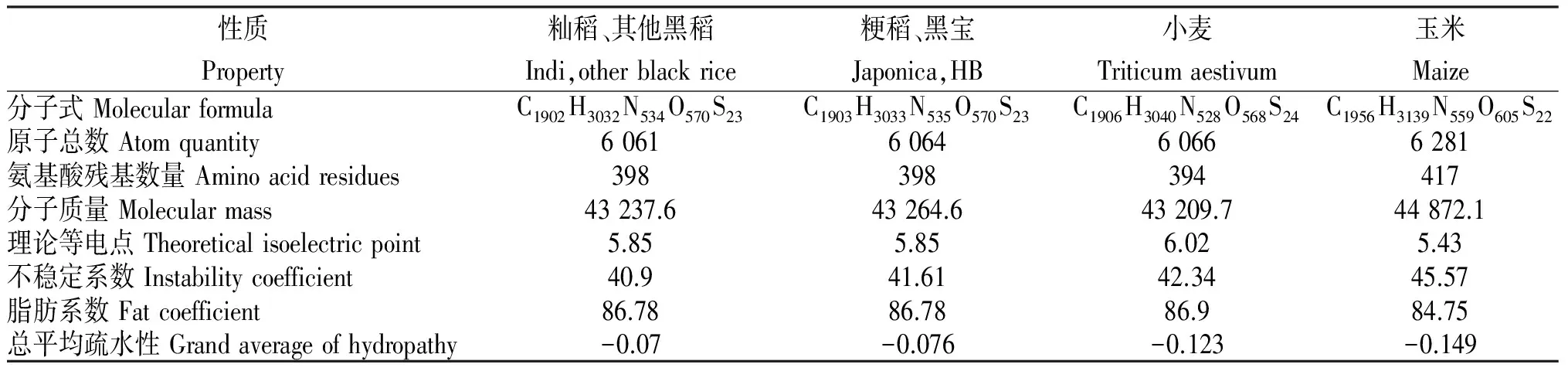
表4 不同植物查尔酮合成酶的理化性质
2.4 不同黑稻品种CHS基因的遗传变异性
稻属植物查尔酮合成酶有398个氨基酸,通过比较此8个品种与其他水稻查尔酮合成酶基因发现,CDS区存在的4个变异中只有473位的A/G转换导致第158个氨基酸组成的改变,粳稻和品种黑宝此处为Asn,籼稻、黑优粘、黑丰、黑帅、黑米A、培811、黑米B和云黑为Ser。
从表4可知,158位氨基酸变化导致查尔酮合成酶的疏水性有改变,籼稻为-0.076,粳稻为-0.070,在稳定性方面,籼稻和其他7个品种黑稻要高于粳稻和黑宝,其他理化性质无影响。与小麦和玉米相比,其疏水性均低于小麦的-0.123和玉米的-0.149,水稻较为稳定,推测其查尔酮合酶在植物体内作用时间相对较长。
3 结论与讨论
汉中地处秦巴山区腹地,位于汉江中上游,是陕西水稻的主产区,特种稻资源丰富,产五色稻,以黑稻最为著名,种质资源丰富[15]。汉中地区黑稻资源的研究主要集中在资源库建立、常规育种、种植技术和黑稻花青苷的综合开发利用等方面[16-17],也有学者[18]进行汉中水稻DNA遗传多样性及黑稻蛋白质标记研究[19]。黄酮类化合物是一类天然次生代谢物,广泛存在于各种陆生植物当中,在植物器官花色素积累、抗胁迫、抗菌和细胞的发育与分化等过程中有着重要的作用[20]。CHS是黄酮类物质合成途径中的关键酶,其突变、沉默或过表达等可能影响多种黄酮类物质合成,目前CHS基因的研究很多,但大都集中在对豆科植物异黄酮合成的研究[21-22]。如许锋等[23]研究认为,银杏CHS基因的表达水平、酶的活性水平、类黄酮的含量三者之间有良好的正相关关系。CHS在植物颜色形成过程中也起着重要作用,尤其是在颜色鲜艳的观赏植物中,其研究重在改良花色,提高植物的观赏价值[24],CHS的特异性表达与花色相关,其表达量可引起花色的变化[25]。由于植物体内黄酮物质的形成与色素的沉积是植物形成具有的一个基本特征,故CHS基因作为一个相对古老的基因,基因结构和序列相对保守,但在漫长的历史进化中,由于基因的变异与物种分化导致不同物种的CHS有较大差异。
黑稻、黑麦和黑玉米等黑色有机农作物因其独特的品质较好的保健功能受到了消费者青睐,市场经济价值很大,故CHS的表达量对籽粒颜色的影响正成为禾本科植物分子遗传学和育种研究的热点。由于CHS基因是一个较为保守的古老基因,尽管不同群体之间存在遗传变异,但不影响植物生长发育,能否以该位点作为研究水稻起源进化与分化研究的参考点,也是本研究关注的问题。因此,本研究设计以CHS基因为目标来分析黑稻花青苷合成与该基因遗传变异的关系,为黑稻育种实践中相关代谢途径关键酶的基因修饰研究奠定基础。另外,欲通过分析汉中黑稻CHS这一古老基因的遗传多态性,探索黑稻与普通稻在遗传分化上的差异,为汉中丰富的黑稻资源保护和开发提供一些分子遗传数据。
本研究以NCBI公布的籼稻和粳稻CHS基因序列为参考发现,在汉中8个黑稻栽培品种CHS基因编码区非常保守,品种内存在4个多态位点。以CHS基因序列为基础进行聚类分析判定8个品种的均为籼稻,与实际相符合,禾本科之间亲缘关系很近,稻属植物之间同源性极高,表明查尔酮合成酶基因作为不同属间的亲缘关系鉴定完全可行,用于种内亚种间的亲缘关系分析和起源进化研究也是可行的,但对于种内品种间的亲缘关系分析和品种鉴定不可靠,只可作为参考,并需扩大样本数量系统分析。在8个品种中7个查尔酮合成酶氨基酸序列相同,仅有黑宝品种存在1个氨基酸的变异,该品种CHS氨基酸序列与粳稻相同,可能暗示黑宝品种与粳稻在选育过程中存在一定的亲缘关系。通过将汉中8个黑稻CHS基因编码区序列与普通的籼稻与粳稻的基因序列比对分析认为,水稻CHS基因高度保守,其少量变异不影响查尔酮合成酶的功能,序列变异与花青苷合成无相关性。但其表达量的高低、环境效应和其他因素对黑稻花青苷的积累是否存在影响还需进一步研究。通过本研究可确定以CHS基因序列为基础进行种间和亚种之间的亲缘关系鉴定是可行的,但不可作为品种间鉴定分析。研究结果表明,查尔酮合成酶基因是一个较为保守的古老基因,虽然不同品种之间存在一定差异,但无插入和缺失突变,不会造成移码变异,造成该基因功能的明显变化,从而影响植物正常生长发育。
[1] 侯夫云,王庆美,李爱贤,等.植物花青素合成酶的研究进展[J].中国农学通报,2009,25(21):188-190.
[2] 陈起萱,凌文华.黑米和红米对兔主动脉脂质斑块面积和血脂的影响[J].卫生研究,2000,29(3):170-172.
[3] 谢黎虹,罗玉坤,陈 能.红米和黑米的营养功效研究进展[J].西部粮油科技,2003,28(6):35-37.
[4] 孙 玲,张名位,池建伟,等.黑米的抗氧化性及其与黄酮和种皮色素的关系[J].营养学报,2000,22(3):246-249.
[5] 李新生,吴升华.陕西黑稻资源及其开发利用[J].资源科学,1998,20(6):67-73.
[6] 陈 新.银杏苯丙氨酸解氨酶(PAL)的基因克隆和生物信息学分析[D].成都:成都中医药大学,2003.
[7] Ma Tingrui, Zhang Jinwen, Liang Huiguang, et al. Plant Anthocyanin Synthesis and Gene Regulation[J].Agricultural Science & Technology,2012,3:507-511,540.
[8] 杨会娜,田新惠,李艳军,等.天然棕色棉查尔酮合成酶基因(GhCHS1)的克隆和实时定量表达分析[J].棉花学报,2010(1):42-48.
[9] 王金玲,瞿礼嘉,陈 军,等.CHS基因外显子2的进化规律及其用于植物分子系统学研究的可行性[J].科学通报,2000(9):942-950.
[10] 蒙 华,李成磊,吴琦等.金荞麦查尔酮合成酶基因CHS的克隆及序列分析[J].草业学报,2010(3):162-169.
[11] 张明姝.菊花查尔酮合成酶基因的克隆与序列分析[D].北京:北京林业大学,2006.
[12] 钟德馨,方袁梦梦,郭壮浩,等.决明查尔酮合成酶全长基因序列的克隆与分析[J].生物技术通报,2013(5):99-104.
[13] Ursula Niesbach-Kl sgen, Ellen Barzen, Jürgen Bernhardt, et al. Chalcone synthase genes in plants: A tool to study evolutionary relationships [J] Journal of Molecular Evolution,1987,26:213-1125.
[14] Zhang Y, Bog R. Evluation of authentication of methodology on genonmic DNA isolation of plant[J].Drag Evaluation,2004,4:292-297.
[15] 王胜宝,冯志峰,李新生,等.陕西汉中特种稻产业发展现状及前景[J].新疆农业科学,2010(S2):147-150.
[16] 李新生,邓文辉,吴三桥,等.陕西三种特种稻米氨基酸及品质分析[J].氨基酸和生物资源,2001(4):1-3.
[17] 曹小勇,李新生.黑米花色素苷类色素研究现状及展望[J].氨基酸和生物资源,2002(1):3-6.
[18] 张 羽,李新生,冯志峰,等.陕西省有色稻资源的SSR多态性分析[J].植物遗传资源学报,2011(5):828-832.
[19] 丁 锐.黑稻主要栽培品种过氧化物酶同工酶的研究[J].安徽农业科学,2005(6):947-948.
[20] 朱见明,李 娜,张亚军,等.苜蓿黄酮的研究进展[J].草业科学,2009(9):156-162.
[21] 单丽伟,汪 勇,王美玲,等.大豆类黄酮生物合成关键酶CHS基因的克隆及表达分析[J].西北植物学报,2012(11):2164-2168.
[22] 牛天敏,马会勤,陈尚武.大豆查尔酮合成酶(CHS)基因的克隆、表达及其在雪莲提取液中的代谢产物分析[J].中国生物工程杂志,2007(2):58-63.
[23] 许 锋,程水源,王 燕,等.TAIL-PCR方法快速克隆银杏查尔酮合成酶基因及序列分析(英文)[J].果树学报,2007(2):237-243.
[24] 赵叔灵,郭传明,陈俊愉.植物花色呈现的生物化学、分子生物学机制及其基因工程改良[J].西北植物学报,2003,23(6):1024-1035.
[25] 杨 丽,刘雅莉,王跃进.百合查尔酮合成酶(chs)基因的cDNA克隆与分析[J].河南农业科学,2011,40(1):124-126.
(责任编辑: 刘忠丽)
Genetic Variation of Chalcone Synthase Gene of Black Rice Planted in Hanzhong
LI Ruiqing1, LIU Huan1, TIAN Leiming1, LU Hongzhao1, ZHANG Lirui2, ZHANG Hui2, ZHANG Tao1*
(1.SchoolofBiologicalScienceTechnologyandEngineering,ShaanxiUniversityofTechnology,Hanzhong,Shaanxi723000;2.NanzhengHighSchool,Nanzheng,Shaanxi723100,China)
In order to provide a scientific basis for the origin and the synthesis mechanism of anthocyanin, the differences of chalcone synthase(CHS) gene sequenc were analyzed amongO.sativaand different plants in this study. PCR amplify theCHSsequence of eight kinds of black rice in Hanzhong and sequencing, comparing the CHS mRNA sequence amongO.sativaby DNAMAN and clustering between different species by MEGA5.0. Results: Three genetic variation in Hanzhong black rice was discovered and four genetic variation inO.sativawas found. Comparing the CHS amino acid sequence, one amino acid variation in HB (a kind of black rice in Hanzhong) was found. The genetic relationship rice was the closest, and the eight rice varieties were.The homology of CHS was very high in gramineae. Conclusion:CHSis an ancient gene, that can be as a reference gene for species identification.CHSis the rate-limiting enzyme, but mutation of CHS sequence may be not related to anthocyanin synthesis in rice.
black rice; chalcone synthase; sequence analysis; genetic variation
2015-07-22; 2016-02-24修回
陕西省科技厅农业攻关项目“植物源杀螨剂研究与产品开发”(2013K02-26-02);陕西省“春笋计划”2014-2015年度课题研究项目
李蕊清(1994-),女,在读本科,专业方向:生物科学。E-mail:liruiqing248@163.com
*通讯作者:张 涛(1978-),男,硕士,副教授,从事分子遗传学研究。E-mail:zl780823@163.com
1001-3601(2016)03-0104-0018-05
S511.2+1
A
猜你喜欢
古今农业(2022年2期)2022-08-15
安徽农学通报(2022年8期)2022-05-06
天然产物研究与开发(2018年9期)2018-10-08
江西农业(2018年9期)2018-02-11
天然产物研究与开发(2016年1期)2016-06-05
小猕猴学习画刊(2016年9期)2016-05-14
小主人报(2015年2期)2015-09-15
小主人报(2015年1期)2015-03-11
世界热带农业信息(2014年3期)2014-08-12
小雪花·成长指南(2014年1期)2014-03-04
