
植物生物量分配的调节机制
2016-02-15 09:44郭海霞
安徽农学通报 2016年22期
郭海霞
摘 要:生物量分配调整是植物适应资源变化的一个重要方式,关于生物量分配调整机制的理论主要包括激素调节假说、糖調节假说、源汇假说和C-N平衡假说。其中,由于C-N平衡假说抓住了C、N两个决定生物量分配的关键因子,因而得到了许多研究者的支持。该文从C-N平衡假说的模型、发展以及局限等方面做了综述。
关键词:生物量分配;C基质;N基质;C-N平衡
中图分类号 Q948 文獻标识码 A 文章编号 1007-7731(2016)22-0020-02
Abstract:The adjustment of biomass allocation is one of the way plant adapts to resource variation. Four main theories on the drive of biomass allocation have been proposed:hormonal regulation hypothesis,sugar regulation hypothesis,source-sink hypothesis and C-N balance theory. C-N balance theory is widely supported because it is based on C,N substrate,which are two factor decide the biomass allocation of the plant. In this paper,the C-N balance model,its development and limit was discussed.
Key words:Biomass allocation;C substrate;N substrate;C-N balance
生物量分配是研究植物对环境适应策略的一个重要方向。植物对某一器官的生物量分配的增加,必然会以另一器官生物量分配的减少为代价。而不同器官代表了植物的不同功能,因此,这种生物量分配格局的变化往往反映了植物各器官功能的调节。一般认为,植物会将更多的生物量分配用于吸收受限最严重的资源。比如,地下资源限制时,植物会将更多的生物量分配给根系,而与之相对,当地上资源受限制时,会将更多的生物量分配到冠层[1-3]。然而,实际情况并没有那么简单,研究者观察到这样一些现象,CO2升高对植物生物量没有影响,K、Mg和Mn不足时,根叶的生物量分配降低[4]。由此可见,需要掌握生物量分配的调节机制,以准确把握生物分配随资源的变化。
1 关于生物量分配调节机制的理论
目前关于生物分配机制的理论主要包括激素调节假说、糖调节假说、源汇假说和Thornley的C-N平衡假说。糖假说和源汇假说中的糖分、源汇强度都是碳水化合物,可以用C来表征。激素假说中,植物体内的C、N变化引起激素的变化,然后激素又反过来调节植物的C、N[5]。由此可见,这三个理论的核心物质都是C、N。而C-N平衡假说讨论就是C、N对生物量分配的决定作用,C-N平衡假说认为,植物通过叶片和根之间的C、N转移实现生物量的重新分配,叶片和根之间的生物量分配决定于两个器官之间可利用C、N的流通[6-7]。自建立以来,C-N平衡假说也得到了许多研究者的支持[8]。
2 C-N平衡模型
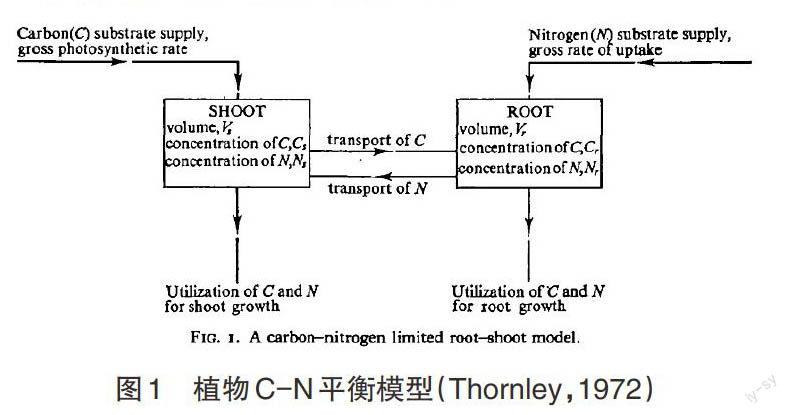
C-N平衡假说来源于Thornley的C-N平衡模型中,该模型的发展主要经历了3个阶段。其中,最具影响力的是第二阶段。C-N平衡模型可概括为以植物C、N状态为变量,根冠比为因变量的函数方程。植物的生长和生物量分配决定于C、N的转运和利用。模型适用于稳定条件下,处于营养生长阶段的植物。模型将C、N在植物体内转运和利用的过程描述如图1所示。
图1 植物C-N平衡模型(Thornley,1972)
整个模型过程可以这样描述:植物通过冠层吸收C基质,通过根吸收N基质。吸收后的基质一部分在本器官中消耗掉(此时该器官成为该基质的库),一部分运往另一个器官(此时该器官称为该基质的汇)。因此,根或冠吸收的物质主要包括3个部分,一部分用于根或冠的生长,一部分运输的其它部分,剩下的保留在基质库中。模型运用Michaelis-Menten的动力理论,计算基质使用效率。按照该理论C、N的利用速率随C、N浓度的增加呈线性增长,最后达到饱和。饱和时C的利用速率为,N的利用速率为。基质的转运速率则由源库之间的浓度和运输阻力决定,等于。
3 C-N模型的限制及拓展
C-N平衡模型建立之初,存在以下一些局限性:(1)模型以稳定状态为基础;(2)只考虑营养生长阶段;(3)没有阐明C、N基质到底是什么。基于这些限制,后面的学者做了进一步研究和推导。
首先,Werf等(1993)通过模型模拟得出,在稳定条件下得出的分配函数一样适用于不稳定状态。与C-N平衡模型一致,Werf将C、N向根、叶的生物量分配也视为植物C、N状态的函数。试验者将Dactylis glomerata先种植在全营养环境中,随后将它们种在N浓度一致的环境下,并以不同的速率向它们施加N,试验结果表明,无论在稳定条件下,还是养分变化条件下,植物对根、叶的C、N分配都与植物的N状态紧密相关,通过试验测定数据与模拟曲线的对比得出,稳定条件下得到的函数在非稳定条件下一样成立。
此外,Wilson(1988)通过总结得出,如果把繁殖器官当成一个强汇,那么在繁殖阶段,C-N平衡模型一样可行。通过花和幼果的摘除试验证明,繁殖器官的强汇作用导致根冠比发生改变。
另外,根据Thornley的假说,C-N平衡模型中的关键的物质是substrate,即基质。他所指的基质是非结构性的C、N。根据Thornley的观点,根叶之间的C、N浓度差决定了C、N向叶片和根系的转运,也间接决定了叶片和根系的生物量分配。但是,现有的试验研究资料并未证实这一点。研究者首先注意到的是叶片N含量、淀粉含量与生物分配之间的联系。Ericsson(1995)通过总结多种资源变化对植物根冠比的影响发现,与生物量分配相关的N指标主要是植物叶片N,与生物量分配相关的C,一般是叶片中的淀粉含量。随后有研究者又注意到C:N比例对生物量分配的影响。Kohl等(1998)研究发现,芦苇(Phragmites australis)的总糖/总氨基酸和天门冬酰胺/蔗糖的比例与现存生物量之间存在显著的相关性。Sarinen等(1998)和Grechi等(2007)的研究都发现,非结构性碳水化合物(TNC)与自由氨基酸(FAA)之间的比例与生物量分配紧密相关。尽管叶片和根系见C、N基质浓度差对生物量分配的决定还未通过试验证实,但是从已有的研究已经能看到C-N平衡与生物量分配的紧密联系。
4 展望
生物量分配调整只是植物响应资源变化的外在表现之一,掌握其调整的内在机制,有助于掌握植物适应资源变化的生态策略。C-N平衡模型在理论上抓住了C、N这两个关键物质,但其难点是在试验中如何对非结构性C、N进行的监测。另外,现实中,在某一时间段,C、N的来源和去向并不仅限于叶片和根系。因此,在未來的研究中,需要选择恰当的模式植物进行研究。
参考文献
[1]Brouwer,R.Nutritive influences on the distribution of dry matter in the plant[J].Netherlands Journal of Agricultural Science,1962,10(5):361-376.
[2]McConnaughay,K.D.M.,J.S.Coleman.Biomass allocation in plants:Ontogeny or optimality[J].A test along three resource gradients.Ecology,1999,80(8):2581-2593.
[3]Marcelis,L.F.M.Sink strength as a determinant of dry matter partitioning in the whole plant[J].Journal of Experimental Botany,1996,47:1281-1291.
[4]Ericsson,T.Growth and Shoot-Root Ratio of Seedlings in Relation to Nutrient Availability[J].Plant and Soil,1995,168:205-214.
[5]平晓燕,周广胜,孙敬松.植物光合产物分配及其影响因子研究进展[J].植物生态学报,2010,34(1):100-111.
[6]J.H.M.Thornley.A Balanced Quantitative Model for Root:Shoot Ration Vegetative Plants[J].Annals of Botany,1972,36:431-441.
[7]J.H.M.Thornley.Shoot-Root Allocation with Respect to C,N and P-an Investigation and Comparison of Resistance and Teleonomic Models[J].Annals of Botany,1995,75(4):395-401
[8]Wilson,J.B.A Review of Evidence on the Control of Shoot-Root Ratio,in Relation to Models[J].Annals of Botany,1988,61:433-449.
[9]Werf,A.V.d.,T.Enserink,B. Smit,et al.Allocation of Carbon and Nitrogen as a Function of the Internal Nitrogen Status of a Plant-Modeling Allocation under Non-Steady-State Situations[J].Plant and Soil,1993,155:183-186.
[10]Kohl,J.G.,P.Woitke,H. Kuhl,M. Dewender,et al.Seasonal changes in dissolved amino acids and sugars in basal culm internodes as physiological indicators of the C/N-balance of Phragmites australis at littoral sites of different trophic status[J].Aquatic Botany,1998,60(3):221-240.
[11]Saarinen,T.Internal C∶N balance and biomass partitioning of Carex rostrata grown at three levels of nitrogen supply[J].Canadian Journal of Botany-Revue Canadienne De Botanique,1998,76(5):762-768.
[12]Grechi,I.,et al.Effect of light and nitrogen supply on internal C∶N balance and control of root-to-shoot biomass allocation in grapevine[J].Environmental and Experimental Botany,2007,59(2):139-149.
(責编:张宏民)
