
扦插棚内外甜樱桃矮化砧吉塞拉植株光合特性的比较研究
2016-02-05 09:38徐秀玉车兴凯高华君王家喜孙山
山东农业科学 2015年12期
关键词:光合特性
徐秀玉++车兴凯++高华君++王家喜++孙山


摘要:为给甜樱桃矮化砧吉塞拉绿枝扦插苗的大田移栽技术提供参考依据,以盆栽的‘吉塞拉6号为试材,对扦插棚内、外生长的幼苗叶片形态和光合特性进行了比较研究。结果显示:与棚外植株相比,棚内植株的叶面积、叶绿素含量分别提高了51.0%、81.2%,而比叶重、叶绿素a/b值却降低了35.9%、18.9%;棚内植株叶片的羧化效率、光合能力、光补偿点、光饱和点和水分利用效率分别下降20.59%、18.49%、45.90%、28.57%和15.98%.但表观量子效率增加29.91%;棚内植株的光合最适温度范围变宽,为25-32℃;叶绿素荧光的非光化学猝灭( NPQ)的启动速率和启动后值均明显降低。结果表明,低光、高湿环境下生长的棚内幼苗不仅光合能力弱,热耗散能力也低,即使经过20天的适应锻炼,其叶片结构和光合功能仍没有达到自然光下发育叶片的水平。
关键词:吉塞拉;扦插棚;光合特性;叶绿素荧光
中图分类号:S662.501
文献标识号:A 文章编号:1001-4942 (2015)12-0020-04
甜樱桃矮化砧木吉塞拉是德国以欧洲酸樱桃(Prunus cerasus)与灰毛叶樱桃(P.canescens)进行种间杂交培育的三倍体杂种,其与大多数甜樱桃品种嫁接亲和良好,具有明显的矮化、早实、丰产等优点。由于吉塞拉为无性系砧木,不能采用种子繁殖,而常规压条及硬枝扦插生根又很困难,其苗木繁育目前多采用当年生的嫩梢进行绿枝扦插。吉塞拉绿枝扦插在大棚内进行,插条的生根、生长均处于适宜的光照、湿度及温度等环境条件下,夏季移栽大田时,会遭遇强光、相对湿度降低等环境胁迫,成活率较低。
本研究以扦插棚内、外生长的吉塞拉盆栽苗为材料,重点比较两种环境下植株的光合特性差异,为制定提高大田移栽成活率的技术措施提供参考依据。
1 材料与方法
1.1 材料
试验于2014~2015年在山东省果树研究所试验场进行。春季发芽前,取1年生‘吉塞拉6号(PnLnus cerasus×P.canescens)扦插苗进行盆栽,盆口直径20cm,高18cm。每盆装配制营养土(熟园土:沙:干鸡粪=7:2:1)3.5kg,种植1株。将10株置于自然光强下,10株置于扦插棚内。正常浇水和病虫害防治管理。8—9月间对棚内苗适应锻炼(去遮阴网,放风降温、降湿)20天后,每株固定选取新梢中部完全展开的功能叶测定。
1.2 测定项目及方法
1.2.1 光合气体交换参数的测定采用CIRAS-3型便携式光合系统(PP Systems,美国)测定,测定光强、CO2浓度和叶温等均由CIRAS-3自控系统控制。光合启动和水分利用效率(WUE)测定时,光强为1000 μmol.m-2-2.s-1,C02浓度为370μmol·mol-1,叶温为(25±0.2)℃,相对湿度为75%。水分利用效率的计算参照Martin等的方法;光合响应曲线的测定参考孙山等的方法。根据光强一光合(Pn-PAR)响应曲线,计算表观量子效率、光补偿点,估算饱和光强、最大净光合速率(A);根据C02-光合(Pn-C02)响应曲线和细胞间隙C02-光合(Pn-Ci)响应曲线,计算羧化效率、C02补偿点、C02饱和点,以气孔限制值(Ls)为零(以Ci为370μmol.mol-1计算)时的光合速率代表光合能力( A370),气孔限制值按Ls=(1-A/A370)×100%计算。测定温度一光合(Pn -T)响应曲线时,从15~45℃分13个梯度,在每一温度下适应10min后进行测定。
1.2.2 叶绿素荧光参数的测定叶绿素荧光参数采用FM-2型便携脉冲调制式荧光仪(Hansatech,英国)测定。将待测叶片暗适应30min后测最大荧光(Fm),然后将叶片置于1000μmol.m-2.s-1的光强下,每隔20 s测1次光下最大荧光(Fm')、稳态荧光(Fs),计算实际光化学效率(φPSⅡ=1-Fs/Fm')和非光化学猝灭系数(NPQ=Fm/Fm' -1)。
1.2.3 光合色素含量测定 叶片中叶绿素a、叶绿素b、类胡萝卜素含量和叶绿素a/b(Chla/b)参照AmOn的方法测定。
1.2.4 叶面积及比叶重测定 叶面积采用Li -3000型自动叶面积仪(Li-COR,美国)测定,测定后将叶片105℃杀青10min,75℃烘干48h,取出称其干重,计算其比叶重:比叶重=叶干重/叶面积。
1.3 数据统计及处理
采用Microsoft Excel 2007进行试验数据分析,Sigma Plot 10.0软件作图。
2 结果与分析
2.1 棚内外吉塞拉植株叶片形态及其光合色素含量的差异
由表1可以看出,棚内植株的叶面积比棚外植株提高了51.0%,而比叶重却降低35.9%。棚内植株叶片的光合色素含量明显高于棚外植株,其中叶绿素a、叶绿素b、类胡萝卜素含量分别增加了73.2%、116.7%、62.5%,导致棚内植株叶片的叶绿素a/b比值(Chla/b)明显低于棚外植株。表明,在棚内生长条件下,植株营养生长受到促进,叶绿素含量尤其叶绿素b含量提高,因而可以增强捕获光能的能力,适应棚内低光环境,但叶片质量明显下降。
2.2 棚内外吉塞拉植株叶片的光合作用对光合有效辐射的响应
根据Pn-PAR响应曲线,计算所得相关参数见表2。结果表明,与棚外自然光下生长的植株叶片相比,棚内植株叶片的最大净光合速率、光补偿点、光饱和点和水分利用效率分别下降27.22%、45.90%、28.57%和15.98%,但表观量子效率增加了29.91%。受光强的影响,棚内吉塞拉植株叶片具有部分阴生叶的特点,以适应棚内的低光、高湿环境,对强光、干旱的适应调节能力较弱。endprint
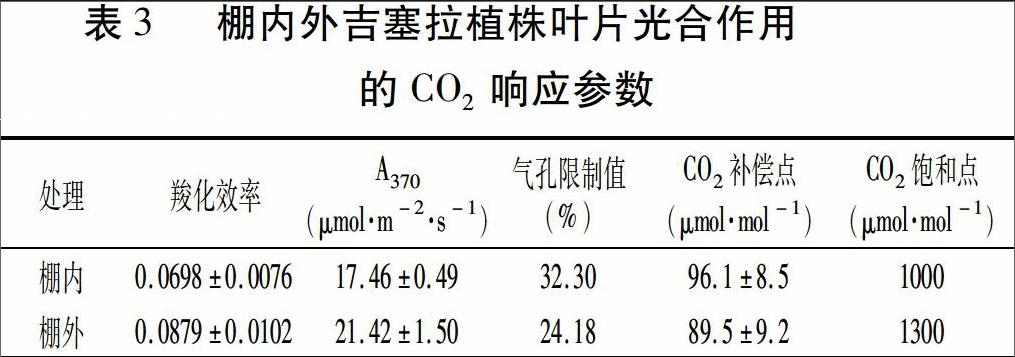
2.3 棚内外吉塞拉植株叶片的光合作用对C02浓度的响应
根据Pn-C02、Pn-Ci响应曲线,计算所得相关参数见表3。结果表明,棚内植株叶片的羧化效率和光合能力(A370)分别比棚外植株低20.59%和18.49%,而C02补偿点比棚外植株高7.37%,暗示着其光合暗反应活性及CO2利用率低于棚外植株。此外,在相对湿度较低的自然条件下测定,棚内植株叶片的气孔限制值比棚外植株增加了8.12个百分点,表明棚内植株耐旱性弱于棚外植株。
2.4 棚内外吉塞拉植株叶片的光合速率对温度的响应
由图1可以看出,非强光下,棚外植株的光合最适温度为25~30℃,而棚内植株的光合最适温度范围变宽,为25~32℃。高温下棚内植株光合速率的下降明显慢于棚外植株,这可能与炼苗期间育苗棚内中午的温度较高有关。
2.5 棚内外吉塞拉植株叶片光化学效率的比较
φPSⅡ的初始斜率表示光化学反应的启动速率,由图2-A及表4可以看出,棚内植株叶片的光化学反应启动较快,表明当从暗处暴露到光下时,其光化学反应能够快速地启动,在最短时间内提高对光能的利用。但当光化学反应启动后,棚外植株的φPSⅡ大于棚内植株,表明光化学反应启动后棚外植株对光能的利用率高于棚内植株。植物吸收光能后一方面进行光化学反应,一方面将剩余的激发能大部分以热的形式耗散掉,非光化学猝灭系数(NPQ)反映植物热耗散能力的变化。棚内植株叶片的NPQ启动速率和启动后值均明显降低(图2-B,表4),表明其热耗散能力低于棚外植株。
3 讨论与结论
植物在低光下生长会引起叶片在形态和结构上发生变化,部分具有阴生叶的特点,主要表现为叶片变薄、叶面积适当增大、叶绿素含量增加等,这是植物对弱光环境的一种适应,有利于植物捕获较多的光能。本试验结果表明,棚内吉塞拉植株叶片的光合色素含量、表观量子效率明显高于棚外植株,但是叶片质量、羧化效率、光合能力、光补偿点、光饱和点均低于棚外植株,表明棚内植株叶片已适应低光环境,对强光的适应调节能力较弱。光是植物进行光合作用的能量来源,但如果其吸收的激发能不能被及时有效地耗散掉,过剩激发能会产生大量的活性氧分子,对光合器官和其它组织造成伤害,严重时造成植物死亡。当棚内扦插幼苗移栽至大田时,剧烈的强光胁迫将成为其面临的首要障碍。
空气湿度对植物的生长与发育也有重要影响,高湿度能增加叶面积,使气孔增大,提高气孔导度和蒸腾速率,同时引起气孔对失水的反应迟钝,使其关闭速度慢于低湿度下生长的叶片。本研究中,棚内吉塞拉植株叶片适应棚内的高湿环境,水分利用效率比棚外植株下降了15.98%,对干旱失水的调节保护能力较差。叶片失水会对光合结构,特别是光系统I造成严重伤害。大田空气湿度远低于育苗棚室,会进一步加剧扦插幼苗的光抑制,也是其必须适应的环境变化因子。
在长期进化过程中,植物为了适应多变的生长环境,形成了多种耗散过剩激发能、避免光合机构遭受光破坏的保护机制,其中依赖叶黄素循环的热耗散是最重要的保护机制之一。当激发能过剩时,捕光天线色素蛋白复合体则会启动热能耗散,来减轻光抑制;棚内植株叶片的NPQ的启动速率和启动后值均明显降低,表明其热耗散能力低于棚外植株。
本试验中棚内植株在中低光下对高温的耐受性增强,可能与后期去除遮阴网造成午间棚内极端温度较高有关。夏季大田自然条件下,高温常常是伴随着强光出现的,单纯的人为变温处理与大田条件下发生的光、温协调变化相差较大,而且影响光合作用的另一重要生态因子空气相对湿度也随着光、温的变化而变化。因此,移栽大田后自然高温对植株的影响仍需进一步研究。
综上所述,棚内吉塞拉植株不仅光合能力较低,热耗散能力也较低,即使经过适应锻炼后,其叶片结构和光合功能仍未达到自然光下生长的植株叶片水平。endprint
猜你喜欢
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
湖北农业科学(2015年16期)2015-10-28
长江蔬菜·学术版(2015年1期)2015-07-10
长江蔬菜·学术版(2015年6期)2015-06-24
天津农业科学(2015年6期)2015-06-16
