
红外增温对库尔勒香梨越冬期树干形成层主要生理参数的影响
2016-02-13 06:15:00艾克来木艾合买提阿不来提买买提巴特尔巴克阿丽腾吐尔孙哈力萨吉旦阿卜杜克日木祖力克艳麻那甫
干旱地区农业研究 2016年6期
艾克来木·艾合买提,阿不来提·买买提,巴特尔·巴克,阿丽腾·吐尔孙哈力,萨吉旦·阿卜杜克日木,祖力克艳·麻那甫
(1.新疆农业大学草业与环境科学学院,新疆乌鲁木齐830052;2.和田地区林业管理站,新疆和田848000)
红外增温对库尔勒香梨越冬期树干形成层主要生理参数的影响
艾克来木·艾合买提1,阿不来提·买买提2,巴特尔·巴克1,阿丽腾·吐尔孙哈力1,萨吉旦·阿卜杜克日木1,祖力克艳·麻那甫1
(1.新疆农业大学草业与环境科学学院,新疆乌鲁木齐830052;2.和田地区林业管理站,新疆和田848000)
为了探讨越冬期树干形成层温度变化对形成层组织生理参数的影响,以库尔勒香梨为实验材料,在太阳辐射和气温达到日最大值的时间段,利用控温红外灯将实验组树干阳面形成层温度分别增温达到16℃、19℃、22℃、25℃、28℃、31℃、34℃,采用热电偶温度传感器同时连续监测形成层温度并结合主要生理参数的变化,分析越冬期间树干形成层温度变化引起的融冻交替对树干组织主要生理参数的影响。结果表明,增温达到最大阳面形成层温度日较差的处理条件下,SOD、POD和CAT活性比对照分别降低了83.60%、84.62%、47.56%,相对电导率和可溶性蛋白含量分别升高了41.46%、46.81%;生理指标和形成层温度均呈负相关,其中CAT、可溶性蛋白和相对电导率呈显著性负相关。形成层温度日较差或融冻交替幅度与各生理参数均呈负相关,其中CAT、可溶性蛋白和相对电导率呈显著性负相关。保护酶系中,SOD对形成层温度变化最为敏感;虽然POD保护作用稍微迟钝,但在较强烈的形成层温度变化的条件下起到最好的保护作用;在形成层温度强烈波动的情况下,CAT保持最小变化幅度;可溶性蛋白通过调节渗透势,减轻细胞伤害程度。
库尔勒香梨;越冬期;红外增温;树干形成层温度;生理参数;融冻交替
库尔勒香梨(Pyrusbrestschneideri Rehd.)简称香梨,香梨属蔷薇科、梨属中的白梨系统,是新疆梨和西洋梨的自然杂交后代[1]。库尔勒香梨是新疆的名、优、特色水果,栽培历史已有1 500多年。库尔勒市种植面积达2.93×104hm2,总产值达9.23亿元,成为全市农业增效、农民增收以及出口创汇的主要渠道。但是,多年来冻害严重威胁着香梨产业的健康发展,对香梨产业的健康持续发展造成了重大损失。
冻害和日灼是库尔勒香梨主要的天气灾害,几乎每年都有不同程度的发生。极端低温、极端低温持续时间及它们引起的融冻交替是导致冻害、日灼综合症的主要因子[2-3]。日灼和低温引起的形成层温度差异在整个越冬期间每日都发生,因此,树皮形成层在如此大的温度波动下必定会受伤害。虽然日灼伤害不能直接导致树木死亡[4],但受日灼伤害的树干容易被病原体攻击,也能导致树木死亡[5]。国内外已报道关于日灼伤害的研究,将树干阳阴面温度、树皮颜色、树皮厚度作为重要检测指标。早在1894年Mix和Harvey用水银温度计测定过树皮下面的温度[5-7],目前,国内外对树皮形成层温度的观测基本上采用热电阻或热电偶原理的温度探头。Karels等用热电偶温度计对白桦树树皮南北侧形成层温度进行了测定,结果相差达到16℃[4]。巴特尔·巴克等采用同样的方法测定库尔勒香梨阳、阴面形成层温度,研究结果表明,阳面日较差最高值出现在冬季,达33.14℃,比阴面日较差高17.14℃,说明树干阳面融冻交替幅度比阴面大,更容易受到低温冻害和日灼危害[8]。
前人关于树木冻害和抗寒性机制的研究比较深入,以苹果[9]、梨树[10]、枣树[11]、葡萄树[12]、巴旦杏[13]、石榴[14]等的研究表结果表明树木冻害程度、抗寒性与保护酶系酶活性SOD、CAT、POD、低温半致死温度(LT50)、MDA含量、可溶性糖、可溶性蛋白含量密切相关。关于树木在越冬期间形成层红外增温和生理参数之间关系的研究较少见,本研究以库尔勒香梨为实验材料,在越冬期间通过红外增温树干形成层温度而加大融冻交替引起的伤害程度并测定超氧化物歧化酶SOD、过氧化氢酶CAT、过氧化物酶POD酶活性、相对电导率、可溶性蛋白含量等生理参数从而探究越冬期间冻害和日灼综合症对主要生理参数的影响。
1 材料与方法
1.1 实验材料
实验于2015年1月22日至2015年2月11日在库尔勒市拖布里奇乡梨园(41.67°N,85.92°E)进行,实验材料为23 a生、健康、长势相似的库尔勒香梨树。
1.2 试验方法
选7棵梨树为形成层红外增温实验处理组,5棵梨树为自然状态下各项生理指标对照组,且在实验进行同时连续测定对照组中一棵梨树50 cm高处阳阴面形成层温度。测定形成层温度时将热电偶温度传感器(型号:AV-AT)分别插入到离地面50 cm高处树皮形成层与树皮中间。插入探头时先用刀具在树皮开个1 cm2的小洞,为了便于探头插入,在开好的洞下方拨开宽0.5 cm、高3 cm的正方形树皮(不拔下来),探头从下往上顺树干插入小洞,以探头完全插入树皮内为准,方形树皮还原,用窄的透明胶与探头连线一起固定在树干上并与数据采集器相连(雨根科技有限公司,RR Data-logger,中国),形成连续监测系统,监测实验进行期间每一分钟记录一次树干南、北侧(或阳面、阴面)树皮形成层温度,根据所监测的1min的形成层温度分别计算每小时和日值。利用控温红外灯,模拟太阳辐射(从11∶00至13:30,乌鲁木齐时间)将处理组7棵梨树阳面(南面)形成温度分别逐步达到设定温度16℃、19℃、22℃、25℃、28℃、31℃、34℃,但实际操作中达到的形成层温度分别为16.34℃、19.29℃、22.43℃、25.17℃、28.14℃、31.13℃、34.19℃,连续增温22 d,处理后被增温高度阳阴面取树干形成层,用锡箔纸和纱布包扎后存放液氮罐中,送回实验室转入-80℃的冰箱中保存。树干融冻交替是由树干阳面形成层温度日最高温度和日最低温度之差来表示。
1.3 主要气象因子监测
监测离地面2 m处空气温度(型号:AV-ST)、太阳辐射(型号:AV-20P)、风速(型号:AV-30WS)等气象因子。每15min记录一次数据,监测数据采用美国Avelon公司生产的AR5型数据采集器存储,根据所监测的15 min的气象因子分别计算每小时和日值。
1.4 生理参数的测定
POD活性的测定:愈创木酚法参照郝再斌主编的《植物生理实验技术》[15]。CAT活性的测定:紫外吸收法,参照李合生主编的《植物生理生化试验原理与技术》[16]。SOD活性的测定:氮蓝四唑法,参照李合生主编的《植物生理生化试验原理与技术》[16]。可溶性蛋白含量:参照李合生[16]的方法采用考马斯亮蓝G-250染色法。
1.5 数据处理与分析
采用SPSS20、Excel 2013等软件进行数据分析,显著性分析采用Tukey’s多重测验。采用Pearson相关系数来分析相关性。
2 结果与分析
2.1 自然和增温条件下树干形成层温度的变化
图1是2015年1月24日,自然状态下24 h内树干形成层温度的变化。从图1可以看出,夜间阳阴面没有明显的形成层温度差,阳面温度稍微高于阴面,日出之后,随着太阳辐射和气温的上升,阳、阴面温度差也随着增大,12点到13点(当地时间)之间出现最大值。本实验中,形成层温度日较差为16.84℃。这种短时间内大幅度的形成层温度变化会导致阳面形成层融冻交替,影响细胞膜结构和功能。

图1 自然状态下树干形成层温度24小时变化Fig.1 The natural temperature change of cambium during a day
红外增温条件下,阳面形成层温度日较差均高于自然状态下的形成层温度日较差,平均高1.94℃、4.89℃、8.016℃、10.71℃、13.77℃、16.88℃、19.92℃,可以说明树干人工增温可以明显增大树干组织的融冻交替幅度(表1,图2)。
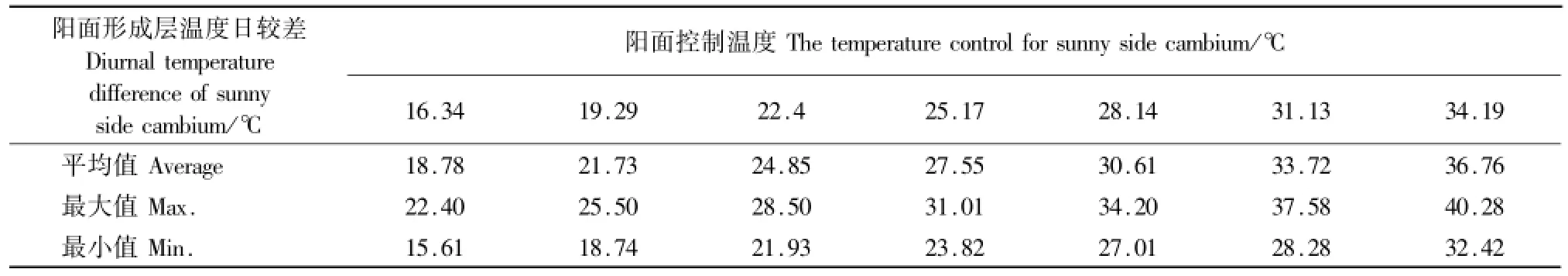
表1 红外增温条件下阳面形成层温度的变化Table 1 The temperature change of sunny side cam bium under infrared heating

图2 红外增温条件下阳面树干形成层温度24小时变化Fig.2 The temperature change of sunny side cambium during a day under infrared heating
2.2 红外增温对树干形成层生理指标的影响
2.2.1 树干形成层红外增温对SOD活性的的影响
在红外增温条件下,随着阳面形成层温度日较差的增大(图3),SOD活性先急剧上升后逐步地下降,与其它保护酶相比,SOD对形成层温度变化最先发出明显的反应,其阳面SOD活性变化的波动性大于阳面CAT活性变化。形成层增温达到16.34℃、31.13℃、34.19℃处理条件下的酶活性与对照组之间的差异达到极显著性水平,34.19℃处理组的SOD活性比对照组降低了83.60%。
2.2.2 树干形成层红外增温对CAT活性的影响CAT活性的变化趋势与SOD相似,自然状态下,CAT活性上升;在红外增温条件下,CAT活性先稍微上升后逐步地下降,阳面形成层温度增温达34℃的条件下其酶活性变化幅度比SOD和POD小,保持较高的酶活性,与对照组酶活性相比降低了47.56%,说明CAT在强烈形成层温度变化的逆境条件下,能降低活性氧自由基对细胞尤其是细胞膜的伤害。阳面形成层温度增温达到28.14℃、31.13℃、34.19℃的条件下,其酶活性与对照组之间的差异达到极显著性水平。
2.2.3 树干形成层红外增温对POD活性的影响 POD活性变化趋势与CAT、SOD不同,自然状态下,POD活性上升,在红外增温条件下,活性先降低后上升又降低,在最大阳面形成层温度日较差的条件下,与对照组酶活性相比,阳面酶活性降低了84.62%,酶活性变化幅度大于SOD和CAT。在较强烈的形成层温度波动的逆境下保持比对照组较高的酶活性,表明保护系中POD在较强的形成层温度变化的条件下起到最好的保护作用,这可能是活性氧自由基积累到一定水平后才能诱发POD保护作用的原因。阳面形成层温度增温达到16.34℃、31.13℃、34.19℃的处理条件下,与对照组之间的差异达到极显著性水平,25.17℃达到显著性差异水平(图5)。

图3 树干形成层红外增温条件下SOD活性的变化Fig.3 The changes of SOD activity in cambium under infrared heating

图4 树干形成层红外增温条件下CAT活性的变化Fig.4 The changes of CAT activity in cambium under infrared heating
2.2.4 树干形成层红外增温对可溶性蛋白含量的影响可溶性蛋白含量在增温条件下保持逐渐上升的趋势,保护系酶活性较高时,与对照相比,保持较小的变化;形成层温度较高、保护系酶活性较低时,可溶性蛋白含量明显上升,阳面形成层温度日较差最大时其含量上升了对照组的46.81%。阳面形成层温度增温达31.13℃、34.19℃的处理条件下,可溶性蛋白含量与对照之间的差异达到极显著水平,25.17℃达到显著差异水平(图6)。
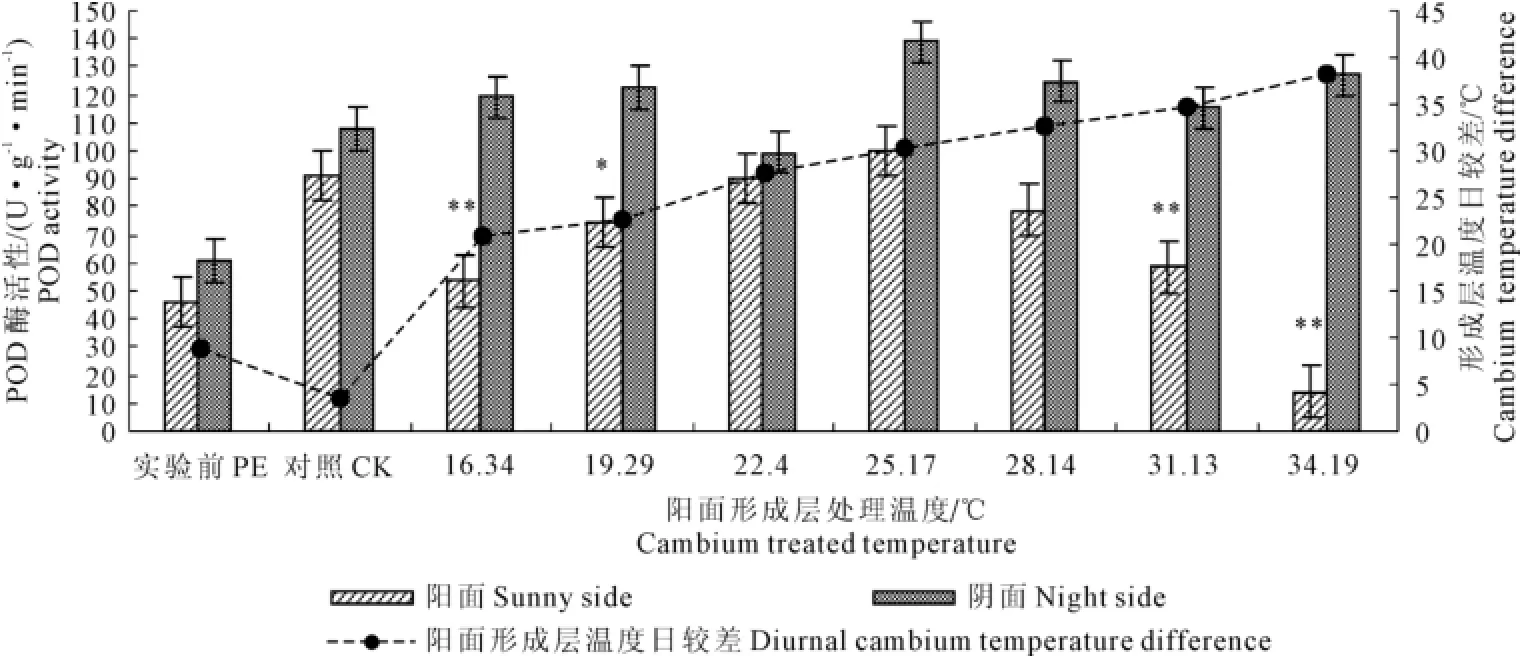
图5 树干形成层红外增温条件下POD活性的变化Fig.5 The changes of POD activity in cambium under infrared heating

图6 树干形成层红外增温条件下可溶性蛋白含量的变化Fig.6 The change of soluble protein content in cambium under infrared heating
2.2.5 树干形成层红外增温对相对电导率的影响
自然状态下,阳面形成层相对电导率稍微降低,在红外增温条件下,相对电导率随着阳面形成层温度日较差增大而上升,在最大形成层温度日较差的处理条件下,电导率比对照升高了41.46%。阳面形成层温度增温达28.14℃、34.19℃的处理条件下,其电导率与对照之间的差异达到极显著性水平。电导率增大表明,细胞膜渗透调节能力降低,细胞膜伤害程度增大,细胞内渗调节物质外渗,保护系与电导率呈负相关,其中SOD与CAT分别呈显著和极显著性负相关,证明形成层温度差引起的渗透调节物质外渗导致细胞结构和功能整体丧失,保护系酶活性降低(图7)。
2.2.6 增温条件下形成层温度与各生理参数之间的相关性分析树干形成层温度与各生理参数的变化是密切相关的(表3),红外增温条件下不同生理指标和形成层温度均呈负相关,其中CAT、可溶性蛋白和相对电导率呈显著性负相关。形成层温度日较差或融冻交替幅度与各生理参数均呈负相关,其中CAT、可溶性蛋白和相对电导率呈显著性负相关。不同生理参数中,SOD与CAT、POD呈正相关,其中与CAT呈显著性正相关(P<0.05),说明在形成层温度变化的逆境下,SOD、CAT、POD协同起到保护作用;SOD与相对电导率、可溶性蛋白含量分别呈显著性负相关;CAT与相对电导率、可溶性蛋白含量分别呈极显著性负相关(P<0.01);可溶性蛋白含量与相对电导率呈显著性正相关。

图7 树干形成层红外增温条件下相对电导率的变化Fig.7 The change of relative electrical conductivity in cambium under infrared heating
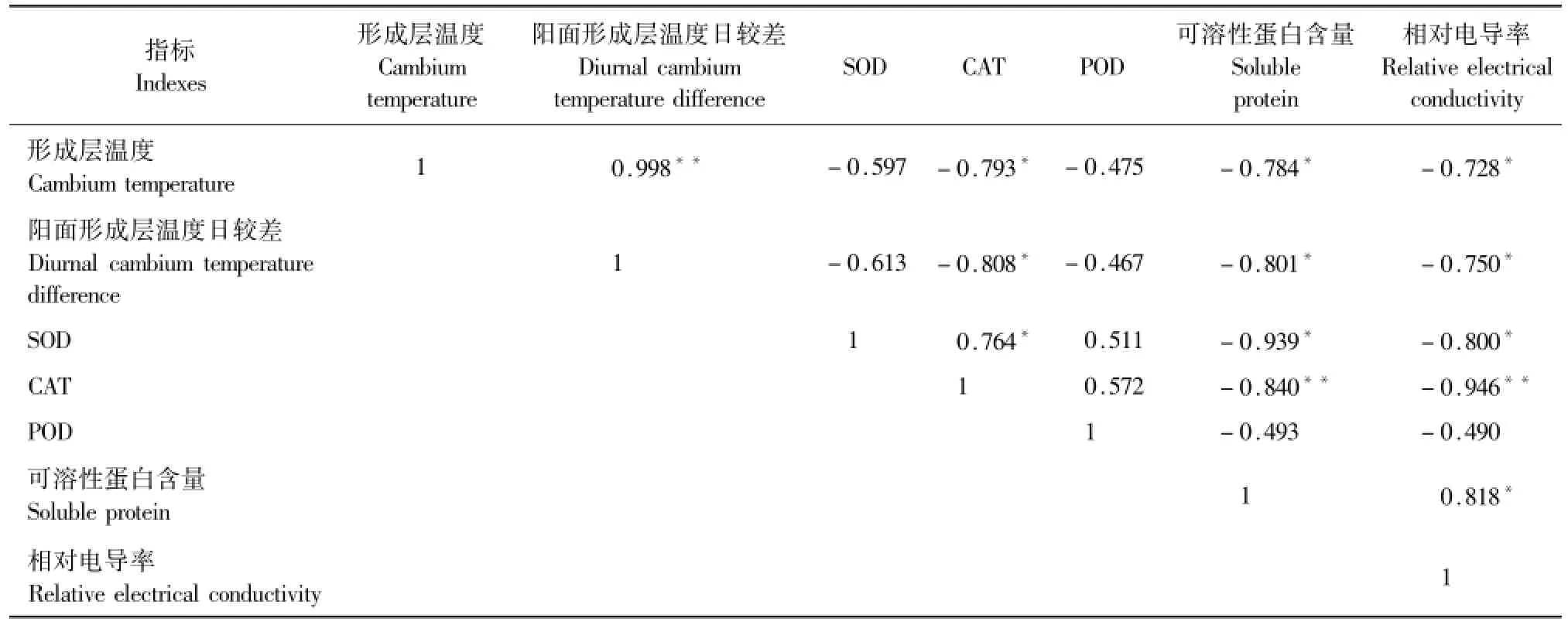
表3 形成层温度与各生理指标相关性分析Table 3 Correlation analysis between temperature and physiological indices of cambium
3 讨论
植物在逆境条件下,在细胞和组织水平上产生活性氧自由基积累,对细胞膜和代谢活动造成破坏,SOD、POD和CAT酶是机体中清除AOC和H2O2的酶,三者协调一致才能使O2-除去或者维持在较低的水平,避免自由基对机体结构和功能上的伤害,尤其是对细胞膜的攻击[18]。
前人对保护酶系与逆境条件之间的关系进行了大量的研究,均证明保护酶活性与植物的抗逆境能力有一定的关系,它们在一定程度上反映植物抵抗能力的强弱。抵抗能力强的物种或者品种在胁迫条件下,具有相对高的酶活性,许多报道认为水分胁迫下植物体内SOD活性与植物抗氧化胁迫能力呈正相关[19]。本研究进行期间果园平均气温为-16℃,阳面形成层温度日较差最大值为40.28℃,这种大幅度形成层温度变化影响细胞正常功能和结构,从而引起各相关生理指标活性和含量变化。本研究形成层温度、阳面形成层温度日较差与生理参数均呈负相关,这种结果可能是由阳面形成层融冻交替对形成层组织和细胞产生的机械伤害和低温生理伤害的综合作用引起。
以上图3~图7显示,随着阳面形成层温度的升高,SOD、CAT、POD活性下降,这可能是由强烈的形成层温度变化和低温引起活性氧自由基的积累,且积累程度超过了保护酶系调节和解除能力的阈值,使活性氧自由基和防御系统的动态平衡遭到破坏,从而发生了膜脂过氧化和渗透调节物质大量外渗,扰乱细胞正常代谢活动,影响细胞结构和功能整体性的结果。
可溶性蛋白的亲水胶体性质强[20],它能提高细胞持水能力,有利于细胞保持正常结构和功能,增加细胞浓度,提高抗寒性。本研究可溶性蛋白含量在保护系酶活性较高时保持相当低的变化,但在强烈的形成层温度波动和较低保护系酶活性的条件下明显增加,表明形成层温度变化幅度较小的条件下,保护系通过协同解除活性氧自由基对细胞膜的伤害;在形成层温度变化强烈,细胞受到一定的伤害、保护系酶不能够起到较好保护作用的条件下,可溶性蛋白含量通过调节渗透势减轻细胞伤害程度。
本实验SOD与POD、CAT活性变化趋势不同,可能是三种酶保护和抵抗机制不同的原因,与对照相比,保护酶系中,SOD对形成层温度变化最先发出反应。形成层增温处理后,POD在形成层温度变化幅度较大时能表现出较高的酶活性,这与陈少瑜等在研究坡柳抗旱树种幼苗时的结论相似[21]。异株荨麻POD与SOD活性变化趋势不同,以往认为SOD是一种诱导酶,而POD是以其为底物的氧化还原酶,在歧化活性氧时生成过氧化氢(H2O2)等过氧化物质,H2O2含量的增多使清除它的酶POD活性增强[22]。本研究表明,随着形成层温度的增高POD表现出相当高的酶活性,CAT保持最小的变化幅度,进一步证明CAT和POD是清除强烈逆境伤害作用的保护酶。
4 结论
越冬期库尔勒香梨树干形成层温度变化明显影响香梨抗逆性相关的主要生理参数,融冻交替和低温综合作用对树干必定会造成一定的影响和伤害。保护酶系中,对形成层温度变化最灵敏的是SOD;在较强烈形成层温度变化情况下,能保持比对照组较高的酶活性,而保护作用稍微迟钝的是POD;在最大形成层融冻交替幅度的条件下,保持最小变化幅度是CAT。较强烈形成层温度变化的逆境下,保护系协同起解除活性氧自由基;在强烈形成层温度变化,保护系酶活性较低的情况下,可溶性蛋白通过调节渗透势减轻细胞伤害程度。
[1]张钊,王野苹.香梨品种种源问题的探讨[J].果树学报,1993,10(2):113-115.
[2]樊丁宇,闫鹏,韩立群,等.基于果园实测气温的库尔勒香梨树越冬冻害成因分析[J].中国农学通报,2014,(7):100-103.
[3]Manion P D.Tree Disease Concepts[M].Englewood Cliff:Prentice-Hall,1991.
[4]Karels T J,Boonstra R.Reducing solar heat gain during winter:the role of white bark in northern deciduous trees[J].Arctic,2003,56(2):168-174.
[5]Mix A J.Sunscald of fruit trees:A type of winter injury[J].Cornell Univ Agric Exp Sta Bull,1916,382:235-284.
[6]Harvey R B.Cambial temperatures of trees in winter and their relation to sunscald[J].Ecology,1923,4:261-265.
[7]Boyce JS.Forest Pathology[M].New York:McGraw-Hill Book Company Inc,1961.
[8]巴特尔·巴克,海利力·库尔班,茹鲜·木沙,郑晓辉.库尔勒香梨树干阳阴面形成层温度的对比分析[J].中国农业气象,2011,32(2):185-189.
[9]闫忠业,吕天星,王冬梅,等.低温下六个苹果新品种抗性生理指标的比较[J].植物生理学报,2015,(1):93-96.
[10]龙春跃,克热木·伊力,买合木提·卡热,等.库尔勒香梨枝条、花芽冻害发生程度与相关生理指标研究[J].新疆农业大学学报,2009,32(2):6-10.
[11]王长柱,高京草,李新岗,等.西北地区枣树主栽品种抗寒性研究[J].果树学报,2011,(5):898-902.
[12]高振,翟衡,张克坤,等.LT-Ⅰ分析7个酿酒葡萄品种枝条的抗寒性[J].中国农业科学,2013,46(5):1014-1024.
[13]吉春容,邹陈,陈丛敏,等.低温胁迫下巴旦杏的形态生理变化及抗寒性研究[J].中国沙漠,2012,32(4):955-962.
[14]徐鹏.石榴冻害的发生及应对措施[J].中国园艺文摘,2014,(1):199-200.
[15]郝再彬,苍晶,徐仲.植物生理实验技术[M].哈尔滨:哈尔滨出版社,2002:204-205.
[16]李合生.植物生理生化试验原理与技术[M].北京:高等教育出版社,2000:165-169.
[17]冯献宾,董倩,李旭新,等.黄连木和黄山栾树的抗寒性[J].应用生态学报,2011,22(5):1141-1146.
[18]王娟,李德全.逆境条件下植物体内渗透调节物质的积累与活性氧代谢[J].植物学报,2001,18(4):459-465.
[19]Scandalios JG.Oxygen stress and superoxide dismutases[J].Plant Physiology,1993,101(1):7-12.
[20]梁锁兴,孟庆仙,石美娟,等.平欧榛枝条可溶性蛋白及可溶性糖含量与抗寒性关系的研究[J].中国农学通报,2015,(13):14-18.
[21]Chen SY.Effects of drought stress on lipid peroxidation and activity of defense enzymes of Dodonaea viscosa,Leucaena leucocephala and Tephrosia candida seedlings[J].Bulletin of Botanical Research,2006,26(1):88-92.
[22]董伊晨,刘悦秋.土壤水分对异株荨麻保护酶和渗透调节物质的影响及其与叶片光合和生物量的相关性[J].生态学报,2009,29(6):2845-2851.
The effect of infrared heating on physiological parameters of korla fragrant pear trunk cambium during over-wintering period
Akrem Ahmet1,Ablet Mamat2,Batur Bake1,Altun Tursunhali1,Sajida Abdukarim1,Zulkiye Manaf1
(1.College of Grassland and Environmental Science,Xinjiang Agricultural University,Urumqi,Xinjiang 830052,China;2.Forest Management Station of Hotan Prefecture,Hotan,Xinjiang 848000,China)
To explain the effect of cambium temperature changes on its physiological parameters during over-wintering period,infrared lamps was used to heat the sunny side cambium of Korla fragrant pear trunk to temperature treatment of 16℃,19℃,22℃,25℃,28℃,31℃and 34℃,respectively,during diurnal maximum solar radiation and air temperature.The actual temperature of trunk cambium was measured by thermocouple temperature sensors and its physiological factors were measured simultaneously.Results showed that,after increasing the diurnal cambium temperature difference to the maximum value,compared with the control,the activities of SOD,POD and CAT were declined by 83.60%,84.62%and 47.56%,and the relative electrical conductivity and soluble protein content were increased by 41.46%,46.81%.The physiological indexes were negatively correlated with the cambium temperature and the correlation of CAT,soluble protein content and relative conductivity was significant.The diurnal cambium temperature difference or freezing-heating cycle was negatively correlated with the physiological parameters and the correlation of CAT,soluble protein content and relative conductivity significantly.Moreover,SOD activity showed the most sensitive response to the cambium temperature change.POD has played a protection against the temperature change while its activity change was smaller than SOD.CAT activity changed the smallest.The soluble protein could relieve the degree of cell damage by adjustingthe osmotic potential.
korla fragrant pear;over-wintering period;infrared heating;trunk cambium temperature;physiological parameters;freezing-heating cycle
S661.2;S601
A
1000-7601(2016)06-0215-07
10.7606/j.issn.1000-7601.2016.06.33
2016-02-11
国家自然科学基金(31260289);新疆维吾尔自治区土壤学重点学科资助
艾克来木·艾合买提(1990—),男,维吾尔族,新疆图木舒克市人,硕士研究生,研究方向为干旱区资源与环境。E-mail:akeram27@163.com
巴特尔·巴克,教授,主要从事干旱区生态与环境研究。E-mail:bateerbake@163.com
猜你喜欢
中国果业信息(2021年6期)2021-12-02 03:59:05
军事文摘(2021年16期)2021-11-05 08:49:16
林业科学(2020年10期)2020-11-30 00:36:58
西藏人文地理(2020年4期)2020-11-19 04:32:00
发明与创新·大科技(2020年4期)2020-06-11 00:34:37
农业工程技术·综合版(2020年1期)2020-04-23 09:38:30
科学导报(2020年15期)2020-04-13 09:58:52
小猕猴学习画刊(2017年1期)2017-02-17 16:04:14
小猕猴学习画刊(2017年1期)2017-02-17 15:57:35
广东第二课堂·小学(2016年11期)2016-12-06 14:29:33
