
不同的丁酸产品在猪禽生产中的应用研究进展
2016-01-19 01:36南京农业大学动物科技学院南京210095
动物营养学报 2015年2期
芦 娜 王 恬 钟 翔(南京农业大学动物科技学院,南京210095)
不同的丁酸产品在猪禽生产中的应用研究进展
芦 娜 王 恬 钟 翔∗
(南京农业大学动物科技学院,南京210095)
摘 要:近年来丁酸盐作为饲用抗生素的替代品,广泛应用于动物生产中。本文就丁酸的生物学功能、几种不同的丁酸产品的特点及其在猪禽生产中的应用作一综述,为实际生产中丁酸产品的应用提供依据。
关键词:丁酸产品;猪;禽;应用
近年来,抗生素的滥用现象十分严重。滥用抗生素不仅会使细菌产生抗药性,畜禽机体免疫力下降,引起畜禽内源性感染和二重感染,还会在畜产品和环境中造成残留[1]。面对日益严峻的食品安全问题,欧盟已于2006年1月起全面禁止抗生素作为生长促进剂在饲料中的使用[2]。
如今,人们不得不研制绿色环保、无残留的新型饲料添加剂,以替代抗生素的使用,而丁酸产品因为满足了这些条件而颇受欢迎。丁酸是一种主要由猪禽结肠微生物发酵而产生的挥发性脂肪酸。大量研究表明,丁酸盐可以提高猪禽生长性能,促进营养物质吸收,刺激消化酶分泌,改善肠道菌群,促进肠道上皮细胞完整性和提高机体免疫。丁酸可直接或间接进行消化道组织的更新和修复。丁酸对人类和动物的直接作用表现在可以使坏死组织(如小肠黏膜)得到迅速更新,而间接作用则与内分泌-神经-免疫系统有关,例如改善人类的肥胖、降低胆固醇合成、预防心脑血管疾病、提高记忆力等[3-5]。此外,丁酸盐可以通过直接影响病毒基因表达和宿主细胞增殖而使细菌毒性减弱。在动物生产中,丁酸盐是一种非常有效的饲料添加剂,尤其在出生不久后添加,可以提高生产性能,保障肠道健康。本文就丁酸产品的生物学功能以及几种不同的丁酸产品在猪禽生产上的应用比较作一综述。
1 机体内丁酸的来源及代谢
短链脂肪酸(short chain fatty acids,SCFAs)主要包括乙酸、丙酸和丁酸,是肠道内厌氧杆菌发酵不吸收的碳水化合物和蛋白质的产物。厌氧杆菌发酵产生SCFAs的浓度与细菌的种类、数量、肠腔中pH、肠腔中底物浓度等有密切关系。参与SCFAs生成的主要细菌有拟杆菌、双歧杆菌、梭菌及许多厌氧的革兰氏阳性菌;盲肠和结肠是生成SCFAs的主要肠段。医学研究发现,口服的SCFAs在近端的肠道全部被吸收,不能到达大肠,但通过增加食物中纤维含量,能有效增加大肠中SCFAs浓度[6]。SCFAs主要来源于食物中的纤维成分,未吸收的淀粉、蛋白质及内源性多糖也能提供少量SCFAs[7]。
SCFAs通过结肠细胞顶端膜被机体吸收,未解离的SCFAs以脂溶形式扩散吸收[8]或通过SCFAs/HCO-3离子交换吸收[9];游离的SCFAs由SCFAs转运载体进行主动运输,SCFAs转运载体包括钠耦合单羧酸转运体(SMCT1)[10]和耦合于跨膜H+梯度的单羧酸转运亚型1(MCT1)[11]。被机体吸收的丁酸,一部分被盲肠、结肠上皮细胞作为能源物质,另一部分被肝脏细胞代谢用于糖异生[12]。
2 丁酸的生物学作用
目前,关于丁酸及其产品的生物学作用的研究大多以人、大鼠、小鼠为试验对象,猪、禽方面的报道较少。
2.1 优化消化道内菌群结构
健康动物的消化道存在着一个稳定的微生物区系,乳酸菌、双歧杆菌等有益菌占绝对优势,然而饲粮及环境等因素容易打破该平衡,引起消化道菌群失调,导致疾病发生。动物摄入丁酸盐后,其有效成分丁酸可以避开胃和小肠而直接进入盲肠和结肠中发挥作用。丁酸进入细菌细胞后,分解为丁酸根离子和氢离子,高浓度的氢离子能够使大肠杆菌、沙门氏菌等有害菌大量死亡,而乳酸菌等有益菌则由于其耐酸性而大量增殖[3]。Galfi等[13]研究发现,丁酸钠可以显著降低猪肠道内沙门氏菌的繁殖及在粪中的排放,降低回肠大肠杆菌的数量。郭传珍等[14]试验表明,饲粮中添加不同剂量的丁酸钠,对肉鸡消化道各段pH均有一定的降低作用,且能够提高乳酸杆菌的数量,抑制大肠杆菌的繁殖。钮海华等[15]研究发现,在饲粮中添加包膜丁酸钠可促进肠道乳酸杆菌的增殖,降低大肠埃希菌的数量。Abdel⁃Mageed[16]研究表明,饲粮中添加0.2%丁酸,可以减少日本鹌鹑肠道微生物区系数量,尤其是病原菌数量,可以提高生长性能和免疫力。
2.2 维持肠黏膜的正常屏障作用
肠道不仅是消化吸收的重要场所,而且具有重要的屏障功能。肠道黏膜屏障由机械屏障、化学屏障、免疫屏障及生物屏障组成。
机械屏障由黏液层和肠上皮细胞组成。肠道上皮表面覆盖着一层黏液,其主要成分是黏液蛋白,这层黏液形成一道保护性屏障,保护肠道上皮细胞免受机械性、化学性、酶类及微生物的破坏。肠上皮细胞作为一个选择性屏障,一方面允许营养物质和其他可溶性物质被吸收;另一方面能够阻止肠腔中的微生物、毒素等进入体内[17]。丁酸盐主要在空肠和回肠段被吸收,并且作为细胞的能量来源可以促进消化道细胞的增殖和成熟,从而维持肠黏膜上皮细胞结构完整性,增强机械屏障作用。赵会利等[18]试验表明,饲粮中添加1%丁酸钠可促进犊牛断奶前后瘤胃绒毛和小肠绒毛的生长,维持肠道黏膜完整性,促进犊牛的胃肠道发育。钟翔等[19]认为,饲粮中添加丁酸钠能够改善断奶仔猪肠道黏膜形态结构,促进消化器官的生长发育。
肠道免疫屏障主要由肠道相关淋巴组织组成。肠道相关淋巴组织包括派伊尔结(Peyer’pat⁃ches,PP)、弥散性黏膜淋巴样细胞及肠系膜淋巴结。王继凤[20]研究发现,丁酸钠可以引起断奶仔猪小肠上皮内淋巴细胞、肥大细胞和肠黏膜杯状细胞的数量不同程度地增加。
肠道中栖息的大量微生物菌群构成了生物屏障。丁酸盐不仅可以进入细菌细胞内分解为丁酸根离子和氢离子,随着胞内氢离子浓度的提升,抑制对氢离子耐受性差的大肠杆菌、沙门氏菌等病原微生物的生长,从而减少病原菌与动物机体争夺营养物质,减少其他消化道疾病的发生;而且丁酸盐还可以促进部分有益菌的生长,使有益菌在肠道内的微生物群内处于优势地位。
2.3 抑制肠道炎症反应
炎症性肠病(inflammatory bowel disease,IBD)包括溃疡性结肠炎(ulcerative colitis,UC)和克罗恩病(Crohn’s disease,CD),是消化道常见疾病之一。
肠道黏液层主要由黏蛋白2(MUC2)、三叶因子3(TFF3)、分泌型免疫球蛋白A(sIgA)等物质组成,是保护肠道上皮细胞的重要防线。Song等[21]研究发现,在三硝基苯磺酸(trinitro⁃benzene⁃sulfonic acid,TNBS)诱导的结肠炎模型小鼠体内,给予丁酸钠可以上调TFF3 mRNA的表达。Paas⁃sen等[22]对人类大肠癌细胞LS174T给予1 mmol/L丁酸盐培养,结果发现,丁酸盐能够增强MUC2 mRNA的表达。但也有研究[23]发现,对溃疡性结肠炎患者采用丁酸盐灌肠治疗,并不影响结肠黏膜中MUC2、TFF3以及sIgA mRNA的表达。
肠黏膜促炎因子水平较高是克罗恩病的一项特征[24]。Sabatino等[25]为了研究口服丁酸钠对克罗恩病的治疗效果,选择13名患者,给予4 g/d的包被片剂丁酸钠,8周后发现,核因子κB(NF⁃κB)和白细胞介素(IL)⁃1β的水平显著降低。Segain等[26]研究发现,丁酸可通过抑制蛋白激酶(IκBα)降解,使NF⁃κB不能从NF⁃κB和IκBα复合物中分离出来,NF⁃κB不能进入细胞核,进而使肿瘤坏死因子α(TNF⁃α)、IL⁃1β、IL⁃6等促炎因子的mRNA不能转录,使得促炎因子不能表达,炎症反应被抑制。赖衍宗等[27]以TNBS诱导大鼠结肠炎模型并予丁酸钠治疗或美沙拉嗪治疗,同时设置正常对照组和结肠炎模型组。结果发现,与结肠炎模型组相比,丁酸钠治疗组一般情况和结肠组织学表现有所改善,美沙拉嗪治疗组则有明显改善。丁酸钠治疗组和美沙拉嗪治疗组结肠组织转化生长因子β1(TGF⁃β1)阳性表达率显著高于结肠炎模型组,慢性炎症期血浆IL⁃8水平显著降低,IL⁃10水平显著增高,表明丁酸钠对TNBS结肠炎模型大鼠的肠黏膜修复有一定促进作用,该作用可能与其增加TGF⁃β1表达和调节促炎细胞因子与抗炎细胞因子平衡有关。Singh等[28]研究发现,GPR109A(由Niacr1编码)是丁酸在结肠的受体,能够促进结肠巨噬细胞和树突细胞的抗炎症作用,并且能够使它们诱导调节性T细胞和产生IL⁃10的T细胞的分化。另外,GPR109A在丁酸诱导结肠上皮细胞产生IL⁃18过程中起着重要作用。上述研究证明丁酸及其盐类可抑制促炎因子的分泌,从而缓解肠道炎症。
2.4 抑制肿瘤生长,调控细胞凋亡
组蛋白是动物染色体的基本组成单位,组蛋白的乙酰化在基因表达中起着决定性的作用。癌细胞中的组蛋白去乙酰化酶(histone deaceylase,HDAC)过度表达造成的乙酰化失衡导致肿瘤发生,而通过抑制HDAC可以有效抑制肿瘤的发生。HDAC抑制剂能够抑制多种肿瘤生长,根据其结构特征,可分为异羟肟酸类、苯甲酰胺类、环肽类和短链脂肪酸类。诸多研究表明,丁酸盐作为组蛋白去乙酰化酶抑制剂,对多种肿瘤细胞表现出良好的抗肿瘤作用,其可以使肿瘤细胞出现明显的细胞凋亡、增生抑制以及细胞周期阻滞等。
细胞凋亡是在细胞内外因素刺激之下细胞主动的自杀过程,表现为细胞体积缩小,核浓缩、染色质边集,染色体DNA断裂,胞质浓缩及凋亡小体形成等。细胞凋亡过程中有基因的转录和蛋白质合成,其中B淋巴细胞瘤-2(Bcl⁃2)家族凋亡基因表达的改变尤为重要。细胞淋巴瘤-2相关基因(Bax)是促进细胞凋亡的基因。胡卫[29]为研究丁酸钠对结肠上皮细胞凋亡的影响及作用机制,将离体大鼠结肠黏膜固定于Ussing槽中孵育,黏膜面给予4%乙酸,并与正常结肠黏膜对照;在体试验,用醋酸诱导大鼠结肠炎。3 d后随机分为2组,每天分别给予丁酸钠、生理盐水灌肠1次。第11天处死后,取结肠组织进行脱氧核糖核苷酸末端转移酶介导的缺口末端标记法(TUNEL)检测及免疫组化分析结肠上皮细胞凋亡情况。结果发现,丁酸钠灌肠治疗组平均凋亡指数、凋亡指数和Bax表达水平明显低于对照组,可以推断出似于体内肠腔正常浓度的丁酸钠(10 mmol/L)在体外体内对正常和炎症结肠上皮细胞有抑制其凋亡的作用。王凌[30]发现,丁酸钠通过调控线粒体膜上凋亡相关基因Bcl⁃2和Bax表达,使Bcl⁃2/Bax下降,降低线粒体跨膜电位,从而激活下游的含半胱氨酸的天冬氨酸蛋白水解酶(caspase)⁃9及其后的caspase⁃3等级联反应,完成细胞凋亡过程。
丁酸钠能够诱导胞内p21蛋白量增加。p21是一种重要的细胞周期调控蛋白,参与细胞的生长、分化、衰老及死亡过程,同时又与肿瘤发生密切相关。可抑制细胞周期蛋白依赖性蛋白激酶(CDK)的活性,并可直接结合增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)而抑制细胞DNA合成,阻滞细胞周期分裂。陈萍等[31]发现,丁酸钠可通过降低携带B细胞特异单克隆鼠白血病毒整合位点1基因(Bmi⁃1)表达从而激活p21蛋白表达,诱导人食管鳞癌细胞KYSE⁃150细胞周期阻滞、DNA损伤和凋亡。
2.5 缓解机体氧化应激
丁酸盐不仅影响细胞及机体炎症过程,还影响癌变的发生与发展,而这2个过程都涉及到氧化应激[32-33]。氧化应激是机体内活性氧族、活性氮族以及抗氧化物质防御机制的失衡,会引起一系列损害细胞的脂质、蛋白质和DNA的级联反应[34]。
近年来,一些研究表明,丁酸及其盐类能够调控氧化应激。Abrahamse等[35]发现丁酸能够显著降低H2O2诱导的DNA损伤。大鼠摄入抗性淀粉能够降低机体由于摄入高蛋白质饲粮而引起的结肠细胞DNA损伤,DNA损伤与盲肠内丁酸浓度呈负相关[36]。张卫辉[37]试验表明,饲粮中添加丁酸钠不但可以提高正常生理状况下肉鸡的抗氧化功能,还可以抑制由于饲喂皮质醇(CORT)引起的肠黏膜抗氧化水平的下降。Hamer等[38]在人类上的研究发现,100 mmol/L丁酸钠能够显著提高结肠还原型谷胱甘肽(GSH)的含量并且极显著降低尿酸浓度。此外,研究发现,丁酸钠能够逆转和抑制D-苯丙胺(D⁃AMPH)诱导的大脑结构的氧化损伤,并为研究治疗情绪障碍的药物提供可能性[39]。
3 不同的丁酸产品的比较
3.1 丁酸钠
丁酸钠是正丁酸的钠盐,分子式为C4H7O2Na,相对分子质量为110.086 9,熔点250~253℃,呈白色或类白色绒毛状粉末,具有特殊的奶酪酸败样气味,易溶于水,水溶液pH呈碱性。在常规储存和搬运条件下,室温和密闭状态下稳定,但不可与性质相反的物质、粉尘、强氧化剂混存。丁酸钠可能分解产生有害物质,如一氧化碳、二氧化碳、氧化钠、过敏性和有毒的烟尘与气体,可能导致中枢神经系统抑制、眼睛与皮肤过敏、呼吸道与消化道应激。
由于丁酸具有游离性和挥发性的特点,饲料生产中常将其制成相对稳定的钠盐,即丁酸钠。丁酸钠具有水脂两亲性,但偏重于亲脂,在pH 4~6时不容易被分解,主要以非粒子形式被肠黏膜上皮弥散性吸收,也存在与碳酸氢根离子交换吸收的方式,在结肠段吸收最高。丁酸钠在2006年被农业部列入《饲料添加剂品种目录》,其作为酸化剂、电解质平衡调节剂、胃肠道微生态平衡调节剂、诱食剂等的组成成分发挥着特殊作用。
虽然目前丁酸钠在动物生产中被广泛应用,但是仍存在一些问题:1)添加量较大,一般为0.1%~0.3%;2)未经处理流动性差,不易混匀;3)容易与饲料中的维生素、碱性物质等发生反应,影响使用效果;4)进入机体后在胃中解离速度很快,很难到达小肠和大肠,不能在整个消化道中有效发挥作用;5)具有刺激性臭味,影响动物采食量。
3.2 包膜丁酸钠
没有经过保护处理的丁酸钠刺激性较强,容易吸潮,有奶酪酸败的气味,在饲料加工过程和畜禽的胃中容易损失和分解,只有添加量较大时才会到达肠道,而丁酸钠的主要功能是在肠道中发挥。而包膜丁酸钠解决了上述问题,其采用进口特殊植物脂肪包被材料,经特殊包膜工艺处理后,消除了丁酸钠的恶臭气味;有效减少丁酸钠对消化道的刺激,并且可以阻断丁酸钠与饲料中其他组分之间的化学反应,质量稳定,使用方便;选用的特殊包膜材料可以在肠道中逐步释放,因此可做到丁酸钠在整个肠道持续释放,尤其是结肠段的靶向释放,防治结肠炎引起的营养性腹泻发生,改善粪便形态,最大限度地保护动物的肠道健康。
3.3 硅胶控释型丁酸
虽然包膜丁酸钠较普通丁酸钠有更多优点,但包膜技术工艺复杂,成本高昂。因此价格更为低廉、作用效果更好的替代品被开发出来,硅胶控释型丁酸(silica gel controlled⁃release butyrate,SG⁃B)就是其中一种。
硅胶是一种高活性吸附材料,物理及化学性质稳定,具有吸附性高、热稳定性好的特点。目前,硅胶主要被用作吸附剂、干燥剂、增稠剂、填充剂和催化剂载体等。陈禹银等[40]通过研究硅胶吸附丁酸的计量置换吸附模型,发现丁酸在硅胶上的吸附是自发、放热的熵减少过程,随着温度的升高,吸附量减少。胡彩虹等[41]采用室温过量浸渍法制备和表征SG⁃B,热重分析表明丁酸负载量为41.1%,钱仲仓[42]在实验室制备和表征SG⁃B,发现丁酸负载量为37.5%,这与陈禹银等[40]的研究结果基本一致。
通过硅胶对丁酸的控释作用解决了普通丁酸钠不能到达动物后肠的问题,这是SG⁃B显著改善空肠和回肠黏膜形态、保护肠道功能的主要原因。
3.4 丁酸甘油酯和三丁酸甘油酯
丁酸甘油酯是一种短链甘油酸酯,由1分子甘油与3分子丁酸酯化而成,分子式为C16H26O6,存在于奶油、鱼肝油与汗液中。丁酸甘油酯是丁酸和甘油酯化后的产物,是多元醇型非离子型表面活性剂。从其分子结构上看,既能作油包水(W/O)型乳化剂,又可以作水包油(O/W)型乳化剂,将丁酸甘油酯作为一种新型的乳化剂制备微胶囊粉,添加到饲料中,使用方便,安全无毒副作用,还有着特殊的香味,既解决了丁酸为液态难以添加的特点,又改善了直接使用丁酸气味难闻的缺点。丁酸甘油酯被动物采食后,以丁酸和甘油2种营养物的形式被机体吸收。其中丁酸具有供应细胞能量,影响内分泌,促进上皮组织生长发育,抗菌消炎等功能,被认为是抗生素的潜在替代物,从根本上消除抗生素在动物产品中的残留;同时减少饲料中其他乳化剂的用量,提高脂肪和饲料的消化率,降低饲料生产成本和畜禽行业的饲养成本,为生产优质无公害动物性产品提供转思路,具有良好的经济和社会效益[43]。
三丁酸甘油酯(tributyrin)天然存在于乳脂中,由3分子丁酸和1分子甘油组成,化学式为C15H26O6,为白色近油状液体,几乎没有气味或略有脂肪香气。易溶于乙醇、乙醚等有机溶剂,极难溶于水。三丁酸甘油酯作为丁酸的前体物,使用方便,安全无毒副作用,无臭味,能够促进畜禽肠道健康发育,提高机体免疫力,促进营养物质消化吸收,进而提高动物生产性能,是目前较好的营养性添加剂产品[44]。
4 不同丁酸产品在猪禽生产中的应用
4.1 在猪生产中的应用
4.1.1 仔猪
仔猪早期断奶是现代化养猪生产中提高母猪生产性能的重要方法之一,但早期断奶仔猪消化系统和免疫系统发育不完全,加之饲粮、环境等应激,使早期断奶仔猪出现腹泻、免疫功能降低等一系列问题。
断奶对仔猪消化酶的活性和酶谱造成深刻影响。仔猪断奶后采食乳糖的比例减少,乳糖酶的分泌量与活性下降[45]。仔猪消化道内源酶的分泌量较低,胃内仅有凝乳酶、胃蛋白酶,且含量仅为成年猪的1/4~1/3;胃底腺不发达,不能分泌足够盐酸;缺乏游离的盐酸,胃蛋白酶没有活性,不能消化蛋白质,特别是植物性蛋白质[46]。早期断奶仔猪的小肠出现严重的绒毛萎缩、隐窝变深、肠黏膜淋巴细胞增生和隐窝细胞有丝分裂速度加快。仔猪提早断奶,不仅抑制其合成抗体的能力,还会降低细胞免疫水平,断奶时间越早,影响越大。
因此,为缓解早期断奶对仔猪的应激,需要在饲料中添加一些可促进仔猪肠道发育、增强免疫力、提高生产性能、防治下痢等功能性饲料添加剂。丁酸盐因其满足上述条件而被广泛应用于仔猪生产,其应用效果见表1。
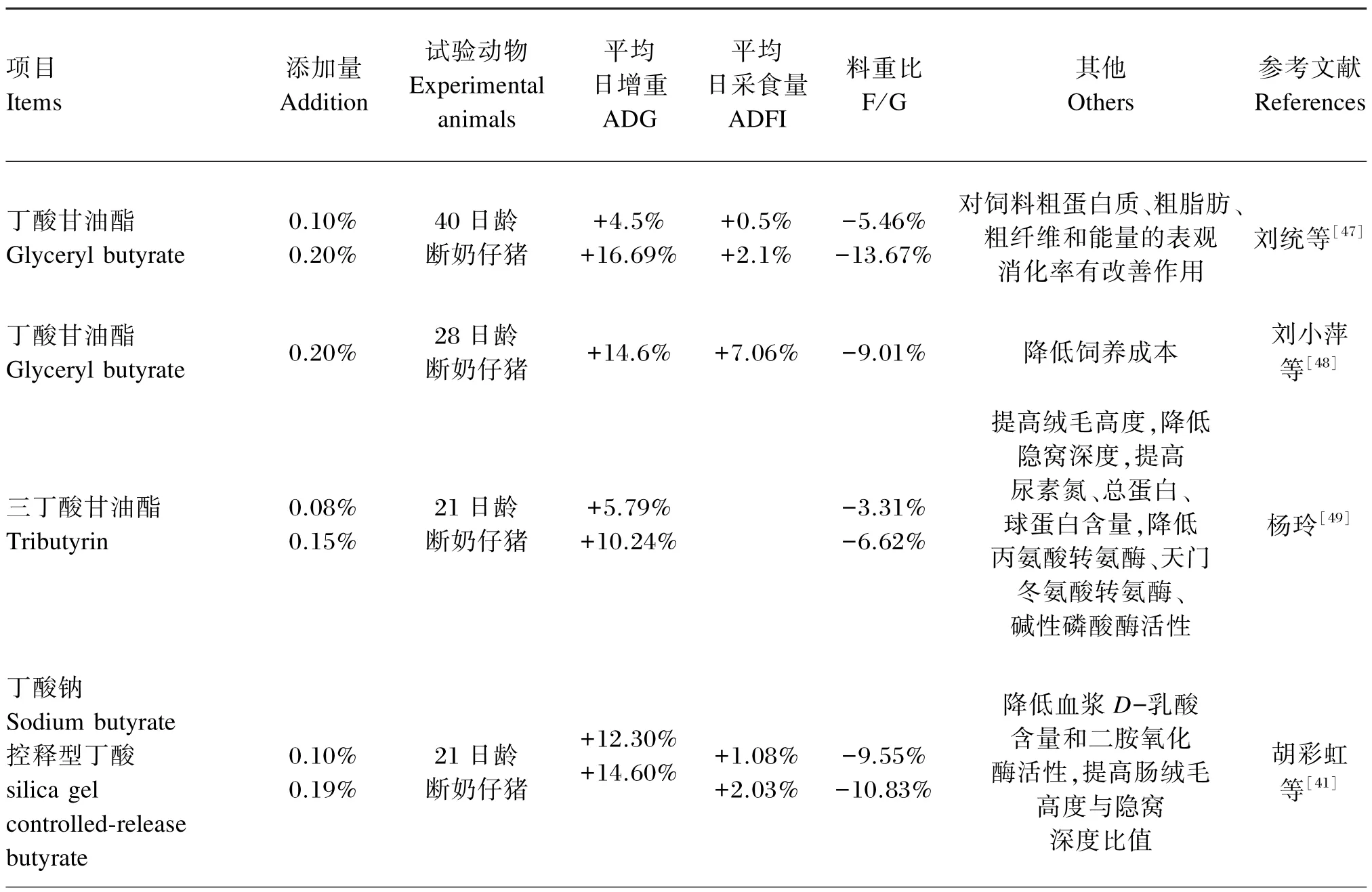
表1 不同丁酸产品在仔猪生产中的应用效果Table 1 Application effects of different types of butyrate products on piglets production

续表1
以上试验结果表明,丁酸产品不仅可以提高仔猪的生产性能,还能够增强其免疫力。根据目前的研究可推断,丁酸产品在仔猪上的添加量可控制在0.05%~0.20%。
4.1.2 母猪
丁酸能够提高子宫平滑肌收缩力,可以使母猪顺产,减少难产的发生,同时,防止母猪骨骼中钙的过度流失,减少母猪腿病发生,种用期得以延长。侯嘉等[57]在泌乳母猪饲粮中添加1 kg/t微囊丁酸钠,减少了母猪分娩助产比例,缩短了产程,提高断奶母猪7 d内的发情率。其降低助产比例与缩短产程的原因可能与其促进钙的吸收有关,钙的充分吸收,提高了子宫平滑肌收缩力,可以促使母猪顺产,减少难产发生率,降低助产引起的子宫炎和阴道炎等生殖疾病的发生率。
此外,丁酸产品还能够提高母猪的生产性能。张小东等[58]将40头妊娠85 d健康二元杂交经产母猪随机分为对照组(基础饲粮)、试验组(妊娠组:基础饲粮+800 g/t微囊丁酸钠;哺乳组:基础饲粮+500 g/t微囊丁酸钠),结果表明,饲粮中添加微囊丁酸钠能够显著提高母猪生产后期采食量(4.56%),显著降低母猪哺乳期的体失重,提高仔猪断奶日增重和断奶体重。毛石明[59]选择胎次相同、体况相近且同期配种的15头健康妊娠后期二元母猪,随机分为3组,分别饲喂基础饲粮(对照组)、基础饲粮+400 g/t包膜丁酸钠和基础饲粮+600 g/t包膜丁酸钠,直到仔猪断奶(28日龄),结果发现,与对照组相比,400和600 g/t包膜丁酸钠分别提高仔猪初生窝质量1.44%和1.90%,提高初生个体质量2.0%和2.6%;经过28 d的哺乳期后,2个剂量包膜丁酸钠分别提高仔猪断奶个体重和平均日增重达4.71%、6.99%和5.41%、8.63%。
4.2 在禽生产中的应用
周磊等[60]为研究微囊丁酸钠对罗曼蛋种鸡生产性能的影响,选取8 000只29周龄健康的罗曼蛋种鸡,随机分为2个组,分别为对照组和试验组。试验组基础饲粮中添加500 g/t微囊丁酸钠。结果发现,微囊丁酸钠使试验组的产蛋率和正品率分别高1.87%和1.21%,表明饲粮中添加微囊丁酸钠能够延长蛋种鸡产蛋高峰期,提高蛋种鸡的产蛋率和正品蛋率,从而提高了经济效益。
赵民等[61]为研究包膜丁酸钠对青脚麻种鸡生产性能的影响,在试验组添加500 mg/kg包膜丁酸钠,结果表明,与对照组相比,包膜丁酸钠组蛋鸡产蛋率提高2.76%,差异显著,高峰产蛋率显著提高6.28%,种蛋合格率显著提高3.16%,死淘率显著降低49.28%。
彭丽莎等[62]研究了三丁酸甘油酯对肉鸡生长性能、养分表观消化率、屠宰性能、肠道形态及微生物菌群的影响,试验组分别饲喂在基础饲粮中添加0.10%、0.15%、0.20%、0.30%、0.40%三丁酸甘油酯的试验饲粮,结果表明:饲粮中添加三丁酸甘油酯有促进肉鸡生长的趋势,其中0.20%三丁酸甘油酯组效果最佳;与对照组相比,0.20%三丁酸甘油酯组的粗脂肪和粗纤维表观消化率均显著提高;肉鸡屠宰性能有改善的趋势,但差异不显著;十二指肠和空肠肠绒毛高度均显著提高,隐窝深度有变浅的趋势,但差异不显著;盲肠乳酸菌数量显著增加,大肠杆菌数量显著减少。在本试验中,肉鸡饲粮中三丁酸甘油酯适宜添加量为0.20%。
邹杨等[63]为了研究饲粮中添加粉剂和包膜丁酸钠对肉仔鸡生产性能、肠道pH以及挥发性脂肪酸(VFA)含量的影响,设置对照组、抗生素组(杆菌肽锌40 mg/kg+硫酸黏杆菌素8 mg/kg)、粉剂丁酸钠组(200 mg/kg粉剂丁酸钠)和包膜丁酸钠组(200 mg/kg包膜丁酸钠),研究结果表明:与对照组相比,丁酸钠对肉仔鸡日均采食量影响显著,添加包膜丁酸钠显著降低了肉仔鸡21~42日龄的料重比。丁酸钠的添加有降低肠道pH的趋势。饲粮中添加200 mg/kg粉剂丁酸钠,可以提高肠道的6.72%的乙酸含量,提高23.08%的丙酸含量,提高49.01%的丁酸含量;添加200 mg/kg包膜丁酸钠,可以提高肠道的10.39%的乙酸含量,提高33.61%的丙酸含量,提高36.80%的丁酸含量。
以上研究表明,不同的丁酸产品均有提高家禽生产性能的作用(表2)。目前国内有关丁酸盐在家禽上的研究较少,不同的丁酸产品的添加量、应用效果等有待进一步研究。
5 小 结
随着抗生素问题的日益突显,越来越多的替代品相继问世。丁酸产品凭借其无污染、无残留等优点而得到广泛关注。丁酸在体内迅速吸收,并且参与机体代谢,能够改善肠道结构,调节肠道微生物菌群,提高免疫力,从而促进动物的生长。综上所述,不同的丁酸产品都可在一定程度上提高动物生产性能,但是,丁酸产品的作用效果受到添加剂量、添加的时间跨度、动物的品种和日龄、观测的位点以及其他不可控因素的影响。因此,丁酸盐对动物的最适宜添加量、对肠道的精确作用方式及全面的分子作用机理有待进一步研究。

表2 不同丁酸产品在家禽生产中的应用效果Table 2 Application effects of different types of butyrate products on poultry production
参考文献:
[1] PAPATSIROS V G,CHRISTODOULOPOULOS G,FILIPPOPOULOS L C.The use of organic acids in monogastric animals(swine and rabbits)[J].Journal of Cell and Animal Biology,2012,6:154-159.
[2] 黄春玲,黄瑞华.抗生素对动物生长与动物产品质量的影响[J].饲料广角,2011(3):36-38.
[3] GUILLOTEAU P,MARTIN L,EECKHAUT V,et al.From the gut to the peripheral tissues:the multiple effects of butyrate[J].Nutrition Research Reviews,2010,23(2):366-384.
[4] CANANI R B,COSTANZO M D,LEONE L.The epi⁃genetic effects of butyrate:potential therapeutic impli⁃cations for clinical practice[J].Clinical Epigenetics,2012,4(1):4.
[5] 韩晓风,王鹏远,马元元,等.丁酸钠对腹膜炎小鼠肠道屏障功能的保护作用[J].中华实验外科杂志,2012,29(9):1765-1767.
[6] 詹彦.丁酸与肿瘤[J].国外医学:肿瘤学分册,2003,30(5):350-353.
[7] 康萍,印遇龙.丁酸的营养研究进展[J].中国生态农业学报,2008,16(1):252-257.
[8] VELÁZQUEZ O C,LEDERER H M,ROMBEAU J L.Butyrate and the colonocyte[M].Dietary fiber in health and disease.New York:Springer,1997:123-134.
[9] MCNEIL N I,CUMMINGS J H,JAMES W P.Rectal absorption of short chain fatty acids in the absence of chloride[J].Gut,1979,20(5):400-403.
[10] GUPTA N,MARTIN P M,PRASAD P D,et al.SLC5A8(SMCT1)⁃mediated transport of butyrate forms the basis for the tumor suppressive function of the transporter[J].Life Sciences,2006,78(21):2419-2425.
[11] CUFF M,DYER J,JONES M,et al.The human co⁃lonic monocarboxylate transporter isoform 1:its poten⁃tial importance to colonic tissue homeostasis[J].Gas⁃troenterology,2005,128(3):676-686.
[12] HIJOVA E,CHMELAROVA A.Short chain fatty acids and colonic health[J].Bratislavské LekŢrske Li⁃sty,2007,108(8):354-358.
[13] GÁLFI P,BOKORI J.Feeding trial in pigs with a diet containing sodium N⁃butyrate[J].Acta Veterinaria Hungarica,1990,38(1/2):3-17.
[14] 郭传珍,曹兵海.丁酸钠对肉鸡肠道pH值、微生物菌群和挥发性脂肪酸的影响研究[J].中国家禽,2009,31(21):14-17,21.
[15] 钮海华,王勇,马文强,等.包膜丁酸钠对仔猪肠道微生物细菌数量的影响[J].饲料研究,2010(11):34-36.
[16] ABDEL⁃MAGEED M A A.Effect of using organic acids on performance of Japanese quail fed optimal and sub⁃optimal energy and protein levels[J].Egyp⁃tian Poultry Science Journal,32(3):625-644.
[17] 曹力.乳酸杆菌对肉鸡肠道黏膜屏障功能的调节及机理[D].博士学位论文.杨凌:西北农林科技大学,2012.
[18] 赵会利,高艳霞,李建国,等.丁酸钠对断奶犊牛生长、血液生化指标及胃肠道发育的影响[J].畜牧兽医学报,2013,44(10):1600-1608.
[19] 钟翔,陈莎莎,李伟,等.丁酸钠对断奶仔猪生产性能和肠道形态的影响[J].饲料研究,2009(10):19-22.
[20] 王继凤.丁酸钠对断奶仔猪肠黏膜结构及黏膜免疫相关细胞影响的研究[D].硕士学位论文.北京:中国农业大学,2005.
[21] SONG M,XIA B,LI J.Effects of topical treatment of sodium butyrate and 5⁃aminosalicylic acid on expres⁃sion of trefoil factor 3,interleukin 1β,and nuclear fac⁃tor κB in trinitrobenzene sulphonic acid induced colitis in rats[J].Postgraduate Medical Journal,2006,82 (964):130-135.
[22] BURGER⁃VAN PAASSEN N,VINCENT A,PUIMAN P J,et al.The regulation of intestinal mucin MUC2 ex⁃pression by short⁃chain fatty acids:implications for epi⁃thelial protection[J].Biochemical Journal,2009,420 (2):211-219.
[23] HAMER H M,JONKERS D M A E,RENES I B,et al.Butyrate enemas do not affect human colonic MUC2 and TFF3 expression[J].European Journal of Gastroenterology&Hepatology,2010,22(9):1134-1140.
[24] BRYNSKOV J,NIELSEN O H,AHNFELT⁃RØNNE I,et al.Cytokines(immunoinflammatory hormones)and their natural regulation in inflammatory bowel dis⁃ease(Crohn’s disease and ulcerative colitis):a re⁃view[J].Digestive Diseases,1994,12(5):290-304.
[25] SABATINO A D,MORERA R,CICCOCIOPPO R,et al.Oral butyrate for mildly to moderately active Crohn’s disease[J].Alimentary Pharmacology&Therapeutics,2005,22(9):789-794.
[26] SEGAIN J P,DE LA BLÉTIÈRE D R,BOURREILLE A,et al.Butyrate inhibits inflammatory responses through NFκB inhibition:implications for Crohn’s disease[J].Gut,2000,47(3):397-403.
[27] 赖衍宗,章颖,杨宏升,等.丁酸钠对TNBS结肠炎模型大鼠肠黏膜修复的影响[J].胃肠病学,2011,16 (7):395-399.
[28] SINGH N,GURAV A,SIVAPRAKASAM S,et al.Activation of Gpr109a,receptor for niacin and the commensal metabolite butyrate,suppresses colonic in⁃flammation and carcinogenesis[J].Immunity,2014,40(1):128-139.
[29] 胡卫.丁酸钠对结肠上皮细胞凋亡的影响[J].武汉大学学报:医学版,2007,28(4):468-470,478.
[30] 王凌.caspase途径在丁酸钠诱导结肠癌细胞凋亡中的作用及调控[D].博士学位论文.武汉:武汉大学,2009.
[31] 陈萍,陈龙,董庆华.组蛋白去乙酰化酶抑制剂诱导人食管鳞癌KYSE-150细胞凋亡的实验研究[J].肿瘤学杂志,2013,19(9):699-703.
[32] REZAIE A,PARKER R D,ABDOLLAHI M.Oxida⁃tive stress and pathogenesis of inflammatory bowel disease:an epiphenomenon or the cause?[J].Diges⁃tive Diseases and Sciences,2007,52(9):2015-2021.
[33] SKRZYDLEWSKA E SULKOWSKI S,KODA M,et al.Lipid peroxidation and antioxidant status in colorec⁃tal cancer[J].World Journal of Gastroenterology,2005,11(3):403-406.
[34] HAMER H M,JONKERS D,VENEMA K,et al.Re⁃view article:the role of butyrate on colonic function [J].Alimentary Pharmacology&Therapeutics,2008,27(2):104-119.
[35] ABRAHAMSE S L,POOL⁃ZOBEL B L,RECHKEM⁃MER G.Potential of short chain fatty acids to modu⁃late the induction of DNA damage and changes in the intracellular calcium concentration by oxidative stress in isolated rat distal colon cells[J].Carcinogenesis,1999,20(4):629-634.
[36] TODEN S,BIRD A R,TOPPING D L,et al.Dose⁃de⁃pendent reduction of dietary protein⁃induced colono⁃cyte DNA damage by resistant starch in rats correlates more highly with caecal butyrate than with other short chain fatty acids[J].Cancer Biology&Therapy,2007,6(2):253-258.
[37] 张卫辉.丁酸钠对肉鸡免疫反应和抗氧化功能调控作用研究[D].博士学位论文.南京:南京农业大学,2011.
[38] HAMER H M,JONKERS D M A E,BAST A,et al.Butyrate modulates oxidative stress in the colonic mu⁃cosa of healthy humans[J].Clinical Nutrition,2009,28(1):88-93.
[39] STECKERT A V,VALVASSORI S S,VARELA R B,et al.Effects of sodium butyrate on oxidative stress and behavioral changes induced by administration of D⁃AMPH[J].Neurochemistry International,2013,62 (4):425-432.
[40] 陈禹银,刘凡.硅胶自水溶液中吸附丙酸和丁酸的计量置换吸附模型[J].物理化学学报,2002,18(1):62-65.
[41] 胡彩虹,朱康,钱仲仓,等.硅胶控释型丁酸对断奶仔猪生长性能和肠黏膜屏障的影响[J].动物营养学报,2011,23(12):2170-2176.
[42] 钱仲仓.控释型丁酸的制备及其对早期断奶仔猪肠屏障影响的研究[D].硕士学位论文.杭州:浙江大学,2010.
[43] 廖和菁,郑为完,易路遥,等.丁酸甘油酯的合成及其在油脂微囊化中的应用[J].食品科学,2009,30 (16):203-205.
[44] 丁斌鹰.三丁酸甘油酯在养猪生产上的应用研究进展[J].饲料工业,2012(增刊1):19-20.
[45] 王恬.仔猪断奶应激及营养调控措施的应用[J].畜牧与兽医,2009,41(5):1-4.
[46] 王继强,赵中生,龙强,等.早期断奶仔猪的生理特点及营养调控措施[J].饲料广角,2007(5):31-34.
[47] 刘统,黄兴国,朱广宁.丁酸甘油酯对断奶仔猪生长性能影响的研究[J].饲料工业,2011,32(22):39-41.
[48] 刘小萍,刘昌林,毛子平,等.丁酸甘油酯在仔猪生产应用中的初步研究[J].江西畜牧兽医杂志,2012 (2):15-16.
[49] 杨玲.三丁酸甘油酯对断奶仔猪生长性能、肠道结构和血液生化指标的影响[J].饲料工业,2012(增刊1):46-49.
[50] 王志刚,章利丰,侯嘉,等.微囊丁酸钠对保育猪生长性能的影响[J].中国饲料,2013(22):29-31.
[51] 陈国顺,徐振飞,赵芳芳,等.包膜丁酸钠对断奶仔猪生长性能和肠道消化酶活性的影响[J].国外畜牧学:猪与禽,2011,31(2):65-68.
[52] 吴胜莲,周映华,何万兵,等.丁酸钠对断奶仔猪生产性能的影响[J].饲料工业,2010,31(7):43-44.
[53] 钟翔,黄小国,陈莎莎,等.丁酸钠对断奶仔猪生长性能和肠道消化酶活性的影响[J].动物营养学报,2009,21(5):719-726.
[54] 李丹丹,冯国强,钮海华,等.丁酸钠对断奶仔猪生长性能及免疫功能的影响[J].动物营养学报,2012,24(2):307-313.
[55] 卢玉发,谢邦瑞.丁酸钠对断奶仔猪生长性能及血清生化指标的影响[J].黑龙江畜牧兽医,2008(7):42-43.
[56] 游金明,付建福,王自蕊,等.丁酸钠和甘露寡糖对断奶仔猪生长性能和免疫功能的影响[J].动物营养学报,2010,22(2):346-351.
[57] 侯嘉,赵民,余荣,等.微囊丁酸钠对泌乳母猪繁殖性能的影响[J].饲料研究,2013(10):60-62.
[58] 张小东,余荣,赵民.微囊丁酸钠对母猪生产性能与仔猪生长性能的影响[J].中国饲料,2012(5):14-16.
[59] 毛石明.包膜丁酸钠对母猪泌乳性能的影响[J].饲料研究,2010(3):32-33.
[60] 周磊,徐小芳,余荣,等.微囊丁酸钠对罗曼蛋种鸡生产性能的影响[J].中国畜牧杂志,2013,49(18):53-55.
[61] 赵民,徐小芳,余荣.包膜丁酸钠对青脚麻种鸡生产性能的影响[J].饲料工业,2012(20):33-34.
[62] 彭丽莎,孙健栋,史艳云,等.三丁酸甘油酯对肉鸡生长性能、养分表观消化率、屠宰性能、肠道形态及微生物菌群的影响[J].动物营养学报,2014,26(2):466-473.
[63] 邹杨,杨在宾,杨维仁,等.不同剂型丁酸钠与抗生素对肉仔鸡生产性能、肠道pH及挥发性脂肪酸含量的影响[J].动物营养学报,2010,22(3):675-681.
(责任编辑 武海龙)
Recent Advances on Application of Different Butyrate Products in Pigs and Poultry Production
LU Na WANG Tian ZHONG Xiang∗
(College of Animal Science and Technology,Nanjing Agricultural University,Nanjing 210095,China)
∗Corresponding author,associate professor,E⁃mail:zhongxiang@njau.edu.cn
Abstract:In recent years,butyrate products have been deeply applied to animal production as substitutes for feed antibiotic.This paper reviewed biological functions of butyrate,characteristics of different types of buty⁃rate and their application in pigs and poultry production.[Chinese Journal of Animal Nutrition,2015,27 (2):359⁃368]
Key words:butyrate products;pigs;poultry;application
通信作者:∗钟 翔,副教授,硕士生导师,E⁃mail:zhongxiang@njau.edu.cn
作者简介:芦 娜(1990—),女,天津人,硕士研究生,研究方向为动物营养与饲料科学。E⁃mail:akiran@sina.com
基金项目:国家自然科学基金(31201808,31472129);中央高校基本科研业务费自主创新重点项目(KYZ201313)
收稿日期:2014-09-02
doi:10.3969/j.issn.1006⁃267x.2015.02.005
文章编号:1006⁃267X(2015)02⁃0359⁃10
文献标识码:A
中图分类号:S816
猜你喜欢
临床神经外科杂志(2022年5期)2022-11-18
当代水产(2022年2期)2022-04-26
中国饲料(2021年17期)2021-11-02
当代水产(2021年2期)2021-03-29
中国粮油学报(2020年10期)2020-11-05
中国乳业(2018年11期)2018-12-13
江苏农业科学(2018年2期)2018-03-12
中国塑料(2016年2期)2016-06-15
烟草科技(2015年8期)2015-12-20
食品工业科技(2014年15期)2014-03-11
