
睡眠影响记忆巩固的同步EEG-fMRI研究*
2016-02-02 12:24:50
心理科学进展 2016年3期
(西南大学心理学部; 认知与人格教育部重点实验室, 重庆 400715)
1 问题的提出
学习后睡眠有利于记忆的巩固, 越来越多的证据表明记忆材料的类型、记忆编码的过程以及学习和睡眠的间隔时间等都会对其后的记忆巩固产生影响, 相应的大脑激活和睡眠指标差别很大(Lewis, 2014)。目前该领域的研究集中在动物实验和人体的行为学水平, 人脑神经成像的研究还比较有限, 更缺乏相关的动态脑网络分析。由于本研究主要采用人体实验, 现将论述研究中涉及到的行为学、EEG和fMRI的神经成像研究, 并分析国内外该领域的发展动态。
1.1 睡眠依赖记忆巩固的行为学研究
睡眠依赖记忆巩固的行为学研究始于动物的睡眠剥夺实验。早期研究证明了睡眠剥夺、部分睡眠剥夺、片段化睡眠都可能导致小鼠学习和记忆成绩的下降。之后的人体研究重点关注了记忆类型和睡眠阶段的关系(Lewis, 2014)。根据多导睡眠图的特点人们把睡眠分为前半夜占优势的非快速眼动睡眠(non-rapid eye movement, NREM)和后半夜占优势的快速眼动睡眠(rapid eye movement,REM)。NREM睡眠包括浅睡期和深睡期, 后者由于其明显的慢波振荡又被称为慢波睡眠(slow-wave sleep, SWS)。记忆根据有无意识的参与被分为陈述性记忆和程序性记忆。前者与事件、语义等相关, 后者与感知觉、技能等相关。在SWS睡眠阶段大量证据支持了其有助于陈述性记忆的巩固和增强, 这些证据包括:经过SWS睡眠后被试数字记忆的正确率提高, 单词回忆的数量增多等(Gais et al., 2007)。REM睡眠主要对程序性记忆及有情绪色彩的陈述性记忆具有巩固作用, 这在视觉分辨、动作学习和序列按键等多个任务中得到证实。这些研究通常在睡眠前后进行反应时和正确率的测试。测试成绩在睡眠后高于睡眠前, 而且不易受新任务的干扰(Karni et al., 1995)。
根据以上研究结果人们发展了多种理论用于解释睡眠依赖的记忆巩固, 其中比较有代表性的是序列假说和双过程假说(Landmann et al., 2014)。序列假说认为SWS和REM两种睡眠的有序推进是记忆巩固的关键。如果干扰学习后这两种睡眠的循环, 被试在后测中的成绩就不会提高(Barrett& Ekstrand, 1972)。双过程假说则认为两种睡眠各有侧重:陈述性记忆巩固主要在SWS睡眠期而程序性记忆和有情绪色彩的陈述性记忆巩固则主要在REM阶段发生。随后人们结合脑成像的结果还提出了海马−内侧前额叶依赖假说:认为清醒时与个人未来计划相关的记忆会激活海马−内侧前额叶系统, 睡眠过程中这些记忆通过海马−内侧前额叶之间的对话得到巩固。默认网络由静息状态大脑自发活动较强的多个脑区构成, 包括了海马和内侧前额叶等。最近的证据表明静息状态也有离线记忆巩固的发生, 其核心结构就是默认网络的腹侧区域(Sami, Robertson, & Miall, 2014)。因此在本研究中我们推测默认网络是睡眠依赖记忆巩固的核心结构, 并开展睡眠过程记忆巩固的动态观察, 以及昼夜更迭过程中海马和内侧前额叶的激活分析, 有望建立基于默认网络的睡眠依赖记忆巩固的新理论。
1.2 睡眠依赖记忆巩固的EEG研究
EEG由于其毫秒量级的时间精度成为研究记忆巩固动态过程的有力工具。目前该领域的研究集中在:记忆巩固的动态电生理过程; 纺锤波和慢波振荡的分类及其在记忆巩固中的作用等, 下面分别予以介绍。
记忆巩固的动态过程主要通过动物的有创性电生理实验得到。这些研究表明觉醒时新皮层和海马会平行地对信息进行编码。海马在随后的SWS睡眠期出现了记忆的重演(replay), 而丘脑则产生了纺锤波。新皮层发出的慢波振荡使各脑区的活动达到了同步从而促进了纺锤波−涟漪波事件的发生。在慢波振荡的去极化阶段纺锤波到达特定脑区, 通过一系列生物化学变换实现了记忆痕迹向新皮层的转移(Diekelmann & Born, 2010)。目前该生理过程尚缺乏人体实验中的直接证据,涟漪波等深部脑电活动能否用无创的头表EEG进行观测也存在问题。本研究拟结合源定位和因果连接分析来构建记忆巩固的动态过程。之前的研究将EEG每个电极作为节点, 以电极间信号的因果关系作为边来构造头表网络。记忆编码过程的头表网络研究表明前额叶和顶叶的θ节律(2~4 Hz)所形成的网络具有较高的信息传输效率, 而且可以通过训练进一步提高这一效率(Langer, von Bastian, Wirz, Oberauer, & Jäncke, 2013)。这些研究普遍存在的弊端是建立的头表脑网络无法规避多个脑区信号叠加所致的模糊性。解决方法之一就是采用源定位技术或融合 fMRI的激活结果来重建大脑层面的电活动。在之前的工作中我们提出了基于fMRI功能网络的源定位技术(Lei et al.,2011), 本研究我们尝试把该技术引入到睡眠脑电分析中发展皮层节律的网络构建技术, 探讨记忆巩固的动态电生理过程。
纺锤波是浅睡期EEG的特征波形, 大量证据表明其在记忆巩固中扮演了重要的角色。在陈述性记忆训练后通常可以观察到纺锤波个数的增加。在一项采用不同情绪人脸图片的记忆任务中图片再认的正确率与纺锤波个数呈正相关(Wagner, Valero-Cabre, & Pascual-Leone, 2007)。最近人们认识到纺锤波其实包含快纺锤波(12~15 Hz)和慢纺锤波(9~12 Hz)两个子成分(Mölle,Bergmann, Marshall, & Born, 2011)。快纺锤波能促进学习后慢波振荡和慢纺锤波的出现, 其可能在睡眠依赖的记忆巩固中发挥了更重要的作用。慢波振荡(约0.75 Hz)是SWS睡眠期EEG的特征波形, 是记忆巩固过程中海马和新皮层进行对话的关键。图片学习后前额叶在浅睡期和枕叶在深睡期都可以观察到慢波振荡幅度的提高, 且其增加程度与行为成绩正相关(Marshall & Born, 2007)。近年来40~100 Hz的γ节律也受到关注, 睡眠过程中的γ节律集中出现在慢波的峰值阶段(Valderrama et al., 2012), 提示在动物实验中发现的频率间的跨频段耦合(cross-frequency coupling)可能也是人类记忆巩固的基础。γ节律的主要功能是特征绑定, 而慢波振荡相对于 γ节律具有更长的振荡周期和更好的传播性。动物实验发现由 γ节律形成的局部耦合能通过θ节律在更远的范围内形成同步从而达到远程协作的目的(Colgin, 2013)。以上研究囿于脑电的空间精度, 无法区分不同类别记忆巩固过程特异的纺锤波和γ节律, 需结合fMRI开展脑区定位研究。鉴于 γ节律与学习记忆过程的密切联系, 我们尝试在本研究中探讨 γ节律如何参与睡眠依赖的记忆巩固。
1.3 睡眠依赖记忆巩固的fMRI研究
fMRI凭借其空间分辨率优势, 为研究不同类型记忆巩固提供了更多的脑区激活信息。目前基于 fMRI的记忆巩固研究主要取得了以下进展:发现了海马和新皮层的重激活现象; 记忆任务完成后数月内激活区域出现了从海马到新皮层的转移。
根据记忆分类的不同, 陈述性记忆重激活发生在海马和内侧前额叶, 而程序性记忆发生在海马和感觉运动皮层或其他记忆编码脑区。早期研究观察到海马和感觉运动皮层在 SWS睡眠中有重复之前记忆编码过程中激活模式的情况, 推测这种活动可能就是动物实验中所发现的重演现象(Rasch & Born, 2007)。后来越来越多的实验重复了这一重激活现象, 而且发现睡眠过程中重激活越多记忆巩固的效果就越好。重激活是否伴随着记忆编码过程脑连接的重现是个值得探讨的问题,需借助滑动时间窗法研究脑连接的瞬时变化。相对于传统网络构建方法在整个扫描期间(约 5~10分钟)计算一个脑网络, 滑动时间窗法将数据进行分割然后分别构建脑网络(Allen et al., 2014)。这有利于分析网络属性随时间改变的情况, 比如小世界性和模块性的动态变化。模块是网络中内部连接密集但对外连接稀疏的节点集团, 最近我们分析了睡眠剥夺后脑网络的模块性, 发现睡眠缺失后默认网络被分裂为两个模块(Wang, Liu,Hitchman, & Lei, 2015)。本研究拟采用时间窗法考察睡眠过程默认网络的动态变化, 探讨重激活中是否存在编码过程脑连接的重现。
fMRI研究的另一个发现是观察到记忆习得后较长时间里记忆提取激活的区域从海马转移到新皮层。单词学习 2天后, 记忆检索过程海马的活动会增强, 同时海马与内侧前额叶皮层间的功能连接也得到加强。但6个月后记忆检索激活的区域有所转移:内侧前额叶皮层有更大的激活并且海马与内侧前额叶间的连接减弱了(Gais et al.,2007)。采用带有情绪效价的图片刺激也重复了这一发现:内侧前额叶的激活随着时间的延长而提高而海马的激活却降低(Takashima et al., 2006)。这些实验都说明随着时间的推移, 暂时储存于海马的记忆信息逐渐转移至皮层使记忆表征在系统水平发生了改变。在睡眠中给予记忆线索刺激会导致记忆的重激活。已有实验采用物体−位置匹配任务, 在睡眠过程提供记忆编码阶段的气味实现了记忆的重激活, 并发现海马和压后皮层的活动,但该研究没有讨论较长时间里重激活脑区是否有转移现象(Diekelmann, Büchel, Born, & Rasch,2011)。根据记忆提取过程的这些研究, 我们推测睡眠过程的记忆重激活可能也会发生重激活脑区从海马到新皮层的转移。
1.4 睡眠依赖记忆巩固的多模态研究
随着神经干预和成像技术的发展, 目前记忆巩固的研究表现出一些新的特点:视角上将静息态和睡眠过程结合、技术上将电生理和神经成像结合, 呈现出多种模态同时采用的局面。
记忆编码后的静息态大脑自发活动, 既可能反映了编码的后效应, 也可能表明已经开始了记忆的进一步加工。有研究发现在顺序按键任务中观察到训练后静息态双侧感觉运动皮层连接的增强, 该连接在先前的学习过程中并不存在, 也与训练中的行为成绩无关, 但这一连接却与之后睡眠导致的记忆巩固显著相关(Gregory et al.,2014)。在序列反应时任务(serial reaction time task,SRTT)中研究者区分了外显学习和内隐学习(即是否提示被试任务中有固定的按键序列)不同的静息态活动, 时间节点选择了学习后10分钟内、半小时后、和6小时后。外显学习中静息态的连接经历了前额叶、深部脑区、感觉运动皮层的变迁,而内隐学习则是感觉运动皮层、深部脑区、最后才到内侧颞叶(Sami et al., 2014)。如果静息态和睡眠一样都有离线的记忆巩固功能, 对比两者的异同将是今后研究的重要方面。由于大脑的在线更新速度不同, 之前的研究发现静息态 fMRI信号的长时记忆性(即幂律指数)比任务态大(He,Zempel, Snyder, & Raichle, 2010)。我们最近的研究讨论了信号长时记忆性的个体差异和人格特质的关系, 发现外向性人格得分与大脑默认网络的长时记忆性高度相关, 推测外向的人可能具有更好的在线处理能力(Lei, Zhao, & Chen, 2013)。本研究中我们将对比静息态和睡眠状态不同的离线巩固机制, 特别是默认网络各脑区自发振荡的长时记忆性, 外显学习后的睡眠阶段可能会加快长时记忆性在不同脑区的转移。
睡眠的神经成像研究需结合电生理指标进行,通常的做法是交替采集:在 fMRI扫描间隔收集EEG信号。这使得在后处理中可以根据电生理指标对 fMRI扫描进行睡眠阶段的划分。但该方法一方面丢弃了多导EEG所具有的空间信息, 另一方面也忽略了许多睡眠特异的脑电波成分, 如纺锤波、γ节律等。一种有效的解决思路是同时采集多导EEG和fMRI信号, 通过对称的数据融合进行动态脑网络研究(Debener, Ullsperger, Siegel,& Engel, 2006)。这一同步EEG-fMRI技术为睡眠研究注入了新的活力。首先, 同步技术为高级脑功能的研究提供了更高的时空分辨率。由于可以更为精细的观测脑的动态变化过程, 使得该技术特别适合于研究睡眠阶段记忆巩固。这将有利于我们分解记忆巩固的不同加工阶段, 以及在同一加工阶段中区分不同的功能结构。其次, 该技术有效的保证了多模态研究过程中被试状态的统一性。同步采集可以保证实验环境、刺激条件、被试的情绪、警觉水平等参数高度一致。最后, 该技术提供了电生理和功能代谢两个指标, 有利于结合电生理和功能代谢两方面已有的工作, 将记忆巩固相关的神经生理机制和大脑结构定位相结合, 得到更为明确收敛的证据(雷旭, 刘欢, 李红,2013)。本研究中我们采用同步EEG-fMRI开展睡眠依赖的记忆巩固研究, 一方面有利于澄清睡眠特异性脑电波的空间分布规律, 区分不同类别记忆巩固过程所特异的脑电波, 解决纺锤波、γ节律的神经产生源问题。另一方面, 探讨记忆习得后的不同时间节点记忆重激活的动态过程, 回答睡眠过程记忆的重激活是否经历了从海马到新皮层的转移过程。我们最近采用同步EEG-fMRI, 观察到默认网络受任务正网络的抑制程度与头表θ节律高度相关, 预示着EEG节律可能反映了大尺度脑网络间的功能连接(Lei, Wang, Yuan, & Mantini,2014)。本研究将延续之前在 fMRI功能连接和EEG节律关系的研究, 重点关注睡眠过程默认网络功能连接和慢波振荡的关系。
2 研究构想
综上所述, EEG的研究发现了以慢波振荡、纺锤波、γ节律为代表的多种睡眠依赖记忆巩固的电生理特征, 而 fMRI的研究则观察到包括内侧前额叶、海马为代表的多个脑区的信息交互。但由于这些技术本身的局限仍无法呈现睡眠依赖记忆巩固在毫秒量级的动态过程, 相关的电生理特征和脑区在记忆巩固过程中的具体功能亟待澄清。基于以上问题本研究拟借助同步 EEG-fMRI在时空分辨率上的优势, 发展一系列动态脑网络分析方法, 研究睡眠依赖记忆巩固的神经机制。主要包括:1)构建默认网络活动的电生理指标, 应用EEG源定位和跨频段耦合分析, 揭示记忆巩固的动态过程; 2)应用滑动时间窗和模块分析, 研究静息态和睡眠过程中默认网络参与离线记忆巩固的异同, 揭示记忆在昼夜更迭过程中得以强化的基本原理; 3)通过多模态数据融合, 揭示记忆类型和睡眠阶段等因素对睡眠依赖记忆巩固的影响,试图建立基于默认网络的睡眠依赖记忆巩固的新理论。在本工作中我们将采用同步EEG-fMRI技术, 从皮层节律分析, 相位−幅度耦合分析、滑动时间窗、动态模块分析和多模态融合5个方面, 对睡眠依赖记忆巩固进行全方位的诠释。主要包括以下3组实验。
2.1 记忆任务
本部分拟借助EEG在时间精度上的优势, 重点分析慢波睡眠和快速眼动睡眠中记忆巩固的动态脑网络特征。主要包括三个方面的工作:第一,在我们之前提出的基于 fMRI功能网络的源定位基础上发展睡眠脑电的皮层节律定位技术, 构建SWS和REM睡眠期默认网络主要节点间的因果连接, 睡眠分期参照美国睡眠医学会(AASM)2007版(Iber, Ancoli-Israel, Chesson, & Quan, 2007)标准进行; 第二, 采用陈述性记忆和程序性记忆任务, 探讨记忆类型不同所致的纺锤波在空间分布、频率范围上的改变, 进一步结合fMRI信号找出不同纺锤波的产生源; 第三, 开展 γ节律调节记忆巩固的实证研究, 特别是前额叶慢波振荡与γ节律间的相位−幅度耦合关系, 对比陈述性记忆和程序性记忆任务中相位−幅度耦合的异同。通过本部分的研究有望构建出不同类型记忆巩固的动态电生理过程。并为之后的同步EEG-fMRI研究奠定数据收集和分析的基础。
陈述性记忆任务采用词对作为材料, 从现代汉语双字形容词感情信息评定表中选择100个中性词作为目标刺激, 形容词的情绪效价强度控制在4到6之间。选词标准包括:双字词汉字无重复、笔画中等、均为常用词。词对匹配完全随机,每位被试的学习材料(词对)均为随机生成且互不相同。实现流程包括:学习阶段安排在第一天临睡前的晚上22:00到23:30之间, 50个词对随机呈现。每个词对呈现3 s, 间隔为1 s采用注视点“+”呈现; 测试阶段在屏幕上呈现线索词要求被试大声快速的说出目标词, 在4 s后无论答案是否正确均呈现正确答案 3 s。如果被试的正确率达不到60%以上将重复学习和测试阶段, 达到正确率标准后进入后面的睡眠阶段同步 EEG-fMRI采集;睡眠后测试在第二天凌晨3:00左右进行。为保证被试的状态和睡眠前相当, 让其冲洗热水澡并休息一下。通过精神运动警觉测试来度量警觉状态和注意力, 去掉睡眠前后有较大改变的被试。重测流程与学习后测试类似但不呈现正确答案。
程序性记忆任务采用手指序列运动任务, 要求被试用非利手完成包含5个数字的敲击任务, 5个数字序列如4-2-3-1-4在屏幕上呈现30 s。要求被试又快又准确的进行按键并重复尽可能多的次数。实现流程:第一天临睡前的晚上 23:00左右对被试进行 12组的训练, 每组持续 30s, 间隔时间30 s, 将最后3组的平均成绩作为睡前行为得分。每组成绩根据正确完成的序列数和出现错误的数量进行评分。睡眠后测试在第二天凌晨 3:00左右进行。为保证被试的状态和睡眠前相当, 让其冲洗热水澡并休息一下。首先进行一次练习,随后是3组30 s的测试, 每组间隔时间为30 s。工作记忆的影响通过在屏幕上呈现刺激序列进行排除, 为杜绝其他干扰因素在30 s按键过程不提供任何反馈。
2.2 睡眠重激活
本部分在物体−位置匹配任务中加入物体对应的声音刺激, 并在记忆习得后的不同时间节点(2小时内、2天和6个月)的SWS睡眠期进行记忆激活。包括两个目标:第一, 采用时间窗法考察睡眠过程默认网络的动态变化, 将其与记忆编码过程进行对比, 探讨重激活中是否存在脑连接的重现, 结合睡眠后测成绩分析网络短时的连接强度起伏和行为准确率的关系; 第二, 在记忆习得后的不同时间节点加入睡眠过程声音刺激, 对比睡眠过程记忆重激活的脑区, 分析记忆习得后记忆重激活的转移情况。通过本部分的研究一方面可以观察到睡眠重激活的动态过程, 另一方面可能揭示睡眠重激活在较长时间里的转移情况。睡眠过程重激活任务分为睡眠前学习、睡眠中重激活和睡眠后测试3个阶段。
睡眠前学习阶段:被试在晚上 21:00进入实验室, 进行精神运动警觉测试。随后进行物体−位置配对学习, 首先在棋盘格某一位置上呈现一个生活用品要求被试观看并记住物品及所在的位置。在练习阶段被试通过鼠标把棋盘格上的物品移动到正确位置。物品呈现的同时播放其特异的声音, 如闹钟呈现时播放叮当声。移动操作结束后呈现该物品正确的位置进行反馈。摆放位置和正确位置的距离如果小于一定范围则判断为正确匹配, 所有项目都正确后进行睡眠前基线测试。
睡眠中重激活:邻近23:00开始佩戴电极帽,进入磁共振腔体中睡眠2小时。整个过程进行EEG记录, 在观察到慢波出现后立即开启 fMRI扫描进行同步 EEG-fMRI记录, 并播放学习阶段物体匹配的声音, 音量与学习阶段相同。学习阶段所有项目的声音被随机分为AB两组, 只有A组在睡眠阶段得到播放, B组将成为后测的行为基线。
睡眠后记忆测试:由于本过程大概在凌晨3:00进行, 为保证被试的状态和睡眠前相当, 让其冲洗热水澡并休息一下。通过精神运动警觉测试来度量警觉状态和注意能力, 去掉睡眠前后有较大改变的被试。最后进行物体−位置匹配的记忆重测, 评估睡眠的记忆巩固效应。
2.3 睡眠与静息态
本部分采用序列反应时任务, 同时记录记忆习得后的不同时间节点(1小时内、2小时后和 5小时后)等量的1小时静息和小睡对离线记忆巩固的影响。包括 3个目标:第一, 采用时间窗法研究静息态和睡眠过程默认网络在连接强度、模块数量的波动性、以及典型连接模式上的异同, 探讨两种离线巩固机制的差异及其与正确率、反应时的关系; 第二, 计算海马、前额叶和感觉运动皮层信号的长时记忆性(即幂律指数), 对比外显学习后同一脑区在不同时间节点上表现出的活动规律。第三, 延续我们之前在fMRI功能连接和EEG节律关系的研究, 重点关注慢波振荡和默认网络脑区连接的关系, 尝试建立基于默认网络的睡眠依赖记忆巩固的新理论。通过本部分的研究, 一方面可以揭示静息态和睡眠过程离线记忆巩固的异同, 另一方面可能澄清在昼夜更迭过程中记忆得以强化的基本原理。下面对序列反应时任务和睡眠结合静息态的实验设计进行重点介绍。
序列反应时任务在屏幕上从左到右4个位置依次设定为 1到 4号位置, 每个位置对应一个按键。实验开始时告知被试将要进行的是反应时测试。按键顺序依次由屏幕给出, 每次呈现一个视觉刺激, 被试按相应键尽快予以反应, 该刺激随即消失并在400 ms后呈现下一个目标按键。我们采用了外显学习方式:告知被试如果刺激数字由
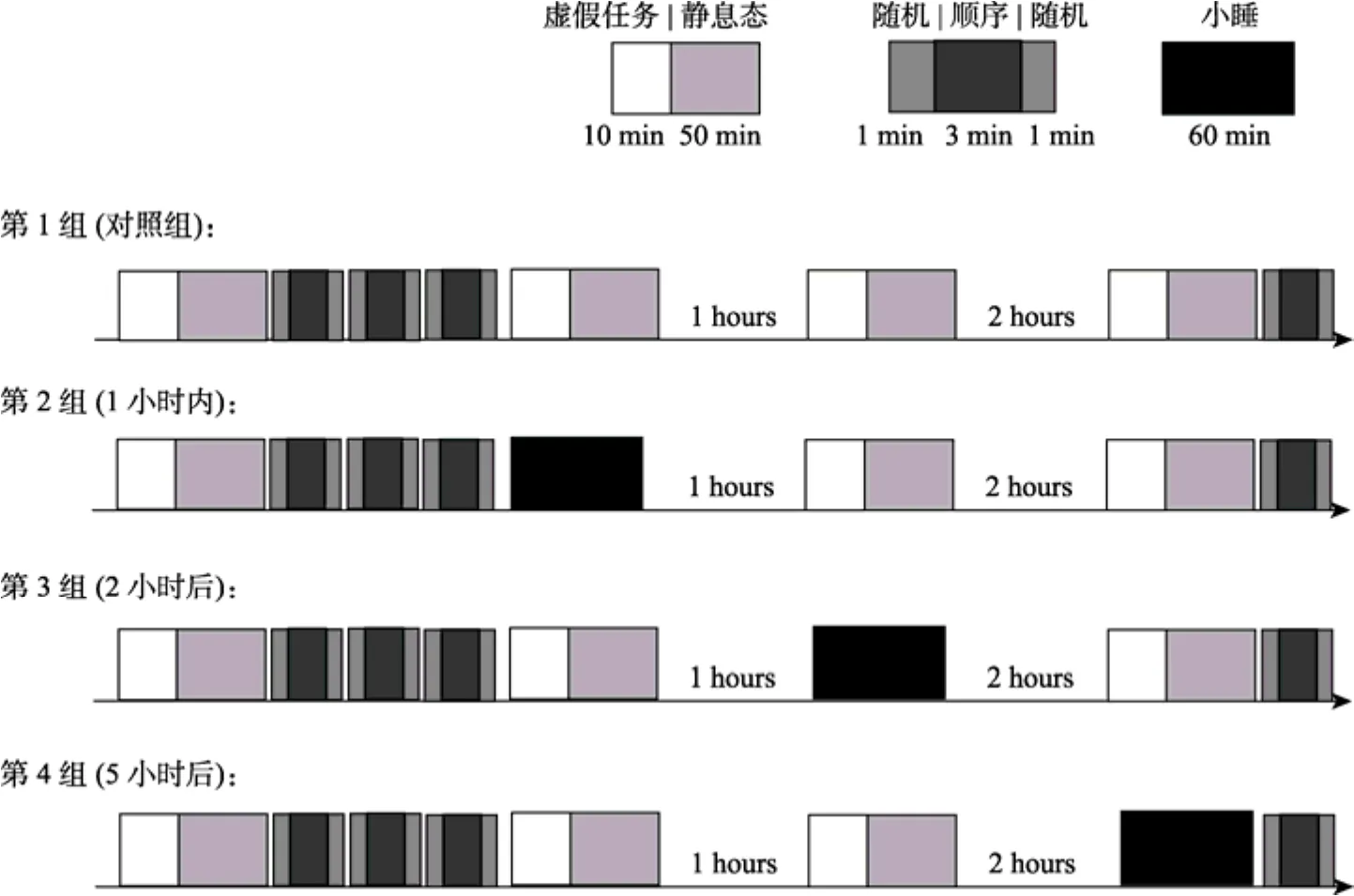
图1 静息态和睡眠状态离线巩固流程图
黑色变为蓝色, 接下来的按键序列是固定的, 如3-2-4-3-2-1-4-2-3-1, 但被试并不知道这个序列的长度和具体的内容。固定序列会多次重复, 并被包含在一个随机序列中。一共进行3个组块共重复固定序列 18次。整个任务约为 16分钟, 要求被试提前1小时到达扫描室。首先佩戴电极帽, 并根据指导语进行实验练习。被试对实验相当熟悉后开始正式的同步实验, 首先扫描静息态然后进入序列反应时任务, 结束后获得相应报酬, 整个实验的时间约为1小时。
睡眠结合静息态实验将学习后的被试随机分为静息态、1小时内、2小时后和5小时后小睡共4组, 如图1所示。对照组在3个时间点的任务均为睁眼盯着屏幕上的注视点“+”, 且要求不能进行系统性的思考。在静息态任务前还有一个模拟人体全身运动的动态光点刺激构成虚假任务(dummy task)。而其他3组在3个时间点允许进行1个小时的小睡。4组均进行同步EEG-fMRI采集,在采集间歇离开磁共振扫描间进入休息间, 可以补充淡水和进行阅读、上网等不剧烈的活动, 禁止摄入咖啡、茶等饮料。
3 理论建构与研究意义
睡眠和记忆都是大脑极为重要的功能, 大量研究表明睡眠有利于记忆的巩固(Lewis, 2014)。但目前关于记忆巩固在无创条件下神经活动的动态过程我们知之甚少, 白天清醒状态下脑的自发活动对记忆巩固的意义和作用还不明确, 记忆巩固的关键环路也有待澄清。神经成像技术的进步使得开展记忆巩固的动态脑网络研究变得可能。动态脑网络的主要着眼点是通过无创的成像手段以思维的速度描绘大脑活动。新兴的同步EEG-fMRI技术可以兼得高时间分辨率的电生理信号和高空间分辨率的血氧代谢信号(雷旭, 尧德中, 2014),为全面观察记忆巩固的动态过程, 厘清记忆巩固的关键回路创造了条件。
对比于传统的系统巩固理论, 本研究建立的基于默认网络的记忆巩固理论有3个突出特征。首先, 在空间分布上除了包括海马和内侧前额叶脑区, 后扣带回在记忆巩固中的作用是本理论的重要特色。后扣带回与多个内在连接网络具有广泛联系, 且在场景记忆检索中起到关键的整合作用。将后扣带回纳入到记忆巩固的研究中, 有利于突显默认网络的整体功能。其次, 在时间跨度上, 基于默认网络的记忆巩固理论有望统一白天的离线巩固和夜晚的睡眠依赖巩固这两个过程。默认网络在白天的任务态激活及走神过程中都有统一的活动模式, 而这一活动模式如何影响记忆巩固是本研究的重要方面。最后, 在研究手段上,本理论的建立有利于跨越基于任务态的记忆研究和基于静息态的记忆障碍研究这两者之间的鸿沟。目前基于静息态的记忆障碍研究由于其实验简单, 耗时短等优势, 正如火如荼的开展起来。基于默认网络的记忆巩固理论有利于从更为宏观, 更加注重任务和静息态结合的思路开展记忆研究。
借助于同步 EEG-fMRI, 本研究将解决以下关键问题:第一, 记忆巩固的动态电生理网络构建:纺锤波的细分和定位, 发展新的基于睡眠脑电的源定位、相位−幅度耦合分析等。第二, 记忆重激活脑区随时间的转移:发展基于时间窗和模块分析的动态分析方法, 对比较长时间范围内睡眠过程声音刺激激活脑区的转移, 揭示重激活中的脑连接模式。第三, 静息与睡眠条件离线记忆巩固的异同:提取静息态和睡眠过程默认网络的动态特性, 特别是海马、前额叶和感觉运动皮层的幂律指数, 对比各脑区在外显学习后不同时期的分布规律。第四, 基于默认网络的睡眠依赖记忆巩固:发展EEG和fMRI基于动态属性的融合方法, 尝试建立基于默认网络的睡眠依赖记忆巩固的新理论。
本研究的开展一方面可以发展新的脑功能探测的动态分析技术, 这些新技术势必将直接有利于解决认知神经科学、心理学和信息学相关领域科学问题; 另一方面对阐明睡眠依赖记忆巩固的神经机制具有深刻的理论意义, 并有可能为治疗学习记忆障碍提供全新的思路。
雷旭, 刘欢, 李红.(2013).睡眠的同步EEG-fMRI研究述评.心理科学进展, 21(3), 448−457.
雷旭, 尧德中.(2014).同步脑电-功能磁共振(EEG-fMRI)原理与技术.北京: 科学出版社.
Allen, E.A., Damaraju, E., Plis, S.M., Erhardt, E.B., Eichele,T., & Calhoun, V.D.(2014).Tracking whole-brain connectivity dynamics in the resting state.Cerebral Cortex, 24(3), 663−676.
Barrett, T.R., & Ekstrand, B.R.(1972).Effect of sleep on memory: III.Controlling for time-of-day effects.Journal of Experimental Psychology, 96(2), 321−327.
Colgin, L.L.(2013).Mechanisms and functions of theta rhythms.Annual Review of Neuroscience, 36, 295−312.
Debener, S., Ullsperger, M., Siegel, M., & Engel, A.K.(2006).Single-trial EEG-fMRI reveals the dynamics of cognitive function.Trends in Cognitive Science, 10(12),558−563.
Diekelmann, S., & Born, J.(2010).The memory function of sleep.Nature Reviews Neuroscience, 11(2), 114−126.
Diekelmann, S., Büchel, C., Born, J., & Rasch, B.(2011).Labile or stable: Opposing consequences for memory when reactivated during waking and sleep.Nature Neuroscience,14(3), 381−386.
Gais, S., Albouy, G., Boly, M., Dang-Vu, T.T., Darsaud, A.,Desseilles, M., … Peigneux, P.(2007).Sleep transforms the cerebral trace of declarative memories.Proceedings of the National Academy of Sciences of the United States of America, 104(47), 18778−18783.
Gregory, M.D., Agam, Y., Selvadurai, C., Nagy, A., Vangel,M., Tucker, M., … Manoach, D.S.(2014).Resting state connectivity immediately following learning correlates with subsequent sleep-dependent enhancement of motor task performance.Neuroimage, 102, 666−673.
He, B.J., Zempel, J.M., Snyder, A.Z., & Raichle, M.E.(2010).The temporal structures and functional significance of scale-free brain activity.Neuron, 66(3), 353−369.
American Academy of Sleep Medicine, & Iber, C.(2007).The AASM manual for the scoring of sleep and associated events: rules, terminology and technical specifications.American Academy of Sleep Medicine.
Karni, A., Meyer, G., Jezzard, P., Adams, M.M., Turner, R.,& Ungerleider, L.G.(1995).Functional MRI evidence for adult motor cortex plasticity during motor skill learning.Nature, 377(6545), 155−158.
Landmann, N., Kuhn, M., Piosczyk, H., Feige, B., Baglioni,C., Spiegelhalder, K., … Nissen, C.(2014).The reorganisation of memory during sleep.Sleep Medicine Reviews, 18(6), 531−541.
Langer, N., von Bastian, C.C., Wirz, H., Oberauer, K., &Jäncke, L.(2013).The effects of working memory training on functional brain network efficiency.Cortex, 49(9),2424−2438.
Lei, X., Wang, Y.L., Yuan, H., & Mantini, D.(2014).Neuronal oscillations and functional interactions between resting state networks: Effects of alcohol intoxication.Human Brain Mapping, 35(7), 3517−3528.
Lei, X., Xu, P., Luo, C., Zhao, J.P., Zhou, D., & Yao, D.Z.(2011).fMRI functional networks for EEG source imaging.Human Brain Mapping, 32, 1141−1160.
Lei, X., Zhao, Z.Y., & Chen, H.(2013).Extraversion is encoded by scale-free dynamics of default mode network.Neuroimage, 74, 52−57.
Lewis, S.(2014).Sleep and memory: To sleep and learn.Nature Reviews Neuroscience, 15(7), 429.
Marshall, L., & Born, J.(2007).The contribution of sleep to hippocampus-dependent memory consolidation.Trends in Cognitive Sciences, 11(10), 442−450.
Mölle, M., Bergmann, T.O., Marshall, L., & Born, J.(2011).Fast and slow spindles during the sleep slow oscillation:Disparate coalescence and engagement in memory processing.Sleep, 34(10), 1411−1421.
Rasch, B., & Born, J.(2007).Maintaining memories by reactivation.Current Opinion in Neurobiology, 17(6),698−703.
Sami, S., Robertson, E.M., & Miall, R.C.(2014).The time course of task-specific memory consolidation effects in resting state networks.Journal of Neuroscience, 34(11),3982−3992.
Takashima, A., Petersson, K.M., Rutters, F., Tendolkar, I.,Jensen, O., Zwarts, M.J., … Fernandez, G.(2006).Declarative memory consolidation in humans: A prospective functional magnetic resonance imaging study.Proceedings of the National Academy of Sciences of the United States of America, 103(3), 756−761.
Valderrama, M., Crépon, B., Botella-Soler, V., Martinerie, J.,Hasboun, D., Alvarado-Rojas, C.,...Le van Quyen, M.(2012).Human gamma oscillations during slow wave sleep.PLoS One, 7(4), e33477.
Wagner, T., Valero-Cabre, A., & Pascual-Leone, A.(2007).Noninvasive human brain stimulation.Annual Review of Biomedical Engineering, 9, 527−565.
Wang, Y.L., Liu, H., Hitchman, G., & Lei, X.(2015).Module number of default mode network: Inter-subject variability and effects of sleep deprivation.Brain Research, 1596, 69−78.
猜你喜欢
昆明医科大学学报(2021年2期)2021-03-29 07:42:30
中国生物医学工程学报(2019年4期)2019-07-16 08:04:02
现代电生理学杂志(2019年3期)2019-02-21 01:28:35
小天使·二年级语数英综合(2018年9期)2018-10-16 11:21:22
第二课堂(初中版)(2018年3期)2018-06-12 06:13:52
广西林业科学(2016年3期)2016-03-16 05:43:23
磁共振成像(2015年1期)2015-12-23 08:52:16
医学研究杂志(2015年5期)2015-06-10 06:43:26
四川生理科学杂志(2014年1期)2014-02-28 14:08:24
中国神经精神疾病杂志(2013年1期)2013-03-11 20:23:36
