
广西来宾蓬莱滩剖面瓜德鲁普统—乐平统(G-L)界线生境型及其意义
2016-02-24 08:41:15韦雪梅江增光白玛曲宗韦恒叶
东华理工大学学报(自然科学版) 2016年4期
韦雪梅, 江增光, 白玛曲宗, 韦恒叶
(东华理工大学地球科学学院,江西 南昌 330013)
广西来宾蓬莱滩剖面瓜德鲁普统—乐平统(G-L)界线生境型及其意义
韦雪梅, 江增光, 白玛曲宗, 韦恒叶
(东华理工大学地球科学学院,江西 南昌 330013)
对广西来宾地区蓬莱滩剖面瓜德鲁普统茅口组(P2m)至乐平统合山组(P3h)进行了详细的生境型分析,共识别出4个大类与5个亚类生境型,包含Ⅱ2型(潮间带与正常浪基面之间),Ⅲ1型(上部浅海上部),Ⅲ2型(上部浅海下部)、Ⅳ型(下部浅海)和Ⅴ型(半深海)。 生境型总体上反映海平面变化的特点。从瓜德鲁普世末期到乐平世早期,海平面呈缓慢上升至缓慢下降再突然上升变化,表现为:瓜德鲁普世末期处于海退期,而乐平世早期则变为海侵阶段。 海退阶段海平面的较低水平对应着该时期主要的生物灭绝,但生物灭绝的详细步调并未与海平面的下降一致,故认为瓜德鲁普世的大规模海退与生物灭绝事件有一定关系,但不是唯一的原因。大规模海退造成海洋生物生存压力增大,尤其是生活在浅海区域的生物,同时伴随的古海洋环境发生恶化,共同导致了大规模的生物灭绝事件。
瓜德鲁普统;生物灭绝;生境型;海平面变化
瓜德鲁普世末期发生了一次早于并且独立于二叠世末期生物事件的生物灭绝事件。 蓬莱滩剖面较完整地记录了此次生物大灭绝事件(Stanley et al, 1994; Jin et al., 1994; Shen et al. 1996; Wang et al., 2000)。Shen等(2009)认为皱壁珊瑚灭绝于Jinogondolellaxuanhanensis带,接着是腕足类;蜓类灭绝于卡匹敦期的J.granti带;牙形刺带在Clarkinapostbitterihongshuiesis带开始更替;而菊石灭绝发生在吴家坪早期的C.postbitteripostbitteri带。 对于该次生物灭绝事件的成因,许多学者给出了不同的观点,如峨眉山LIP的喷发(Wignall, 2001; Courtillot et al., 2003; Wignall et al.,2009a,2009b)、全球海平面下降(Wignall at al., 2009; 邱振等,2010; Kofukuda et al., 2014; Qiu et al., 2014)、卡穆拉(Kamura)变冷事件( Isozaki et al., 2007; Kofukuda et al., 2014)以及深海缺氧(Isozaki et al., 2007; Saitoh et al., 2013)。过去有学者认为峨眉山LIP的喷发引起大陆特别是海洋环境的恶化引起生物灭绝事件(Courtillot et al., 1999; Wang et al, 2000)。但也有人则认为该次生物灭绝事件与峨眉山LIP喷发可能关系不大(Jost et al., 2014)。总之,该时期生物灭绝的原因尚未清楚,未来需要更多的研究工作去证明。
生境型是平坦底栖生物组合类型(Boucot,1981)。殷鸿福等(2004)则将生境型定义为能反应某一特定类型特定环境,具有共同特征的群落生境类型。他们都以水深作为依据对生境型进行划分(Boucot, 1981; 殷鸿福等,2004),故而生境型改变可以反映水深的变化,进而反映海平面的变化。为了进一步验证海平面变化与此次生物灭绝的关系,对蓬莱滩剖面茅口组和合山组进行了野外观察和生境型分析,希望发现更有力的证据。
1 剖面位置和区域背景
蓬莱滩剖面( 23° 41′ 43″N, 109° 19′ 16″E)位于广西来宾市以东约20 km的红水河南岸,该剖面位于来宾向斜的东翼(Wang et al., 2004)。蓬莱滩剖面上瓜德鲁普统上部卡匹敦阶和乐平统底部吴家坪阶的地层出露完整,于2006年被优先作为瓜德鲁普统与乐平统界线的全球层型剖面和点(GSSP)(Jin et al., 2006)。中二叠世茅口期全球海平面相对稳定,广西地区呈现“盆包地”的古地理格局,来宾地区处于台地边缘,发育台缘斜坡-浅水盆地相(周怀玲等,1994)。而晚二叠世吴家坪期,广西地区出现裂陷盆地和陆源碎屑盆地的复合(周怀玲等,1994),古地理格局为“陆-台-盆”相间(王根发等,1997;梅仕龙等1999),而来宾位于台地-深水盆地过渡区域,发育硅岩盆地相(梅冥相等,2004,2007;史晓颖等,2006;王新强等,2008)。
2 样品采集和分析方法
对蓬莱滩剖面茅口组和合山组进行野外岩性观测,所测量研究地层厚度约21 m,根据岩石类型及其组合关系,沉积构造等资料进行岩相分析,共采集样品53件。将样品制成薄片,使用单偏光对样品中所含生物碎片进行统计分析,同时对海绿石进行观察。
3 生境型分析
Boucot(1981)根据底栖生物组合将生境型划分为6个不同水深的生境型。殷鸿福等(1995)根据Boucot划分的生境型进一步细分为7类14个亚类。本文所用的分类方案是引用殷鸿福等(1995)划分的方案(表1)。

表1 生境型划分方案
对53个样品的岩石薄片进行镜下观察,所研究的茅口组底部为块状石灰岩,中部为中薄层石灰岩,而上部为中厚层石灰岩,而合山组为薄层状硅质岩, 根据殷鸿福等(1995)的划分方案,在所研究剖面层位共识别出4类5个亚类(图2),并且对各类生境型岩性特征进行归纳(表2)。
3.1 Ⅱ2型
主要由灰色中厚层泥粒灰岩或颗粒石灰岩组成,单层厚度大约为45~150 cm,风化面为灰白色(图3)。 偶含硅质结核或硅质条带,见少量浸染状海绿石(图3)。生物碎屑丰富,破碎严重(图1, 3),含丰富海百合、腕足、苔藓虫,海星等,有孔虫,介形虫等较常见,含少量腹足、蜓类、绿藻等(图1,3),其中第9层主要为苔藓虫、腕足和海百合生物滩(图1,3),第10层主要为海百合和海星生物滩(图1,3),部分样品发现生物扰动(图3),纹层不发育。
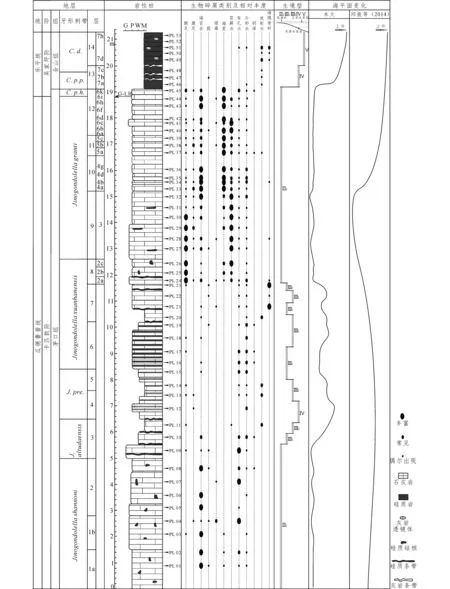
图1 蓬莱滩剖面瓜德鲁普统至乐平统综合柱状图Fig.1 The composite histogram of Guadalupian-Lopingian boundary in Penglaitan section.J. pre. = Jinogondolella prexuanhaenensis, C.p.h. = Clarkina postbitteri hongshuiensis, C.p.p. = C. postbitteri postbitteri, C. d. = C. dukouensis.G:颗粒石灰岩;P:灰泥质颗粒石灰岩;W:颗粒质灰泥石灰岩;M:灰泥石灰岩. 层号中左边为本次采样的编号,右边为Jin 等(2006)的编号;海平面变化对比于邱振等(2014)发表的海平面变化
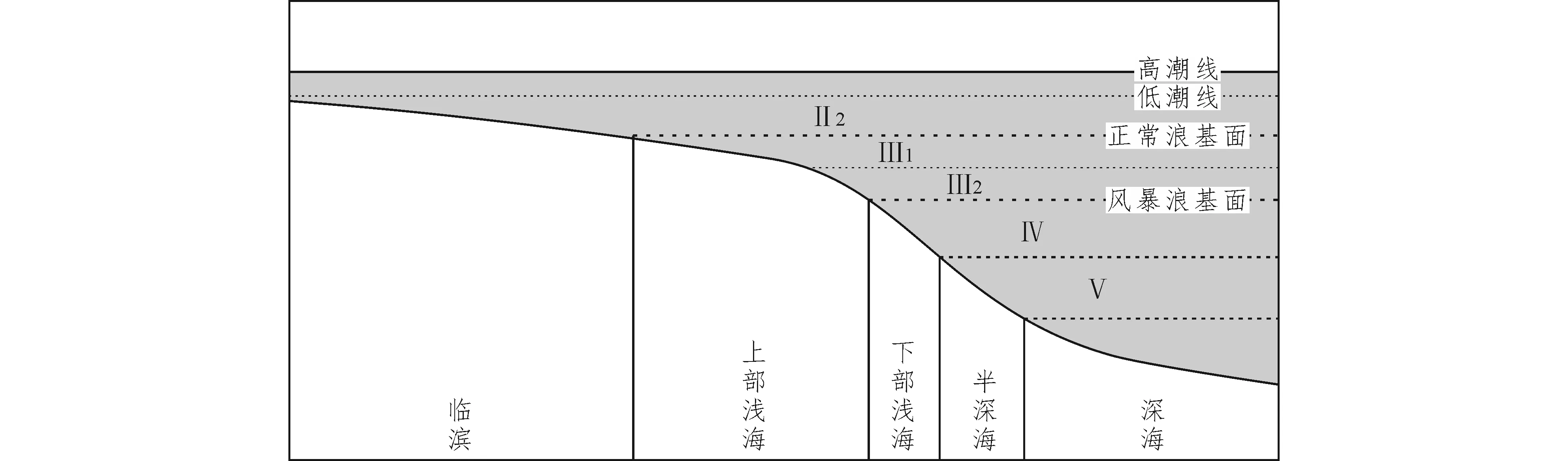
图2 来宾蓬莱滩剖面G-L界线生境型模式图Fig.2 The mode chart of habitat types across G-L boundary in Penglaitan section, Laibin

分类岩相特征Ⅱ潮间及临滨Ⅱ2临滨泥粒灰岩至颗粒灰岩(P-G),含丰富生物碎屑颗粒,颗粒边缘见磨损,部分见海绿石,无纹层Ⅲ上部浅海Ⅲ1上部泥灰岩至粒泥灰岩(M-W),含少量生物碎屑,无或弱纹层Ⅲ2下部泥灰岩(M),含海绿石,纹层较弱,含少量生物碎屑,并且碎屑颗粒细Ⅳ下部浅海Ⅳ1上部Ⅳ2下部泥灰岩(M),纹层明显,含少量放射虫Ⅴ半深海Ⅴ1上斜坡Ⅴ2下斜坡硅质岩,纹层多而明显,含丰富放射虫及硅质海绵骨针

图3 蓬莱滩剖面Ⅱ2型生境型岩相特征Fig.3 The lithofacies characteristics of habitat type Ⅱ2 in Penglaitan section.a.含丰富海百合的灰泥质颗粒石灰岩(单偏光);b.生物扰动(单偏光);c.含丰富生物颗粒的颗粒石灰岩(单偏光);d. 腕足、苔藓虫和海百合生物滩(腕足:黄色箭头;苔藓虫:黑色箭头;海百合:红色箭头. 单偏光);e,f.海百合和海星生物滩(海百合:红色箭头;海星:蓝色箭头. 单偏光);g.中厚层石灰岩野外相片(白色箭头所指,比例尺:人物身高约160 cm);h.茅口组顶部灰岩中的侵染状海绿石(绿色箭头,单偏光)
石灰岩呈中厚层状至块状,说明该时期沉积速率较慢,主要生物为海百合、腕足、苔藓虫和海星等浅海底栖生物,还观察到少量浸染状海绿石,部分样品还观察到生物扰动,说明当时沉积环境为浅水环境,并且处于比较氧化的环境。生物颗粒边缘见磨损,纹层不发育,说明水动力较大,正常浪基面以上,因此将其生境型解释为Ⅱ2型。
3.2 Ⅲ型
3.2.1 Ⅲ1型
主要由中至薄层泥灰岩或粒泥灰岩组成,单层厚度大约为30~50 cm,灰色至深灰色,风化面呈灰白色(图4a下半部),夹似层状硅质岩或与薄层硅质岩互层。含少量生物碎屑颗粒(图4b),主要生物碎屑介形虫和有孔虫(图1),含少量腕足、海百合、海星等(图1)。 部分样品发现生物潜穴,不发育纹层或者发育弱纹层(图4b)。
石灰岩主要呈中薄层,硅质岩呈似层状或薄层状,所含生物颗粒较少,主要生物碎屑为介形虫和有孔虫,浅水生物碎屑含量很少,说明水体变深。纹层不发育或者发育弱纹层,说明水体流动性大,水体深度处于风暴浪基面与正常浪基面之间上半部,所以将这类生境型划分为Ⅲ1型。
3.2.2 Ⅲ2型
主要由中厚层泥灰岩组成,单层厚度为40~60 cm,灰色至深灰色,风化面呈灰白色(图4a上半部),含硅质条带或夹似层状硅质岩,硅质岩呈灰黑色,风化面呈黄褐色。含少量生物碎屑,并且拉径段小,可见较多海绵骨针和少量放射虫(图4d,e)。少数样品观察到少量颗粒状黄铁矿和颗粒状或浸染状海绿石(图4c)。部分样品还观察到风暴沉积,发育弱纹层(图4e)。
石灰岩主要呈中厚层状,生物碎屑含量少并且颗粒粒径较小,主要生物为放射虫和海绵骨针,是深水沉积的标志之一。发现颗粒状黄铁矿,说明水体处于较还原环境, 发育弱纹层,说明具有弱水动力,并观察到风暴沉积,由此推断沉积时水体深度处于风暴浪基面与正常浪基面之间下部,所以将此类生境型划分为Ⅲ2型。
3.3 Ⅳ型
主要由薄层泥灰岩深灰色薄层硅质岩夹石灰质条带或中薄层状灰色泥灰岩组成,单层厚度为15~50 cm,风化面为深灰色或灰色(图5d)。生物碎屑少,并且颗粒细小,主要为少量放射虫(如图5a, b,c),纹层明显。
此处岩性变为以深灰色薄层状硅质岩为主,夹石灰岩条带或中薄层石灰岩,生物碎屑种类变得单一,主要为放射虫,为深水生物,说明水体变得更深,纹层明显,说明水动力较弱,未发现风暴沉积特征,推测水体深度超过风暴浪基面,因此将此类生境型划分为Ⅳ型。
3.4 Ⅴ型
主要由薄层硅质岩偶含灰岩透镜体组成,单层厚度为10~30 cm,深灰色或灰黑色,风化面为深灰色,生物碎屑种类少,含丰富放射虫和硅质海绵骨针(图6)。 纹层多而明显。
岩性为灰黑色薄层硅质岩,偶含灰岩透镜体,并且含丰富的放射虫和硅质海绵骨针,发育多而明显的纹层,是深海沉积的标志,因此将该类生境型解释为Ⅴ型。

图5 蓬莱滩剖面Ⅳ型生境型岩性特征Fig.5 The lithofacies characteristics of habitat type Ⅳ in Penglaitan sectiona,b.含放射虫硅质岩(单偏光);c. 较明显沉积纹层,放射虫(箭头)线性排列(单偏光)d.合山组底部厚至中层硅质岩野外照片(以地质锤作为比例尺)

图6 蓬莱滩剖面Ⅴ型生境型岩相特征Fig.6 The lithofacies characteristics of habitat type Ⅴ in Penglaitan sectiona.硅质岩中明显的沉积纹层,含较多小型生物碎片,如介形虫(单偏光);b.多并且明显的纹层(单偏光);c,d.硅质岩中的放射虫(箭头)(单偏光);e.硅质岩中的海绵骨针(单偏光); f.合山组薄层状硅质岩野外相片(以地质锤为比例尺)
4 生境型及海平面变化
所研究的岩层范围是从瓜德鲁普世茅口组(P2m)至乐平统合山组(P3h),由下到上地层由老到新,岩性和生境型随海平面变化而发生改变,在1,2层岩性为颗粒石灰岩或灰泥质颗粒石灰岩夹硅质结核,生境型为Ⅱ2,说明这时期海平面处于一个相对稳定状态,并且水体较浅。从第3层到第7层,岩性发生了变化,主要为颗粒质灰泥石灰岩或灰泥石灰岩夹硅质条带或者与硅质岩互层,生境型由Ⅲ1逐渐过渡到Ⅲ2再到Ⅳ,又逐渐变为Ⅲ1,并且三种生境型交替出现,出现3个小旋回,说明期间海平面总体呈现上升趋势,并且在上升过程中存在起伏。从第8层至第12层,岩性主要为颗粒石灰岩或灰泥质颗粒石灰岩,偶含硅质结核,生境型为Ⅱ2,说明海平面下降,期间海平面恢复到相对稳定状态,处于浅水环境。第13层岩性变为硅质岩夹硅质条带,生境型突然变为Ⅳ,说明在瓜德鲁普统和乐平统界限,水体突然变深,海平面上升。第14层岩性变为硅质岩偶含灰岩透镜体,生境型为Ⅴ,说明水体变得更深,海平面继续上升。所以从瓜德鲁普世末期到乐平世早期,海平面呈缓慢上升至缓慢下降再突然上升变化,瓜德鲁普统末期处于海退期,而乐平世早期则变为海侵阶段,与Qiu等(2014)所得出的海平面变化趋势总体一致。
5 讨论
对蓬莱滩剖面生境型分析瓜德鲁普统卡匹敦期早期海平面较低,到中期海平面缓慢上升,而到了晚期海平面又恢复到较低的水平。卡匹敦期海平面总体上较低,对应着全球海平面的最低值(Haq et al., 2008)。卡匹敦中期的海平面相对上升对应腕足和皱壁珊瑚的灭绝时期,而卡匹敦期晚期海平面的下降对应沉积来宾灰岩,是蜓类和牙形刺的灭绝时期。位于合山组底部的吴家坪早期,海平面出现快速上升,此时对应着菊石的灭绝高峰期(Shen et al., 2009)。这些分析表明,海平面的较低水平对应着该时期主要的生物灭绝,但生物灭绝的详细步调并没有与海平面的下降一致,说明海平面的下降至少不是该时期生物灭绝的唯一原因,尽管有不少学者认为该时期的海平面下降是主要原因(王国庆,2004; Qiu et al., 2014)。瓜德鲁普世出现显生宙Sr同位素的最低值(Veizer et al.,1999),在全球海平面下降的前提下,Sr同位素的低值反映海底岩浆活动的来源,暗示着此次灭绝事件可能与地球内部动力引起的古海洋条件的变化(如海洋初级生产力、水体氧化还原条件)存在一定联系。蓬莱滩GSSP剖面G-L界限的无机碳-有机碳差值研究发现C同位素负偏反映瓜德鲁普世末期海洋初级生产力降低(Yan et al.,2013)。日本卡穆拉剖面发现卡匹敦期存在反映高的海洋初级生产力的卡穆拉变冷事件(Isozaki et al.,2007; Kofukuda et al.,2014)。除此之外,四川广元朝天剖面发现该时期存在海洋缺氧( Isozaki et al.,2007; Saitoh et al.,2013),这些都是可能导致瓜德鲁普世生物灭绝的因素。
综上所述,生物灭绝事件是一个复杂的过程,瓜德鲁普世的大规模海退使得海洋环境恶化,海洋生物生存压力增大,尤其是生活在浅海区域的活动空间相对较窄的浅海生物,同时古海洋条件发生恶化,导致大规模的生物灭绝事件。
6 结论
对广西来宾地区蓬莱滩剖面瓜德鲁普统末期和乐平统早期生境型进行详细分析,共划分出4个大类,5个亚类,分别为的处于潮间带和正常浪基面之间的Ⅱ2型,处于上部浅海上部的Ⅲ1型,处于上部浅海下部的Ⅲ2型、处于下部浅海的Ⅳ型和处于半深海的Ⅴ型。
蓬莱滩剖面瓜德鲁普统-乐平统(G-L)界线生境型表明,卡匹敦阶处于大规模的海退阶段,并且在第9层即J.granti带上部处于最低水平面,茅口组第3层至第7层,即Jinogondolellaaltudaensis至Jinogondolellaxuanheanensis带之间发生海平面上升,在总体上升的趋势下有小幅起伏;海平面在卡匹敦阶与吴家坪阶界线处,即瓜德鲁普统与乐平统界线(G-L)处突然发生大幅度上升;而在在吴家坪阶早期海平面持续上升,变为海侵阶段。
根据海平面变化与生物灭绝发生时间的对应关系发现,瓜德鲁普世末期低海平面对应着主要的生物灭绝,但该次生物灭绝的详细步调并未完全一致,所以大规模海退不是造成此次生物灭绝的唯一原因。大规模海退可以使海洋生物活动空间减小,生存压力增大,同时海洋环境的恶化导致了此次生物灭绝。
梅冥相, 马永生, 邓军,等. 2007. 滇黔桂盆地及邻区二叠系乐平统层序地层格架及其古地理背景[J]. 中国科学 D 辑: 地球科学, 37(5):605-617.
梅冥相, 郑宽兵, 初汉民,等. 2004. 滇黔桂盆地及邻区二叠纪层序地层格架及古地理演化[J]. 古地理学报, 6(4):401-418.
梅仕龙, 朱自力, 史晓颖,等. 1999. 广西中部二叠系乐平统层序地层研究[J]. 现代地质, 13(1): 11-18.
邱振, 王清晨. 2010. 广西来宾铁桥剖面中上二叠统沉积微相[J]. 沉积学报, 28(5): 1020-1036.
史晓颖, 侯宇安, 帅开业. 2006. 桂西南晚古生代深水相地层序列及沉积演化[J]. 地学前缘, 13(6): 153-170.
王根发, 黄凤鸣, 黄乃和,等. 1997. 桂中-来宾地区上二叠统合山组层序地层分析[J]. 石油实验地质, 19(4): 348-353.
王国庆. 2004. 中扬子北缘二叠纪晚期海平面变化对生物危机事件的影响[D]. 武汉:中国地质大学(武汉).
王新强, 史晓颖. 2008. 桂西北晚古生代乐业孤立碳酸盐岩台地沉积特征与演化阶段[J]. 古地理学报, 10(4): 329-340
殷鸿福, 丁梅华, 张克信,等. 1995. 扬子区及其周缘东吴期-印支期生态地层学[M]. 北京: 科学出版社.
殷鸿福, 杨逢清, 谢树成,等. 2004.生物地质学[M]. 湖北科学技术出版社.
周怀玲, 张振贤. 1994. 广西二叠纪岩相古地理格局[J]. 广西地质, 7(4): 1-12.
Boucot A J. 1981. Principles of benthic marine palaeo ecology[M]. Academic Press, New York:1- 463.
Chen Z Q, George A D, Yang W R. 2009. Effects of Middle-Late Permian sea-level changes and mass extinction on the formation of the Tieqiao skeletal mound in the Laibin area, South China [J]. Australian Journal of Earth Sciences, 56(6): 745-763.
Courtillot V E, Jaupart C, Manighetti I, et al. 1999. On causal links between flood basalts and continental breakup[J]. Earth and Planetary Science Letters, 166(3-4): 177-195.
Courtillot V E, Renne P R. 2003. On the ages of flood basalt events [J]. Comptes Rendus Geoscience, 335(1): 113-140.
Haq B U, Schutter S R. 2008. A chronology of Paleozoic sea-level changes [J]. Science, 322(58-98): 64-68.
Isozaki Y, Kawahata H, Minoshima K. 2007. The Capitanian (Permian) Kamura cooling event: the beginning of the Paleozoic-Mesozoic Transition [J]. Palaeoworld, 16(1-3): 16-30.
Jin Y G, Zhang J, Shang, Q H. 1994. Two phases of the end-Permian mass extinction[M]. In: Embry, A.F., Beauchamp, B., Glass, D.J. (Eds.), Pangea Global Environments and Resources, 17. Canadian Society of Petroleum Geologists, Memoir:813-822.
Jin Y G, Shen S Z, Henderson C M, et al. 2006. The Global Stratotype Section and Point(GSSP) for the boundary between the Capitanian and Wuchiapingian Stage (Permian)[J]. Episodes, 29(4): 253-262.
Jost A B, Mundil R, He Bin, et al. 2014. Constraining the cause of the end-Guadalupian extinction with coupled records of carbon and calcium isotopes[J]. Earth and Planetary Science Letters, 396(12): 201-212.
Kofukuda D, Isozaki Y, Igo H. 2014. A remarkable sea-level drop and relevant biotic responses across the Guadalupian-Lopingian (Permian) boundary in low-latitude mid-Panthalassa: Irreversible changes recorded in accreted paleo-atoll limestones in Akasaka and Ishiyama, Japan [J]. Journal of Asian Earth Sciences, 82(3): 47-65.
Qiu Z, Wang Q, Zou C, et al. 2014. Transgressive-regressive sequences on the slope of an isolated carbonate platform (Middle-Late Permian, Laibin, South China) [J]. Facies, 60(1): 327-345.
Saitoh M, Isozaki Y, Yao J, et al. 2013. The appearance of an oxygen-depleted condition on the Capitanian disphotic slope/basin in South China: Middle-Upper Permian stratigraphy at Chaotian in northern Sichuan [J]. Global and Planetary Change,105(1-2): 180-192.
Shen S Z, Shi G R. 1996. Diversity and extinction patterns of Permian Brachiopoda of South China[J]. Historical Biology, 12(2):93-110.
Shen S Z, Shi G R. 2009. Latest Guadalupian brachiopods from the Guadalupian/Lopingian boundary GSSP section at Penglaitan in Laibin, Guangxi, South China and implications for the timing of the pre-Lopingian crisis [J]. Palaeoworld, 18(2):152-161.
Stanley S M, Yang X N. 1994. A double mass extinction at the end of the Paleozoic Era[J]. Science, 266:1340-1344.
Veizer J, Ala D, Azmy K, et al. 1999.87Sr/86Sr, δ13C and δ18O evolution of Phanerozoic seawater [J]. Chemical Geology,161: 59-88.
Wang W,Cao CQ, Wang Y. 2004. The carbon isotope excursion on GSSP candidate section of Lopingian-Guadalupian boundary[J]. Earth Planet. Sci. Lett, 220(1-2): 57-67.
Wang X D, Sugiyama T. 2000. Diversity and extinction patterns of Permian coral faunas of China[J]. Lethaia, 33(4):285-294.
Wei Hengye, Chen Daizhao, Yu Hao, et al. 2012. End-Guadalupian mass extinction and negative carbon isotope excursion in Xiaojiaba section, Guangyuan, Sichuan[J]. Science China Earth Sciences. 55(9): 1480-1488.
Wignall P B, Sun Y D, Bond D P G, et al. 2009a. Volcanism, mass extinction, and carbon isotope fluctuations in the Middle Permian of China [J]. Science, 324: 1179-1182.
Wignall P B, Védrine S, Bond D P G, et al. 2009. Facies analysis and sea-level change at the Guadalupian-Lopingian Global Stratotype (Laibin, South China), and its bearing on the end-Guadalupian mass extinction [J]. Journal of the Geological Society,166(4): 655-666.
Wignall P B. 2001. Large igneous provinces and mass extinctions [J]. Earth-Science Reviews, 53(1-2): 1-33.
Yan D, Zhang L, Qiu Z. 2013. Carbon and sulfur isotopic fluctuations associated with the end-Guadalupian mass extinction in South China [J]. Gondwana Research, 24(3-4):1276-1282.
Zhou M F, Malpas J, Song X Y, et al. 2002. A temporal link between the Emeishan large igneous province (SW China) and the end-Guadalupian mass extinction [J]. Earth and Planetary Science Letters. 196(3-4): 113-122.
The Habitat type and Is Sgnificance of Guadalupian-Lopingian(G-L) Bundary in Penglaitan Sction, Guangxi
WEI Xue-mei, JIANG Zeng-guang, BAI Ma-qu-zong, WEI Heng-ye
(School of Earth Science, East China University of Technology, Nanchang 330013, China)
The authors have carried on the detailed analysis of the habitat type of maokou formation to heshan formation in Penglaitan section, Guangxi. They identified 4 major categories and 5 subclasses of habitat type, it contained Ⅱ2 (between the intertidal zone and normal wave base level), Ⅲ1 (The upper part of the upper shallow seas), Ⅲ2 (the lower part of the upper shallow sea), Ⅳ (the lower shallow sea) and Ⅴ (the bathyal region). Habitat type generally reflected the characteristics of sea level change. From late Guadalupian to early Lopingian, the sea level showed a trend of changes in slowly rising to slowly falling and then suddenly rising. Late Guadalupian was in regressive period, while early Lopingian became into transgression phase.In regressive period, the comparatively low sea level corresponded to the main extinction in this period, but the detailed step of this mass extinction was not consistent with the descending of sea 1evel. So this article hold that the mass extinction in Guadalupian was associated with Large-scale regression, however, large-scale regression was not the only cause of this extinction. Large-scale regression increased the survival pressure of Marine biological, especially for the organisms living in the shallow water area, at the same time, because of the deterioration of the ancient ocean, a mass extinction event happened in end-guadalupian.
Guadalupian; Extinction; Habitat type; Sea-level changes.
2016-06-23
国家自然科学基金(41302021)
韦雪梅(1989—),女,硕士,主要从事碳酸盐岩沉积学研究。E-mail:weixuemei2015@163.com
10.3969/j.issn.1674-3504.2016.04.005
P534
A
1674-3504(2016)04-0331-10
韦雪梅,江增光,白玛曲宗,等.2016. 广西来宾蓬莱滩剖面瓜德鲁普统—乐平统(G-L)界线生境型及其意义[J].东华理工大学学报:自然科学版,39(4):331-340.
Wei Xue-mei, Jiang Zeng-guang, Bai Ma-qu-zong,et al.2016. The Habitat type and its significance of Guadalupian-Lopingian(G-L) bundary in Penglaitan section, Guangxi[J].Journal of East China University of Technology (Natural Science), 39(4):331-340.
猜你喜欢
沉积学报(2023年6期)2024-01-04 02:57:52
中南大学学报(自然科学版)(2022年9期)2022-10-27 09:28:48
中国石油勘探(2022年5期)2022-10-12 08:05:04
中国民族美术(2022年4期)2022-04-13 08:34:44
石油与天然气地质(2022年2期)2022-03-21 00:50:26
湖南水利水电(2021年6期)2022-01-18 06:07:02
疯狂英语·新悦读(2021年1期)2021-01-27 10:43:00
读友·少年文学(清雅版)(2020年6期)2020-10-20 09:30:46
小学科学(2020年8期)2020-08-31 01:38:24
石油沥青(2019年4期)2019-09-02 01:41:52
