
自闭症碎镜理论之迷思:缘起、问题与前景*
2016-02-01 00:17单春雷
心理科学进展 2016年6期
潘 威 陈 巍 汪 寅 单春雷
(1南京特殊教育师范学院康复科学学院, 南京 210038) (2浙江大学语言与认知研究中心, 杭州 310028)
(3绍兴文理学院心理学系, 绍兴 312000) (4纽约大学心理学院, 纽约 10003)
(5上海中医药大学康复医学院, 上海 201203)
1 引言
自闭症(autism), 又称孤独症, 现与阿斯伯格综合征(Asperger syndrome)、儿童瓦解综合征(childhood disintegrative disorder)以及未分类广泛性发展障碍(pervasive developmental disorder not otherwise specified)统称为自闭症谱系障碍(autism spectrum disorder)。作为病因未明的神经发育疾病(neurodevelopmental disorders), 其主要症状为在各种场合出现持久的社会沟通和交往障碍(涉及社会情感互动缺陷、非语言行为交流缺陷、发展维持和理解人际关系的缺陷), 伴随狭隘的兴趣、刻板的重复行为或活动模式(涉及刻板或重复躯体运动、高度受限的固定兴趣、感觉输入的过度反应或反应不足) (American Psychiatric Association,2013)。基于病情的严重程度, 自闭症患者在眼神交流、面部表情、社会兴趣、同伴关系以及社交或情感互动等方面呈现不同的状态。这些异常的状态与个体的模仿、心理理论(theory of mind) (揣摩他人心智的能力)、共情等社会认知能力(个体对外界状态的表征能力, 包括他人的心理状态或者行为) (Hohwy & Palmer, 2014)的缺损有关。自上世纪80年代以来, 研究者提出了许多理论解释自闭症患者的社会认知障碍, 诸如心盲理论(mindblindness theory)和共情系统化理论(empathizingsystemizing theory)等, 分别做出了自闭症患者心理理论缺失或者共情缺损的解释, 简化了对自闭症复杂行为的解释。然而, 行为学研究显示许多高功能自闭症患者具备显著或有限的心智化能力(mentalising), 只是其社会认知过程中潜在自动的直觉成分仍然是受损的, 而这种异常和损失又是自闭症的独有症状(Lai, Lombardo, & Baron-Cohen,2014), 这表明自闭症的心理理论缺失或共情缺损有可能只是这种直觉成分异常后的副产品。
碎镜理论(broken-mirror theory, BMT)正是从社会认知的直觉成分(潜在自动化的情绪或动作等的模仿与识别)受损层面解释自闭症症状机制的理论假说。作为人类镜像神经元系统(mirror neurons system, MNS)研究在自闭症领域中的推进与拓展, BMT提出了一整套关于MNS功能(社会认知的直觉成分)异常而导致自闭症症状的理论体系。BMT的支持者认为, MNS为自闭症提供了确定的神经机制, 未来的自闭症研究将从纷繁复杂、毫无头绪的境地走向可控的神经科学领域,解决自闭症难题指日可待。一时间, BMT引起了极大地关注。但是随着研究的深入, 越来越多的研究者发现, MNS作为社会认知神经机制的假说显得过于夸大或歪曲, 基于此而形成的BMT在解释自闭症患者的社会认知障碍上面临越来越多的质疑, 其理论的合理性有待确认(Southgate &Hamilton, 2008; Enticott et al., 2013; 陈巍, 汪寅,2015b)。甚至, 在Jarrett (2014)出版的新书《大脑的重大迷思》(Great Myths of the Brain)中, 有关“镜像神经元将我们塑造成人类”以及“碎镜诱发自闭症”的研究已经沦为了第 25位的迷思。一个在解释自闭症成因与表现上曾被赋以无限期待的崭新理论为何遭遇这般现实际遇?自闭症与镜像神经元之间究竟存在着怎样的关系?本文拟从BMT的发展路径出发, 系统回顾与梳理 BMT的内涵与证据, 通过澄清BMT面临的质疑及其证据,尝试对其未来发展做出理性展望。
2 碎镜理论的发展路径
2.1 镜像神经元与社会认知的关系
20世纪 90年代, 意大利帕尔玛大学的Rizzolatti, Fogassi和 Gallese等在猕猴属(基于豚尾猴[macaca nemistrina]和恒河猴[macaca mulatta]的研究)中意外发现了一些神经元, 当猴子自身做出一个动作(比如用手抓取物体)或者观察到研究人员进行同样动作的时候这些神经元出现了类似的激活反应。研究者根据其表现出的镜像特点将其命名为“镜像神经元”, 指出这种镜像的瞬间认知使得猴子仅仅通过观察就能够体验到外界的动作(Rizzolatti & Fabbri-Destro, 2010)。后续的研究表明, 镜像神经元(确切地说是其组成的神经系统)不仅传递运动控制指令(执行动作时激活), 也可让猴经由脑部神经活动达成理解动作和分辨意图的活动(在观察动作时激活; 在仅听到动作声音的条件下; 以及在传递动作意图的关键视觉线索被掩蔽的条件下出现与观察动作同样的激活反应),因此它也被视为模仿、动作认知发展的关键神经机制(Rizzolatti, Fogassi, & Gallese, 2006)。
目前, 基于实验伦理的限制无法采用侵入性技术获得人类存在镜像神经元的直接证据, 但是大量研究应用非侵入技术——例如脑电图(EEG)研究使用μ波(μ波是个体在进行随意运动或观察随意动作时出现的脑电波, 其抑制和激活与MNS的反应相反)、功能性磁共振成像(fMRI)研究采用血氧合度依赖(blood oxygenation level dependent contrast, BOLD)信号(神经元活动所引发之血液动力的改变)以及经颅磁刺激(TMS)通过微量电流刺激大脑等探查了人类的 MNS情况——发现人类大脑主要在顶下小叶(inferior parietal lobule)、腹侧前运动皮层(ventral premotor mortex)及额下回(inferior frontal gyrus)等区域出现镜像反应, 偶尔有研究发现顶上小叶(superior parietal lobule)和小脑(cerebel) (Molenberghs, Cunnington, & Mattingley,2012)、内侧颞叶(medial temporal lobe) (Mukamel,Ekstrom, Kaplan, Iacoboni, & Fried, 2010)等区域也有镜像属性。研究者认为镜像反应是一个系统化的功能, 独立具备该功能的神经元并不存在,因而将人类的镜像反应区域命名为MNS, 目前所指的人类 MNS主要包括:(1)包含脑岛(岛叶)(insula)和前额叶皮质(anterior mesial frontal cortex)的边缘镜像系统, 在理解和感受他人情绪时产生激活反应; (2)包含顶下小叶、腹侧前运动皮层以及额下回尾部的顶额镜像回路(parieto-frontal MNS circuit), 在观察或模仿单纯(不包含情绪成分)动作或行动时产生激活反应(Casartelli &Molteni, 2014)。
基于 MNS将外界动作或情绪映射到自身的能力, 这种连接自我与他人(self-other)关系的属性启发了研究者对其与社会认知关系的诸多研究。例如, Iacoboni等人(1999)研究发现MNS在模仿和动作理解中都发挥了作用, 他们在实验中要求被试观察和模仿手部动作, 然后根据提示再现该动作, 结果被试的MNS不仅在模仿手部动作时出现的激活, 而且在根据提示再现该动作时还出现了反应增强的现象, 研究者推测这种反应增强表明再现动作时MNS已经潜在理解了动作。随后,Iacoboni等人(2005)的研究发现MNS具有分辨意图的功能。他们为被试呈现了3组影片, 分别是:空白背景上一只手伸出来抓住一只杯子; 两个茶杯摆设的场景, 一个场景的摆设像是准备好让人享用下午茶, 另一个场景则好像用餐已毕等待收拾; 第三组影片是在不同场景下有一只手伸出来抓住杯子, 从而透露出动作的意图。结果显示被试对于具有清楚意图的动作反应最强烈, 而且在观察“饮用”或“收拾”的手部动作时, MNS出现不同的反应, 并且这两种场景下镜像神经元的激活程度都比看到空白背景的手拿杯子动作或是单纯只是观看场景的摆设来得更为强烈。Wicker等人(2003)的研究显示镜像神经元与共情也有关系。研究者让被试依次嗅闻刺激性气味或者观看别人吸入刺激性气味时面部表情的影片以体验恶心的情绪体验, 结果发现两种条件下被试的前脑岛(岛叶)区的反应相似, 而脑岛(岛叶)区也是人类MNS的重要组成部分。Gazzola, Aziz-Zadeh和 Keysers(2006)发现MNS在只听到动作声音的情况下就能够出现和执行动作时的同样地反应, 这表明MNS能够基于动作的相关属性实现对于动作的理解。
一些专家和学者认为, MNS的出现为我们理解社会行为提供了一个统一的神经机制视角, 也将掀起一场理解社会行为的革命(Iacoboni,2008)。当然, 即便 MNS的功能在当时并未得到澄清, 但它与社会认知的关系对于自闭症研究来讲, 确实也提供了崭新的思路。
2.2 碎镜理论的内涵与论据
自闭症的核心是社会认知的缺损和异常。这一结论早已得到了来自心理学、神经电生理以及脑功能成像等研究的证实。行为学的研究也显示自闭症患者普遍存在异常的社会认知特征, 如社会知觉功能受损、执行功能障碍以及异常的感知和信息处理等。
由于自闭症患者普遍存在社会认知异常,MNS又似乎是社会认知的神经机制, 这使得研究者意识到自闭症与 MNS二者之间可能存在某种关联, 并推测自闭症患者的MNS是异常的。来自EEG、fMRI和行为学研究的结论支持了这一想法。Oberman等人(2005)利用EEG记录自闭症患者和正常发育(typical development, TD)被试在观察和执行手部动作(例如抓握物品)时的 μ波情况,结果显示 TD被试在观察和执行动作时都有显著的μ波抑制, 而自闭症患者在执行动作时有μ波抑制, 但是在观察动作时则没有 μ波抑制。这表明自闭症患者执行动作的感觉运动皮层正常, 但是涉及动作模仿的MNS异常。Dapretto等人(2006)的fMRI研究记录了自闭症和TD被试在观察和模仿情绪时的反应, 结果显示自闭症被试的MNS活动存在异常, 主要表现在右侧额下回后部反应薄弱, 左额下回区域反应强烈。Martineau, Andersson,Barthélémy, Cottier和 Destrieux (2010)通过 fMRI研究自闭症和 TD被试在观察和执行手部动作时的表现, 同样显示自闭症被试的 MNS的反应异常。Pineda (2013)研究发现经由 μ波反应动作的生物反馈训练可以影响自闭症患者的 MNS反应,显示自闭症与MNS之间存在关联。更多支持BMT的实验证据详见表1。
结合自闭症、MNS与社会认知三者的关系,Williams, Whiten, Suddendorf和 Perrett (2001)、Ramachandran和 Oberma (2006)以及 Iacoboni和Dapretto (2006)等人提出了碎镜假说。“碎镜”是MNS功能异常的形象化表述, 主要观点是 MNS功能异常导致了自闭症患者的社会认知缺陷。个体在社会认知功能, 诸如模仿、共情以及动作理解等方面表现不一致继而导致自闭症患者之间症状的多样性。虽然MNS的研究反映它与模仿、共情和动作理解有极大的关联, 但是每位研究者对它们之间的关系有不同的看法, 因而BMT也同样出现了对于自闭症的不同解释。Hamilton (2013a)将 BMT归纳为 3个亚版本:模仿说(imitation version of BMT)、模拟说(simulation version of BMT)以及锁链说(chaining version of BMT)。
模仿说认为模仿是关系个体对他人动作表征的重要能力, 涉及了感知觉信息向运动信息的转换, 它的异常使得自闭症患者难以将视觉或听觉信息转换为动作, 限制了他们跟随和理解社交活动的能力, 因而自闭症是MNS异常而导致的模仿障碍, 继而影响了一系列社会认知能力的发展。模拟说从广义上拓展了 MNS的功能, 认为 MNS是所有对“自我与他人”的状态作出模拟反应的神经机制。自闭症患者不仅是模仿的障碍, 其共情或动作理解等都存在异常情况。Oberman等人(2005)指出MNS承载了个体内在的模拟机制, 是共情、心理理论、语言等正常发展所必须的生理基础。锁链说认为当我们在进行某种动作时, 实际上是在串连一系列的动作片段, 其顺序由我们的意图所决定。自闭症患者所表现出的社会认知缺陷是因为认知动作序列的MNS异常所致。这类MNS的异常致使个体在面对外界他人的动作或状态时无法理解完整的意图, 就像面对一个个静态的动作片段, 因而难以做出合适的反应, 表现为持续的社会交往和沟通障碍。Boria等人(2009)认为回答“为什么”类问题需要对于动作意图有整体的理解, 并利用行为研究考察了 TD和自闭症
儿童这一能力, 他们让被试观看一系列图片:包括中性背景中的电话; 手摸电话; 手抓话筒; 手抓电话机身, 然后提问“为什么这么做”以及“她在做什么”, 自闭症患者在回答“为什么”问题比回答“做什么”类问题上的表现显著较差, 这支持了锁链说的假设。

表1 支持BMT的研究
BMT的模仿说、模拟说以及锁链说从 MNS不同功能上丰富了自闭症研究的视角, 具有相当的吸引力。但是从另一角度来看, BMT的多样性似乎也表明人类 MNS和自闭症的研究仍然处于初级的摸索阶段。因此, 对于充满诱惑且又迷雾重重的BMT作出理性的反思, 考察其存在问题以明晰未来发展就显得尤为必要。
3 碎镜理论的现实境遇
按照传统的理解, BMT对MNS、社会认知和自闭症的相互关系给予了比较系统的透视。它打开了从社会认知的神经机制探索自闭症的通道,沿着这一通道我们对于自闭症有了更深入的理解。然而, 越来越多的研究者同时也意识到BMT所渲染的镜像神经元神话更像是一种执着的冒险(陈巍, 汪寅, 2015a)。迄今为止, 关于人类是否具有 MNS?MNS与社会认知存在怎样的具体关系?自闭症患者碎镜与否等诸多迷思仍在萦绕。种种现实问题无不反映出 BMT在极具吸引力的光环下似乎更多地面临着何去何从的尴尬境地。
3.1 人类MNS研究的内在缺陷
在审视MNS与BMT的问题之前, 我们有必要了解一些事实, 即人类的MNS研究本身存在内在缺陷。最严格意义上, 在这些问题尚未得到解决之前研究者甚至无法宣称人类具有 MNS或者说测量到了MNS的反应。
第一, 人类大脑中是否真正存在镜像神经元的问题仍值得商榷。目前仅有一例基于癫痫病人的单细胞研究显示人类的神经元可能具有镜像属性(Mukamel et al., 2010)。在猴类实验中, 研究发现的腹侧前运动皮层中具有镜像属性的细胞仅占该区细胞的 6%, 而且大部分猴类实验任务中猴子观察或执行抓握动作是经过先期训练的, 实验获得的数据并不能表明某一脑区较高强度的反应是单独的镜像神经元活动还是任务差异所导致(Kilner & Lemon, 2013), MNS究竟在动作观察或执行中发挥了多大作用同样有待确认(Perkins,Stokes, McGillivray, & Bittar, 2010)。这样的情况同样存在于人类MNS研究中。Lingnau, Gesierich和Caramazza (2009)的研究通过重复抑制原理(重复呈现某一刺激, 其相关的神经元会出现抑制反应)并未发现存在镜像反应的神经元。另一方面,许多学者一直倾向于对具有镜像属性的皮层回路进行进一步的研究, 但当前关于 MNS分布和功能的研究同样存在争议。Molenberghs,Cunnington和 Mattingley (2009)综合多项研究的元分析发现, 额下回区域并不能作为对应模仿功能发生的脑区, 广义镜像神经元区域仍然没有得到广泛的认同。
第二, 学术界尚难宣称获得了真实或精确的MNS反应。当前研究虽然采用了fMRI、EEG或者TMS等多种技术, 但是肌肉的兴奋水平、皮质脊髓的兴奋性、静息状态下的μ波等并不一定能匹配MNS的反应状态(Cattaneo et al., 2007)。μ波可能只是观察者自身感觉(触觉)的反应指标而不是观察外界动作的反应指标(Coll, Bird, Catmur, &Press, 2015)。Fitzgerald, Fountain和 Daskalakis(2006)指出MEG研究中使用的β波(β波反应了大脑初级运动皮层的兴奋性, 被当做MNS激活的一项指标)与被试实验前接受的磁刺激治疗、服用的药物、观察的认知或运动任务相关较大, 不具备实验的可重复性。而且, 当前匹配MNS反应的指标多是区域性的大批神经元反应, 研究者很难从这种复杂的区域神经系统的反应中区分出真实的MNS反应与其他运动神经元的反应。Shaw和Czekóová (2013)指EEG的研究无法明确神经系统失调或者兴奋的具体空间定位, 被试的感官经验和后期训练会影响脑电反应。Virji-Babul, Rose,Moiseeva和Makan (2012)的研究中就发现14~16个月大的婴儿观察其他婴儿爬行时的脑电要比行走时强烈的多, 也就是说脑电的变化存在经验依赖, 即我们当前通过EEG获得的所谓MNS的激活或者异常可能依赖于观察者是否具有被观察动作的经验, 这使得研究中容易出现经验差异导致结果不同的情况。此外, Bastiaansen等人(2011)的研究显示年龄同样影响MNS的反应。他们在自闭症患者观察面部表情的任务中发现, 不同年龄的自闭症患者在MNS反应上出现了显著差异, 成人被试较儿童具备更强的反应, 研究者推测MNS在18~55岁之间仍在持续的发展和完善。这些问题的存在显示研究者不仅难以明确区分研究中记录到反应, 而且对于人类MNS发展机制的理解还处在初级水平, 年龄、经验或者其他因素仍可能影响脑电或真实MNS的反应, 人类MNS研究的信度和效度有待提升。
3.2 MNS至多是社会认知的附带原因
即便我们忽略人类MNS研究的间接性, MNS与社会认知的关系似乎并非那么密切。首先, 社会认知所需的有效模仿涉及视觉分析、目的表征、动作选择等认知过程, 这些功能显然不是MNS所影响的简单的动作复制。Caramazza, Anzelotti,Strnad和Lingnau (2014)指出, 镜像神经元在动作理解中只参与了将动作转换为视觉表征的处理过程, 在此意义上并不能认为镜像神经元具有动作理解的功能。Janssen和Scherberger (2015)的分析显示顶叶皮层 PFG区和 V6A脑区等非镜像区在将客体信息由视觉转换为动作指令的过程中发挥了重要作用, 虽然目前尚不明了这些区域和MNS如何共同促进指向客体的动作的产生。
其次, MNS的主要功能也非是动作模仿或意图理解。Newman-Norlund, van Schie, van Zuijlen和Bekkering (2007)研究发现被试的MNS在进行辅助动作(complementary actions) (例如将手悬停在杯子上方这一动作, 它是举起手和握杯子或抓起杯子的中介辅助动作)的时候比观察模仿动作(imitative actions) (例如模仿研究者示范的抓杯子动作)的时候反应更加强烈, 因而指出 MNS主要影响的是各类动作的耦合即动作与动作如何连贯从而产生目的性行为而不是模仿或执行行为。Kana, Wadsworth和Travers (2011)综合分析了多个MNS与模仿关系的研究认为, MNS可能具有诸如注意力调整、自我与他人匹配或者目标认知等能够促进模仿的辅助功能, 但其主要功能并非模仿。此外, 动作理解往往涉及观察者的感觉系统和运动系统, 这一过程中更多的需要前运动区神经元的激活(Rizzolatti & Sinigaglia, 2010), 而镜像神经元关涉的动作理解仅仅是基于 MNS对于视觉输入的本能反应(Rizzolatti & Sinigaglia, 2010;Cook, Bird, Catmur, Press, & Heyes, 2014)。Kosonogov (2012)认为镜像神经元的主要功能不是认知动作目的或理解意图, 可能只是在大脑的某些结构对动作目地或意图进行分辨之后参与了一些步骤, 但并非重要角色。
再次, 镜像神经元的功能是联想学习(associative learning)的产物。近期的一系列收敛性证据1收敛性证据(converging evidence)是指在科学研究中当一系列实验始终支持某个假定的理论, 同时又能共同排除那些最主要的竞争性理论时, 这些实验证据就具有高度的收敛性。收敛性证据之所以重要, 是因为不同的方法很有可能涉及不同的假设, 而当一个假设能够通过众多基于不同假设的证伪检验时, 这些证据就构成了一个强有力的证据链, 并指向一个具有说服力的结论(Stanovich, 2012)。显示,MNS所具有的功能具有极强的可塑性, 这反映了这些功能不太可能是生物进化特异于社会认知的。例如, 动作理解的MNS反应可以经由训练改变(例如要求被试在观察到食指动作时以小指做反应, 多次训练后会MNS也会出现针对食指动作的非镜像反应) (Catmur, Walsh, & Heyes, 2007;Catmur, Mars, Rushworth, & Heyes, 2011)。来自婴儿EEG研究的证据也显示, 在婴儿大脑有关感觉运动耦合(perceptual-motor couplings)以及理解他人的这种耦合都依赖于相关视觉运动经验过程中的联想学习(de Klerk, Johnson, Heyes, & Southgate,2015)。据此, Hickok (2014)认为MNS与动作理解的原因无关, 只是在我们选择如何使用他人动作的过程中发挥了的作用, MNS的激活反应或许更应该是动作理解的结果。
由此可见, MNS与社会认知之间的关系尚未完全明朗。现存的实验结论并不支持有关两者关系的最初设想。正如Steinhorst和Funke (2014)总结的:“宣称MNS的重要社会功能只是在直觉上是可信的, 但这并不是衡量一个理论真实与否的科学标准”。目前, 已有学者明确指出 MNS至多是社会认知的附带原因(contributory cause)。即,MNS对于产生社会认知既非必要也非充分的, 虽然它的确有助于产生社会认知(Spaulding, 2013)。未来可以考虑进一步明确 MNS究竟在社会认知中扮演了何种角色, 以及它何种程度的异常会影响以及如何影响了社会认知。
3.3 MNS功能障碍不是自闭症的必要条件
从自闭症与MNS的关系来看, 近期很多研究显示自闭症患者的MNS是正常的, MNS功能障碍并不是导致自闭症的必要条件, 研究者认为前后研究矛盾的结果多是由于被试或者实验条件差异而造成的。
Raymaekers, Wiersema 和 Roeyers (2009)的研究记录了高功能自闭症和 TD儿童观察和执行手部动作的情况, 结果发现两组被试在两种情况下均出现显著的 μ波抑制现象, 且两组之间没有显著差异。Fan, Decety, Yang, Liu和Cheng (2010)的研究也发现自闭症与 TD被试在观察手部动作时均出现显著的 μ波抑制, 只是自闭症组无法模仿观察到的动作, 研究者认为自闭症患者的MNS正常, 可能只是功能受到抑制。Marsh和 Hamilton(2011)发现, TD和自闭症被试在观察合理(rational)和不合理(irrational) (以“合理”与“不合理”分别考察被试的动作理解能力和单纯的镜像反应)的手部动作时, 两组被试在MNS的左前内沟区域同样激活, TD被试同时激活了扣带回及其双侧皮质的神经回路。当观看不合理的手部动作时, 两组被试无显著差异。Sowden, Koehne, Catmur, Dziobek和Bird (2015)开展的行为学研究显示自闭症患者的自动模仿能力正常, 他们和匹配组在模仿手指举起的任务上表现并无差异, 但是在分别考察被试的自动模仿能力(例如, 在不同颜色背景下模仿手部动作, 以剔除相同的背景可能对模仿的促进)和空间兼容(spatial compatibility)对模仿行为影响(例如, 在相同背景下, 要求被试观察左手动作时用右手模仿, 以考察同样的空间背景能否促进被试的模仿)的实验中, 却发现自闭症患者的用时更短, 即自闭症并非模仿能力缺损, 反而更可能是一种异常强烈的自动模仿倾向所导致的疾病。由此, 许多研究者认为自闭症患者的MNS正常, 而且镜像反应与动作理解是不同脑区的功能, MNS似乎并不涉及动作理解, 与自闭症的社会认知异常和缺损关系不大。
此外, 如果自闭症的成因是MNS的缺损, 那么自闭症患者大脑的MNS应该有别于正常人。但是Toal等人(2010)研究了65位自闭症患者的大脑,结果发现自闭症患者的内侧颞叶梭状回和小脑比常人稍小, 额下回或顶下小叶等经典MNS区与常人并无差异。Duerden, Mak-Fan, Taylor和Roberts(2012)的研究分析了 22个自闭症研究的报告, 结果显示自闭症患者与正常人在前额叶、扣带回和脑岛(岛叶)存在大小差异, 但额下回或顶下小叶区等镜像区域并无结构或功能的区别。Hamilton(2013a)分析了25个利用fMRI、EEG或眼球追踪技术研究自闭症患者MNS功能的研究报告, 指出并无确凿的证据支持BMT关于自闭症患者MNS异常说法, 并且指出基于BMT的干预方法也没有效果。更多质疑BMT的实验证据详见表2。
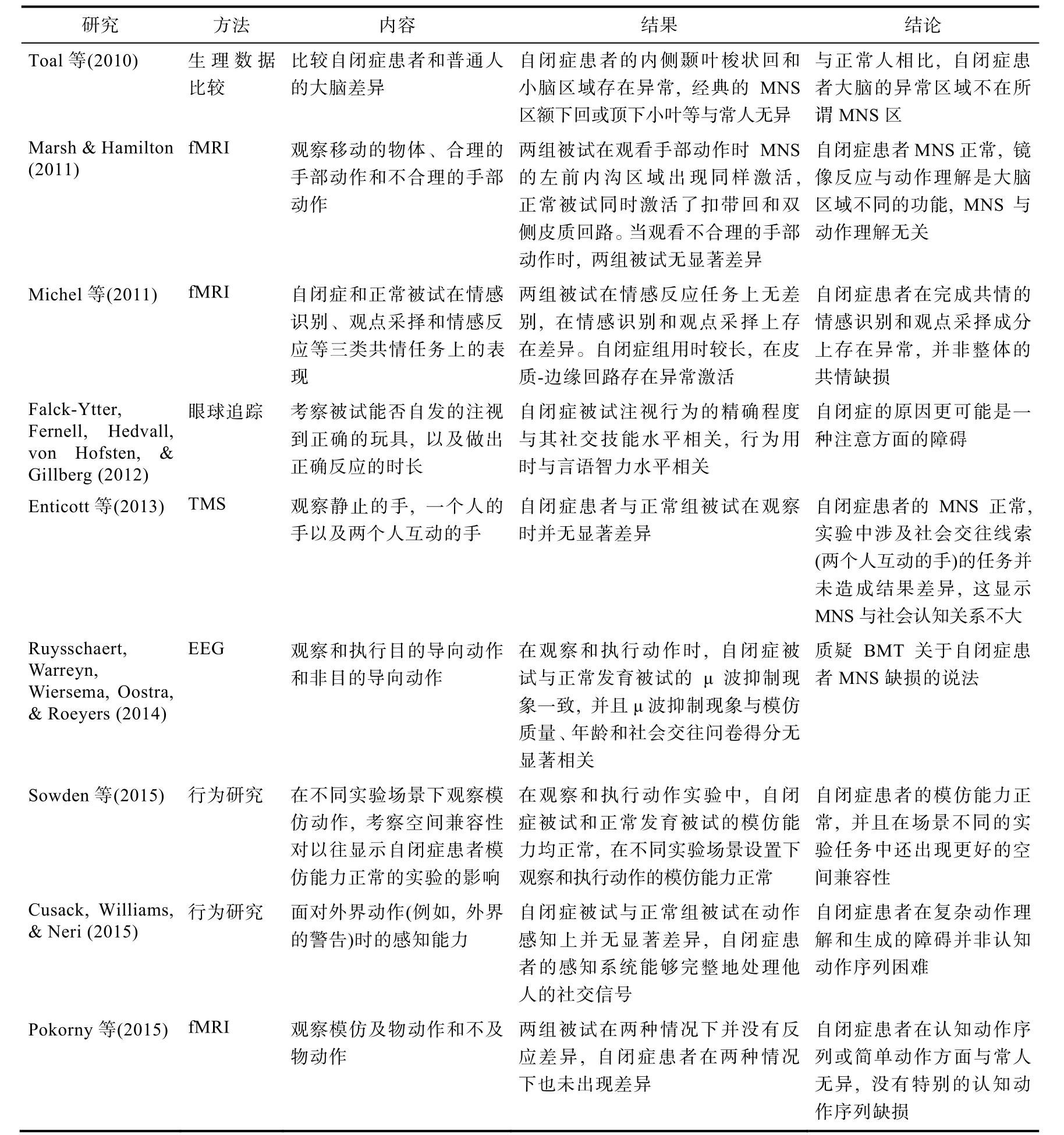
续表2
综上, MNS功能障碍不是自闭症的必要条件。新近研究显示自闭症患者的MNS正常, 自闭症患者与正常人的大脑 MNS区域也没有显著差异。早期关于自闭症患者MNS异常的说法可能是MNS的功能受到抑制或者是实验任务与被试的差异导致了不同的结果。研究者更倾向认为自闭症患者MNS是正常的, 只是其功能由于自闭症的继发原因受到抑制。
3.4 碎镜理论各版本的观点缺乏依据
BMT的问题不仅在于MNS与社会认知、自闭症与MNS的关系模糊, 其各种理论版本亦缺乏说服力。
从BMT的主要观点看, 模仿说认为MNS是模仿的重要基础, 自闭症患者表现出的模仿障碍是MNS功能异常的表现。然而, 当前诸多研究显示 MNS与模仿的关系不大, 而即便有研究表示MNS应该参与了模仿的重要环节, 但基于自闭症患者的研究发现他们的模仿能力并未受影响。Smith和 Bryson (2007)研究了自闭症患者在社交手势(如对人竖大拇指)和哑剧手势(如握拳)方面的模仿能力, 结果对比非自闭症的发展迟滞和TD被试, 在明确的要求下, 自闭症患者能够完成大致准确的模仿, 但是难以完成对手势的命名和按照实验者的语言指令再现手势。这说明自闭症患者的模仿能力是正常的, 他们的问题可能在于不知道何时模仿以及模仿什么。
就模拟说而言, 也有研究显示自闭症患者在动作理解和共情上并没有困难。Michel等人(2011)的研究考察了自闭症患者和正常被试在情感识别、观点采择和情感反应等三类共情任务上的表现, 发现自闭症患者与正常被试在情感反应上并无差别, 但是在情感识别和换位思考任务上表现较差, fMRI记录显示了两组被试在皮质- 边缘区域(cortico-limbic)的反应差异。这显示自闭症患者似乎并非整体的共情(认知共情和情感共情)异常,可能只是有利于完成共情的情感识别和换位思考方面存在缺损, 并非模拟“自我与他人”状态的损伤。
BMT的锁链说认为, 自闭症患者认知动作序列的MNS异常, 所以在复杂的动作理解和生成上存在障碍。但是Spunt, Falk和Lieberman (2010)的研究却显示在“为什么”类问题(例如实验者做出喝茶行为, 提问被试该实验者“如何做(用杯子喝)”和“为什么做(为了保持清醒)”)上表现较差不是动作理解的障碍, 而是由于无法表征他人的心理状态。因而, Boria等(2009)发现自闭症患者在“为什么”类问题表现较差并不能说明他们无法认知动作序列。Cusack等(2015)在研究中调查了自闭症患者的动作感知能力, 结果发现自闭症被试与TD组被试在动作感知(例如, 外界的警告)上并无显著差异, 自闭症患者的感知系统能够完整地处理他人的社交信号, 他们在复杂动作理解或生成上的异常表现可能是由于不知何时做出反应以及如何做出反应, 并不符合锁链说的假设。Pokorny等人(2015)的研究考察了自闭症患者和 TD成人在观察模仿及物动作(主体指向客体的动作, 例如伸手越过书拿杯子)和不及物动作(例如伸手越过书)时 MNS的反应情况, 结果发现二者在两种情况下并没有反应差异, 而且自闭症患者在两种情况下也未出现差异, 这表明自闭症患者在认知动作序列或简单动作方面与常人无异, 没有特别的认知动作序列缺损。
从上述的这些事实可以发现, BMT几乎是在一个没有牢固根基的情况下突现的一座美丽的空中楼阁。即便我们忽略人类MNS研究的先天缺陷,关于MNS与社会认知的具体关系、自闭症患者的MNS究竟是否异常, 或者自闭症各症状之间的关联如何受到 MNS的影响等问题的答案依然是模糊的。或许, 这也是我们整理几近揭开自闭症谜团的兴奋之情, 拨开BMT迷思, 走向新征程的时候了。
4 碎镜理论及自闭症研究的未来发展
镜像神经元的发现至今已20多年(di Pellegrino,Fadiga, Fogassi, Gallese, & Rizzolatti, 1992), 而BMT的提出也已将近 10年(Ramachandran &Oberman, 2006)。这10年正好是镜像神经元从神经神话(neuromyth)降到理性思考的10年。由于科学界对镜像神经元功能至今仍然没有统一意见,各个领域对其研究及相关争论还会持续下去。同时,随着自闭症研究对象的变化, 镜像神经元的社会响应学说的提出, 双个体社交范式、第二人称神经科学范式与计算精神病学的兴起, 我们相信镜像神经元与自闭症的真正关系将被逐步阐明。
4.1 新研究对象对于碎镜理论的验证
近些年来, 自闭症的发病率提高了30倍, 例如美国从1980年的0.05%提升到2014年的1.5%。有研究表明自闭症发病率的猛增主要是由于诊断手段的变化(Hansen, Schendel, & Parner, 2015)。旧的诊断手段(DSM-V之前)详细规定了界定自闭症患者与正常个体的标准以及轻度和重度自闭症的界限。然而随着自闭症流行病学的研究以及遗传学对自闭症相关基因的分析(Chakrabarti et al.,2009; Robinson et al., 2011), 人们发现自闭症在总人群中的发生概率以及严重程度呈连续正态分布, 并且某些人格特质也与自闭症症状高度重叠(Baron-Cohen, 2009; Ronald & Hoekstra, 2011)。如果把临床诊断中用于测量自闭症特征(autistic traits)的自闭症问卷(autism spectrum quotient, AQ)施用在总人群, 那么 93%的正常个体都会正态分布于32分以下(AQ总分为50分), 而99%的自闭症获诊个体则正态分布于 32分以上(Baron-Cohen,Wheelwright, Skinner, Martin, & Clubley, 2001)。考虑到自闭症患者的高度异质性(heterogeneity) (即个体之间智力、社会成长背景、缺损社会认知功能的数量和程度差异极大), 之前研究中许多相互矛盾的实验结果极有可能是异质性所致(Bölte,Poustka, & Constantino, 2008)。为了避免异质性对于研究结果的干扰, 一些学者建议可以(跳过自闭症患者)直接研究正常个体中自闭症特征和镜像神经元活动的相关关系, 从而来间接验证BMT:比如让正常个体完成一些测量自闭症特征的问卷,然后比较是否自闭症特征较高的个体其 MNS的活动较自闭症特征低的个体更弱。考虑到目前该类型的研究还不多, 并且一半支持(Puzzo, Cooper,Cantarella, Fitzgerald, & Russo, 2013; Cooper,Simpson, Till, Simmons, & Puzzo, 2013)一半反对BMT (Haffey, Press, O'Connell, & Chakrabarti,2013; Sims, Neufeld, Johnstone, & Chakrabarti,2014), 将来我们需要更多在正常个体中的研究来验证BMT。
4.2 新理论对于镜像神经元功能的探索
虽然本文第3部分的大量科学证据基本否定了 BMT的观点(即镜像神经元受损是自闭症的主要病因), 但是人们并不能完全排除自闭症患者某些细微社会认知功能的缺损是镜像神经元异常活动所致。因此, 在最终消除 BMT“神话”之前, 我们还需尽可能阐明镜像神经元的真正功能。之前的理论主要把镜像神经元和动作理解、模仿等功能联系在一起, 但本文的 3.2部分详细例证了镜像神经元既不是动作理解的充分必要条件, 其活动特征也与动作模仿所需的精确复制性相违背。那么, 除动作理解和模仿外, 镜像神经元还可能参与其他什么社会认知功能?
Hamilton (2013b)最近提出了一个新理论:镜像神经元可能主要参与社会响应功能(social responding)。受辅助动作(complementary actions)实验的启发(Newman-Norlund et al., 2007), Hamilton认为镜像神经元不仅仅在被动地分析所观察到的动作信息, 其更重要的功能是从自身运动皮层中激发那些能回应该目标动作的运动表征, 从而为接下去可能做出的社会回应行为做准备。为了便于理解该功能, 我们可以想象以下情景:当你的朋友在饭桌上把啤酒杯递给你的时候, 镜像神经元可能不仅仅帮你理解那是一个“握着啤酒杯的动作”, 其更重要的是诱发你接下去做出合适的社会反应, 即“张开手掌做出接杯子的动作”。在这里, 由于啤酒杯的两侧不一样(一侧是柄, 另一侧是宽大的杯面), 你看到他人的动作(细握杯柄)和你自身回应的动作(张开手掌去握杯面)会稍有不同; 但如果这是一个普通的透明玻璃杯(圆柱状无柄), 则观察和回应的动作完全相同的。这种允许镜像神经元在动作观察和执行时编码一致和非一致的表征很好地解释了为什么只有 1/3的镜像神经元是“严格一致的镜像神经元” (strictlycongruent mirror neurons), 而2/3的镜像神经元是“宽泛一致的镜像神经元” (broadly-congruent mirror neurons) (Rizzolatti & Craighero, 2004; Kilner &Lemon, 2013)。
虽然该理论才刚刚提出, 并且还没有实验证据直接论证, 但从逻辑上来说, 镜像神经元的一些重要特征都支持社会响应理论, 如推测性镜像神经元(predictive mirror neurons)或逻辑相关的镜像神经元(logically-related mirror neurons) (Keysers& Gazzola, 2014), 以及联想学习性(几乎所有人类社会响应动作是通过后天学习的产物) (Cook et al., 2014)。考虑到自闭症患者的日常行为中严重缺乏社会响应动作, 这极有可能是镜像神经元与自闭症联系最紧密的环节。
4.3 新研究范式对于镜像神经元以及自闭症研究的推动
受到还原主义(redunctionism)的影响, 心理学研究往往聚焦于单一的外界刺激和过度细化的认知过程。这种影响在镜像神经元的研究中也颇为明显。虽然日常生活中人类的社会行为丰富多彩, 镜像神经元的实验主要还是围绕在简单的个体动作上(比如观察一只手在拿杯子, 或者一张脸在笑)。我们认为, 运用双个体社交范式(dyadic social interaction paradigm)来研究镜像神经元是否参与复杂的群体动作理解可能是将来的热点(Wang & Quadflieg, 2015)。举两个例子:首先, 当我们看到一些社会情景, 比如一位学生拉着另外一位快要摔倒的学生的胳臂时, 镜像神经元是如何理解这个群体行为?这两个学生各自的行为极为不同, 我们的镜像神经元是如何用唯一的运动皮层来同时模拟这两种不同的肢体动作?镜像神经元又如何区分这个动作是亲社会性的(如扶起摔倒的同学)而不是反社会性的(两个同学打架)?另外一个例子, 当你看到两个小朋友打闹以及他们面部表情的时候, 我们是如何理解以下 4种微妙的行为意图:快乐玩耍(两个小朋友都在笑), 真正打架(两个小朋友都在生气), 一方欺凌另一方(推搡的小朋友在笑, 被推搡的小朋友生气), 一方挑衅而另一方被激怒(推搡的小朋友在生气, 被推搡的小朋友在笑)。理解这些复杂的群体行为涉及到对于两个个体动作(或面部表情)的宏观解读:仅仅明白单一个体动作(比如拉手臂、摔倒、哭、笑)不足以让我们完全理解整个动作的意义。在这里, 简单的镜像神经元模拟说似乎无法帮助个体理解复杂的群体动作(因为个体无法同时模拟两种不同的全身动作), 所以极有可能需要其他的认知过程或神经网络去参与整合多种单一动作的信息。考虑到自闭症患者缺乏对于事物 “格式塔式”的宏观理解(参考弱中心统合理论), 我们可以想象自闭症患者在日常生活中遇到的动作理解困难很有可能不是镜像神经元对于单一动作理解的问题, 而是某些负责整合多种动作信息并对其进行格式塔式解读的认知过程或神经网络出了问题(即在动作理解方面的“见木不见林”) (Simmons et al., 2009, Bölte, Holtmann, Poustka, Scheurich, &Schmidt, 2007)。我们相信将来会有不少研究运用这种新型多个体社交范式去冲击 BMT对于动作理解缺损的解释。
此外, 第二人称神经科学范式(second-person neuroscience paradigm)可能是未来推动镜像神经元和自闭症研究的另一个重要方向 (Schilbach,2015)。社会神经科学目前主要采取的是第三人称式的研究方法(third-person neuroscience), 即被试在核磁共振仪中被动地观察实验者所提供的各种社会刺激(如动作、面部表情、社会场景)。这种像看电影似地感知社会刺激而不是像日常生活中那样主动参与到社会交流中的范式在近些年来饱受诟病(Schilbach et al., 2013)。因为无论从神经现象学角度还是认知科学角度, 大脑在经历这两种不同范式时的状态极为不同(Schilbach, 2014):前者被称为离线社会认知(off-line social cognition)而后者则是被称为在线社会认知(online social cognition)。这两者的关系就如同认知主义(Cognitivism)和生成主义(Enactivism)般微妙:前者认为大脑主要负责信息处理从而帮助个体更好认识理解这个世界,而后者强调大脑服务于个体更好地与周围的环境互动从而更好适应这个世界(Cisek & Kalaska,2010)。让我们回到上面提到过的递啤酒杯的例子:当你看到你的朋友在饭桌上把啤酒杯递给你的时候, 你的大脑很有可能已经准备启动接啤酒杯的动作; 但是当你回家在沙发上观看用手机录下的一模一样的递啤酒杯的动作时, 你的大脑不太可能激活任何和接啤酒杯相关的动作, 因为你是以第三人称被动地旁观递啤酒的动作, 而不是以第二人称设身处地地作为递啤酒动作的对象。换而言之, 你身处的环境决定了你大脑会对相同的视觉信息做出不同的生理回应。借助于近些年来实验仪器设备的发展, 目前已经有不少研究采取了第二人称神经科学范式, 如用基于眼动仪的实时交互社交实验(eye-tracker based real-time social interaction)以及基于虚拟现实技术的社会互动实验(immersive virtual reality technology)(Pfeiffer, Vogeley, & Schilbach, 2013), 而这些实验也表明不同的脑区以及不同的神经网络分别参与到第二人称和第三人称范式的实验中(Schilbach et al., 2013)。
因此, 结合前面讨论到的镜像神经元社会响应理论(Hamilton, 2013b), 将来对于镜像神经元的研究应该更多的采取第二人称式的范式, 即在交互情景下(reciprocal/ interactive social contexts)监测MNS对于观察到的动作的回应。对于自闭症研究来说, 考虑到之前研究发现不少患者在完成某些离线社会任务时(如仅仅通过观察面部图片来识别情绪)与正常被试无显著差异, 但是到了日常生活中面对类似的刺激仍有困难(如自闭症患者可以区分微笑的脸和愤怒的脸, 但是他们不知道行为上该如何回应这两种表情), 将来研究应该多采取第二人称的研究方式来细化自闭症患者在在线社会认知以及离线社会认知上是否存在缺损(Klin, Jones, Schultz, & Volkmar, 2003; Schilbach,Eickhoff, Cieslik, Kuzmanovic, & Vogeley, 2012)。
4.4 计算精神病学(computational psychiatry)的兴起
受从吝啬律(law of parsimony)的影响, 科学研究往往偏爱用简单的理论去解释精神疾病的病因。BMT之所以在提出时就影响巨大, 其主要原因是能解释自闭症中几乎所有社会功能的缺陷(如动作理解、共情、模仿、读心、语言发展障碍)。然而, BMT这种全面性同时也变成了其劣势:由于自闭症患者的异质性太强, 不同社会功能的受损程度也大不相同, 仅仅基于某个统一的神经基础的受损自然无法去解释这样的复杂性。
计算精神病学的兴起或许可能帮助我们克服这些单一理论的缺陷, 从而更好的解密自闭症的病因(Montague, Dolan, Friston, & Dayan, 2012;Wang & Krystal, 2014)。计算精神病学通过设计神经网络模型、模拟数据运算以及预测患者临床表现, 它可以比较以及结合多种现存的理论。例如,对于自闭症来说, BMT可以和心盲理论相结合:通过建模 MNS脑区和心智化脑区(mentalizing system)之间的神经通路和交互作用, 从而推测正常个体中两个系统如何共同运作或者弥补互作来实现一些社会认知功能, 然后再在自闭症患者身上进行功能性脑成像实验, 看看这些不同神经网络的交互作用是否出现了问题。这样的研究思路可能更接近事实, 因为大脑的运作和社会功能的实现都是由负责不同微小认知功能的脑网络进行协同合作的结果。MNS可能负责对于社会动作信息的初步分析和情绪的初步共鸣, 之后复杂的社会推测功能则需要心智化脑区对于之前信息更加深入地处理。如果这两个神经网络自身都没有问题, 但是相互的协作出了问题, 则有可能最终表现出一些复杂的行 为学特征或临床表现:比如不少自闭症患者动作理解的功能无障碍(即MNS功能正常), 在外显的读心过程(explicit theory of mind task)也没问题(即心智化脑区也功能正常),但是在内隐的读心过程(implicit theory of mind task)发生了问题, 这很有可能是MNS产生的初步动作理解产物无法自动被心智化脑区所分析和利用。总而言之, 未来计算精神病学可以验证和整合之前大量的理论, 其发展将会大大加深我们对于自闭症病因的理解。
陈巍, 汪寅. (2015a). 镜像神经元是认知科学的“圣杯”吗?
心理科学, 38(1), 237–242.
陈巍, 汪寅. (2015b). 基于镜像神经元的教育: 新“神经神话”的诞生?教育研究, 36(2), 92–101.
American Psychiatric Association. (2013).Diagnostic and statistical manual of mental disorders(DSM-5)(5th ed.).Arlington, VA: American Psychiatric Publishing.
Baron-Cohen, S. (2009). Autism: The empathizing-systemizing(E-S) theory.Annals of the New York Academy of Sciences,1156, 68–80.
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martin, J., &Clubley, E. (2001). The autism spectrum quotient (AQ):Evidence from Asperger syndrome/high functioning autism,males and females, scientists and mathematicians.Journal of Autism and Developmental Disorders, 31(1), 5–17.
Bastiaansen, J. A., Thioux, M., Nanetti, L., van der Gaag, C.,Ketelaars, C., Minderaa, R., & Keysers, C. (2011).Age-related increase in inferior frontal gyrus activity and social functioning in autism spectrum disorder.Biological Psychiatry, 69(9), 832–838.
Bernier, R., Dawson, G., Webb, S., & Murias, M. (2007).EEG mu rhythm and imitation impairments in individuals with autism spectrum disorder.Brain and Cognition, 64(3),228–237.
Bölte, S., Holtmann, M., Poustka, F., Scheurich, A., &Schmidt L. (2007). Gestalt perception and local–global processing in high-functioning autism.Journal of Autism and Developmental Disorders, 37(8), 1493–1504.
Bölte, S., Poustka, F., & Constantino, J. N. (2008). Assessing autistic traits: Cross-cultural validation of the social responsiveness scale (SRS).Autism Research, 1(6), 354–363.
Boria, S., Fabbri-Destro, M., Cattaneo, L., Sparaci, L.,Sinigagli, C., Santelli, E., … Rizzolatti, G. (2009).Intention understanding in autism.PLoS One, 4(5), e5596.
Caramazza, A., Anzelotti, S., Strnad, L., & Lingnau, A.(2014). Embodied cognition and mirror neurons: A critical assessment.Annual Review of Neuroscience, 37, 1–15.
Casartelli, L., & Molteni, M. (2014). Where there is a goal,there is a way: What, why and how the parieto-frontal mirror network can mediate imitative behaviours.Neuroscience and Biobehavioral Reviews,47, 177–193.
Catmur, C., Walsh, V., & Heyes, C. (2007). Sensorimotor learning configures the human mirror system.Current Biology, 17(17), 1527–1531.
Catmur, C., Mars, R. B., Rushworth, M. F., & Heyes, C.(2011). Making mirrors: premotor cortex stimulation enhances mirror and counter-mirror motor facilitation.Journal of Cognitive Neuroscience, 23, 2352–2362.
Cattaneo, L., Fabbri-Destro, M., Boria, S., Pieraccini, C.,Monti, A., Cossu, G., & Rizzolatti G. (2007). Impairment of actions chains in autism and its possible role in intention understanding.Proceedings of the National Academy of Sciencesof the United States of America, 104,17825–17830.
Chakrabarti, B., Dudbridge, F., Kent, L., Wheelwright, S.,Hill-Cawthorne, G., Allison, C., … Baron-Cohen, S.(2009). Genes related to sex steroids, neural growth, and social-emotional behavior are associated with autistic traits, empathy, and Asperger syndrome.Autism Research,2(3), 157–177.
Chien, H. Y., Gau, S. S., Hsu, Y. C., Chen, Y. J., Lo, Y. C.,Shih, Y. C., & Tseng, W. Y. (2015). Altered cortical thickness and tract integrity of the mirror neuron system and associated social communication in autism spectrum disorder.Autism Research, 8(6), 694–708.
Cisek, P., & Kalaska, J. F. (2010). Neural mechanisms for interacting with a world full of action choices.Annual Review of Neurosciences, 33, 269–298.
Coll, M. P., Bird, G., Catmur, C., & Press, C. (2015).Cross-modal repetition effects in the mu rhythm indicate tactile mirroring during action observation.Cortex, 63,121–131.
Cook, R., Bird, G., Catmur, C., Press, C., & Heyes, C. M.(2014). Mirror neurons: From origin to function.Behavioral and Brain Sciences, 37(2), 177–192.
Cooper, N. R., Simpson, A., Till, A., Simmons, K., & Puzzo,I. (2013). Beta event-related desynchronization as an index of individual differences in processing human facial expression: Further investigations of autistic traits in typically developing adults.Frontiers in Human Neuroscience,7, 159.
Cusack, J. P., Williams, J. H. G., & Neri, P. (2015). Action perception is intact in autism spectrum disorder.The Journal of Neuroscience, 35(5), 1849–1857.
Dapretto, M., Davies, M. S., Pfeifer, J. H., Scott, A. A.,Sigman, M., Bookheimer, S. Y., & Iacoboni, M. (2006).Understanding emotions in others: Mirror neuron dysfunction in children with autism spectrum disorders.Nature Neuroscience, 9, 28–30.
de Klerk, C. C. J. M., Johnson, M. H., Heyes, C. M., &Southgate, V. (2015). Baby steps: Investigating the development of perceptual-motor couplings in infancy.Developmental Science, 18, 270–280.
di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., &Rizzolatti, G. (1992). Understanding motor events: A neurophysiological study.Experimental Brain Research,91(1), 176–180.
Dinstein, I., Thomas, C., Humphreys, K., Minshew, N.,Behrmann, M., & Heeger, D. J. (2010). Normal movement selectivity in autism.Neuron, 66, 461–469.
Duerden, E. G., Mak-Fan, K. M., Taylor, M. J., & Roberts, S.W. (2012). Regional differences in grey and white matter in children and adults with autism spectrum disorders: An activation likelihood estimate (ALE) meta-analysis.Autism Research, 5(1), 49–66.
Enticott, P. G., Kennedy, H. A., Rinehart, N. J., Tonge, B. J.,Bradshaw, J. L., Taffe, J. R., … Fitzgerald, P. B. (2012).Mirror neuron activity associated with social impairments but not age in autism spectrum disorder.Biological Psychiatry, 71(5), 427–433.
Enticott, P. G., Kennedy, H. A., Rinehart, N. J., Bradshaw, J.L., Tonge, B. J., Daskalakis, Z. J., & Fitzgerald, P. B.(2013). Interpersonal motor resonance in autism spectrum disorder: Evidence against a global “mirror system”deficit.Frontiers in Human Neuroscience, 7, 218.
Falck-Ytter, T., Fernell, E., Hedvall, Å. L., von Hofsten, C.,& Gillberg, C. (2012). Gaze performance in children with autism spectrum disorder when observing communicative actions.Journal of Autism and Developmental Disorders,42(10), 2236–2245.
Fan, Y. T., Decety, J., Yang, C. Y., Liu, J. L., & Cheng, Y. W.(2010). Unbroken mirror neurons in autism spectrum disorders.The Journal of Child Psychology and Psychiatry,51(9), 981–988.
Fitzgerald, P. B., Fountain, S., & Daskalakis, Z. J. (2006). A comprehensive review of the effects of rTMS on motor cortical excitability and inhibition.Clinical Neurophysiology,117(12), 2584–2596.
Gazzola, V., Aziz-Zadeh, L., & Keysers, C. (2006). Empathy and the somatotopic auditory mirror system in humans.Current Biology,16, 1824–1829.
Hadjikhani, N., Joseph, R. M., Snyder, J., & Tager-Flusberg,H. (2006). Anatomical differences in the mirror neuron system and social cognition network in autism.Cerebral Cortex, 16, 1276–1282.
Haffey, A., Press, C., O'Connell, G., & Chakrabarti, B.(2013). Autistic traits modulate mimicry of social but not nonsocial rewards.Autism Research, 6(6), 614–620.
Hamilton, A. F. de C. (2013a). Reflecting on the mirror neuron system in autism: A systematic review of current theories.Developmental Cognitive Neuroscience, 3, 91–105.
Hamilton, A. F. de C. (2013b). The mirror neuron system contributes to social responding.Cortex, 49(10), 2957–2959.
Hansen, S. N., Schendel, D. E., & Parner, E. T. (2015).Explaining the increase in the prevalence of autism spectrum disorders: The proportion attributable to changes in reporting practices.JAMA Pediatrics, 169(1), 56–62.
Hickok, G. (2014).The myth of mirror neurons: The real neuroscience of communication and cognition. New York:W. W. Norton & Company.
Hohwy, J., & Palmer, C. (2014). Social cognition as causal inference: Implications for common knowledge and autism. In M. Gallotti & J. Michael (Eds.),Perspectives on social ontology and social cognition(pp. 167–189).Netherlands: Springer.
Iacoboni, M. (2008). Mesial frontal cortex and super mirror neurons.Behavioral and Brain Sciences, 31, 30.
Iacoboni, M., & Dapretto, M. (2006). The mirror neuron system and the consequences of its dysfunction.Nature Review Neuroscience,7, 942–951.
Iacoboni, M., Molnar-Szakacs, I., Gallese, V., Buccino, G.,Mazziotta, J. C., & Rizzolatti, G. (2005). Grasping the intentions of others with one’s own mirror neuron system.PLoS Biology,3(3), e79.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H.,Mazziotta, J. C., & Rizzolatti, G. (1999). Cortical mechanisms of human imitation.Science, 286(5449),2526–2528.
Janssen, P., & Scherberger, H. (2015). Visual guidance in control of grasping.Annual Review of Neuroscience, 38,69–86.
Jarrett, C. (2014).Great myths of the brain. London: Wiley Blackwell.
Kana, R. K., Wadsworth, H. M., & Travers, B. G. (2011). A systems level analysis of the mirror neuron hypothesis and imitation impairments in autism spectrum disorders.Neuroscience and Biobehavioral Review, 35, 894–902.
Keysers, C., & Gazzola, V. (2014). Hebbian learning and predictive mirror neurons for actions, sensations and emotions.Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1644), 20130175.
Kilner, J. M., & Lemon, R. N. (2013). What we know currently about mirror neurons.Current Biology, 23(23),R1057–R1062.
Klin, A., Jones, W., Schultz, R., & Volkmar, F. (2003). The enactive mind, or from actions to cognition: Lessons from autism.Philosophical Transactions of the Royal Society B:Biological Sciences, 358, 345–360.
Kosonogov, V. (2012). Why the mirror neurons cannot support action understanding.Neurophysiology, 44(6),499–502.
Lai, M. C., Lombardo, M. V., & Baron-Cohen, S. (2014).Autism.Lancet, 383(9920), 896–910.
Lingnau, A., Gesierich, B., & Caramazza, A. (2009).Asymmetric fMRI adaptation reveals no evidence for mirror neurons in humans.Proceedings of the National Academy of Sciences of the United States of America,106(24), 9925–9930.
Marsh, L. E., & Hamilton, A. F. de C. (2011). Dissociation of mirroring and mentalising systems in autism.NeuroImage,56(3), 1511–1519.
Martineau, J., Andersson, F., Barthélémy, C., Cottier, J. P., &Destrieux, C. (2010). Atypical activation of the mirror neuron system during perception of hand motion in autism.Brain Research, 1320, 168–175.
Martineau, J., Cochin, S., Magne, R., & Barthélémy, C.(2008). Impaired cortical activation in autistic children: Is the mirror neuron system involved?International Journal of Psychophysiology, 68(1), 35–40.
Michel, T. M., Herholz, S., Finkelmeier, A., Schneider, F.,Brügmann, E., Haeck, M., … Habel, U. (2011). P03-298-the neuronal correlates of empathy in autism spectrum disorders.European Psychiatry, 26(supp1), 1467.
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2009).Is the mirror neuron system involved in imitation? A short review and meta-analysis.Neuroscience and Biobehavioral Reviews, 33(7), 975–980.
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2012).Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies.Neuroscience and Biobehavioral Reviews, 36(1), 341–349.
Montague, P. R., Dolan, R. J., Friston, K. J., & Dayan, P.(2012). Computational psychiatry.Trends Cognitive Science,16(1), 72–80.
Mukamel, R., Ekstrom, A. D., Kaplan, J., Iacoboni, M., &Fried, I. (2010) Single-neuron responses in humans during execution and observation of actions.Current Biology,20(8), 750–756.
Newman-Norlund, R. D., van Schie, H. T., van Zuijlen, A. M.J., & Bekkering, H. (2007). The mirror neuron system is more active during complementary compared with imitative action.Nature Neuroscience, 10, 817–818.
Oberman, L. M., Hubbard, E. M., McCleery, J. P., Altschuler,E. L., Ramachandran, V. S., & Pineda, J. A. (2005). EEG evidence for mirror neuron dysfunction in autism spectrum disorders.Cognitive Brain Research, 24, 190–198.
Perkins, T. J., Bittar, R. G., McGillivray, J. A., Cox, I. I., &Stokes, M. A. (2015). Increased premotor cortex activation in high functioning autism during action observation.Journal of Clinical Neuroscience, 22, 664–669.
Perkins, T. J., Stokes, M., A., McGillivray, J., & Bittar, R.(2010). Mirror neuron dysfunction in autism spectrum disorders.Journal of Clinical Neuroscience, 17, 1239–1243.
Pfeiffer, U. J., Vogeley, K., & Schilbach, L. (2013). From gaze cueing to dual eye-tracking: Novel approaches to investigate the neural correlates of gaze in social interaction.Neuroscience and Biobehavioral Reviews, 37, 2516–2528.
Pineda, J. A., Carrasco, K., Datko, M., Pillen, S., & Schalles,M. (2014). Neurofeedback training produces normalization in behavioural and electrophysiological measures of high-functioning autism.Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1644),20130183.
Pokorny, J. J., Hatt, N. V., Colombi, C., Vivanti, G., Rogers,S. J., & Rivera, S. M. (2015). The action observation system when observing hand actions in autism and typical development.Autism Research, 8(3), 284–296.
Puzzo, I., Cooper, N. R., Cantarella, S., Fitzgerald, P. B., &Russo, R. (2013). The effect of rTMS over the inferior parietal lobule on EEG sensorimotor reactivity differs according to self-reported traits of autism in typically developing individuals.Brain Research, 1541, 33–41.
Ramachandran, V. S., & Oberman, L. M. (2006). Broken mirrors: A theory of autism.Scientific America, 295, 62–69.
Raymaekers, R., Wiersema, J. R., & Roeyers, H. (2009).EEG study of the mirror neuron system in children with high functioning autism.Brain Research, 1304, 113–121.
Rizzolatti, G., & Fabbri-Destro, M. (2010). Mirror neurons:From discovery to autism.Experimental Brain Research,200(3–4), 223–237.
Rizzolatti, G., & Sinigaglia, C. (2010). The functional role of the parieto-frontal mirror circuit: Interpretations and misinterpretations.Nature Reviews Neuroscience, 11,264–274.
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system.Annual Review of Neuroscience, 27, 169–192.
Rizzolatti, G., Fogassi, L., & Gallese, V. (2006). Mirrors in the mind.Scientific American, 295(5), 54–61.
Robinson, E. B., Koenen, K. C., McCormick, M. C., Munir,K., Hallett, V., Happé, F., … Ronald, A. (2011). Evidence that autistic traits show the same etiology in the general population and at the quantitative extremes (5%, 2.5%,and 1%).Archives of General Psychiatry, 68(11), 1113–1121.
Ronald, A., & Hoekstra, R. A. (2011). Autism spectrum disorders and autistic traits: A decade of new twin studies.American Journal of Medical Genetics, 3, 255–274.
Ruysschaert, L., Warreyn, P., Wiersema, J. R., Oostra, A., &Roeyers, H. (2014). Exploring the role of neural mirroring in children with autism spectrum disorder.Autism Research,7, 197–206.
Schilbach, L. (2014). On the relationship of online and offline social cognition.Frontiers in Human Neuroscience,8, 278.
Schilbach, L. (2015). Eye to eye, face to face and brain to brain: Novel approaches to study the behavioral dynamics and neural mechanisms of social interactions.Current Opinion in Behavioral Sciences, 3, 130–135.
Schilbach, L., Eickhoff, S. B., Cieslik, E. C., Kuzmanovic,B., & Vogeley, K. (2012). Shall we do this together?Social gaze influences action control in a comparison group, but not in individuals with high-functioning autism.Autism, 16(2), 151–162.
Schilbach, L., Timmermans, B., Reddy, V., Costall, A., Bente,G., Schlicht, T., & Vogeley, K. (2013). Toward a second-person neuroscience.Behavioral and Brain Sciences, 36, 393–462.
Shaw, D. J., & Czekóová, K. (2013). Exploring the development of the mirror neuron system: Finding the right paradigm.Developmental Neuropsychology, 38(4),256–271.
Simmons, D. R., Robertson, A. E., McKay, L. S., Toal, E.,McAleer, P., & Pollick, F. E. (2009). Vision in autism spectrum disorders.Vision Research, 49, 2705–2739.
Sims, T. B., Neufeld, J., Johnstone, T., & Chakrabarti, B.(2014). Autistic traits modulate frontostriatal connectivity during processing of rewarding faces.Social Cognitive Affective Neuroscience, 9(12), 2010–2016.
Smith, I. M., & Bryson, S. E. (2007). Gesture imitation in autism: II. Symbolic gestures and pantomimed object use.Cognitive Neuropsychology, 24, 679–700.
Southgate, V., & Hamilton, A. F. de C. (2008). Unbroken mirrors: Challenging a theory of autism.Trends in Cognitive Sciences, 12(6), 225–229.
Sowden, S., Koehne, S., Catmur, C., Dziobek, I., & Bird, G.(2015). Intact automatic imitation and typical spatial compatibility in autism spectrum disorder: Challenging the broken mirror theory.Autism Research,doi:10.1002/aur.1511
Spaulding, S. (2013). Mirror neurons and social cognition.Mind and Language, 28(2), 233–257.
Spunt, R. P., Falk, E. B., & Lieberman, M. D. (2010).Dissociable neural systems support retrieval of how and why action knowledge.Psychological Science, 21, 1593–1598.
Stanovich, K. E. (2012).How to think straight about psychology(10th ed.). Upper Saddle River NJ: Pearson Education.
Steinhorst, A., & Funke, J. (2014). Mirror neuron activity is no proof for action understanding.Frontiers in Human Neuroscience, 8, 333.
Toal, F., Daly, E. M., Page, L., Deeley, Q., Hallahan, B.,Bloemen, O., … Murphy, D. G. M. (2010). Clinical and anatomical heterogeneity in autistic spectrum disorder: A structural MRI study.Psychological Medicine, 40(7),1171–1181.
Virji-Babul, N., Rose, A., Moiseeva, N., & Makan, N. (2012).Neural correlates of action understanding in infants:Influence of motor experience.Brain and Behavior, 2(3),237–242.
Vivanti, G., Nadig, A., Ozonoff, S., & Rogers, S. J. (2008).What do children with autism attend to during imitation tasks?Journal of Experimental Child Psychology, 101(3),186–205.
Wang, X. J., & Krystal, J. H. (2014). Computational psychiatry.Neuron, 84, 638–654.
Wang, Y., & Quadflieg, S. (2015). In our own image?Emotional and neural processing differences when observing human-human vs human-robot interactions.Social Cognitive and Affective Neuroscience, 10, 1515–1524.
Wicker, B., Keysers, C., Plailly, J., Royet, J. P., Gallese, V.,& Rizzolatti, G. (2003). Both of us disgusted in my insula:The common neural basis of seeing and feeling disgust.
Neuron, 40(3), 655–664.
Williams, J. H. G., Whiten, A., Suddendorf, T., & Perrett, D.I. (2001). Imitation, mirror neurons and autism.Neuroscience and Biobehavioral Reviews, 25, 287–295.
猜你喜欢
文萃报·周五版(2021年14期)2021-06-08
中国生殖健康(2020年7期)2021-01-18
当代党员(2020年20期)2020-11-06
小康(2018年23期)2018-08-23
现代装饰(2018年5期)2018-05-26
海峡姐妹(2017年5期)2017-06-05
丹青少年(2017年2期)2017-02-26
中国生化药物杂志(2015年4期)2015-07-07
小康(2015年4期)2015-03-31
弹箭与制导学报(2015年1期)2015-03-11
