
不同脂肪源饲料对津新鲤(Cyprinus carpio var.Jian)生长及脂类代谢的影响
2016-01-10 01:28方珍珍乔秀亭白东清
饲料工业 2016年8期
■刘 燕 方珍珍 曲 木 乔秀亭 白东清
(天津农学院水产学院天津市水产生态及养殖重点实验室,天津 300384)
植物油脂产量巨大,且价格相对鱼油稳定低廉,是一种潜在的甚至是唯一的能替代鱼油的脂肪源。很多研究也证明了植物油,如大豆油、亚麻油、菜籽油、橄榄油、棕榈油、玉米油等在水产饲料中的良好应用,并且在青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、团 头 鲂 (Mega-lobramaamblycephala)、黑 鲷 (Acanthopagrus schlegelii)、太 平 洋 鲑 (Oncorhynchus spp.)、虹鳟(Oncorhynchus mykiss)等品种上开展研究,发现在满足鱼体必需脂肪酸需求的前提下,其他油脂替代鱼油不仅不会对鱼体生长产生负面影响,而且适宜的替代还会有更好的促生长效果,但是不同脂肪源会对鱼类脂质代谢和抗氧化能力产生不同的影响。津新鲤是以建鲤为基础选育的新品种,具有抗寒能力强、繁殖力高、生长速度快和起捕率高等优点。关于津新鲤脂肪代谢,尤其是饲料中添加不同脂肪源对津新鲤脂肪代谢的关键酶FAS及LPL表达的影响却报道很少。当前,对替代鱼油脂肪源的研究大多数集中在鲑鳟鱼类,而很少涉及鲤科鱼类。本试验是以津新鲤为研究对象,探讨了不同脂肪源饲料对其生长及脂肪代谢酶活性和LPL、FAS基因表达的的影响,阐明津新鲤对脂肪的利用及转化能力,探索津新鲤饲料替代鱼油的最适脂肪源,进而降低饲料成本,提高饲料利用率,提升环境效益,同时也为进一步探索相关脂代谢酶的作用机理及鱼类脂肪代谢的分子调控机制的研究提供参考资料。
1 材料与方法
1.1 试验用鱼及饲养管理
本试验采用的津新鲤幼鱼由天津市换新水产良种场提供,试验在天津市天祥水产有限责任公司进行。在混养池塘上以泡沫浮板搭建18个1 m×1 m×2 m的沉性暂养网箱。试验鱼驯化适应环境7 d以后,选取体质健壮、规格一致、无伤无病,初始体重为(44.34±1.58)g,初始体长为(15.20±0.87)cm,随机分为6组,每组3个重复,每个重复70尾鱼。饲养过程中水温为(28±1.3)℃,pH值为(7.8±0.1),每日投喂2 次(8:30和15:00),日投喂率为体重的6%,养殖时间为8周。
1.2 试验设计
以鱼粉、豆粕、菜粕、花生粕和棉粕作为蛋白源,分别以豆油、鸡油、玉米油、棕榈油、菜籽油、棉籽油作为脂肪源,配制6种不同脂肪源的等氮、等能试验饲料,依次记为Diet 1、Diet 2、Diet 3、Diet 4、Diet 5、Diet 6,各饲料原料均通过粉碎机粉碎全部过40目分析筛,混合均匀后,使用江苏牧羊集团牧羊MUZLMV4型饲料制粒机制成直径为2.80 mm的沉性颗粒饲料,各组试验饲料的组成与营养水平见表1。
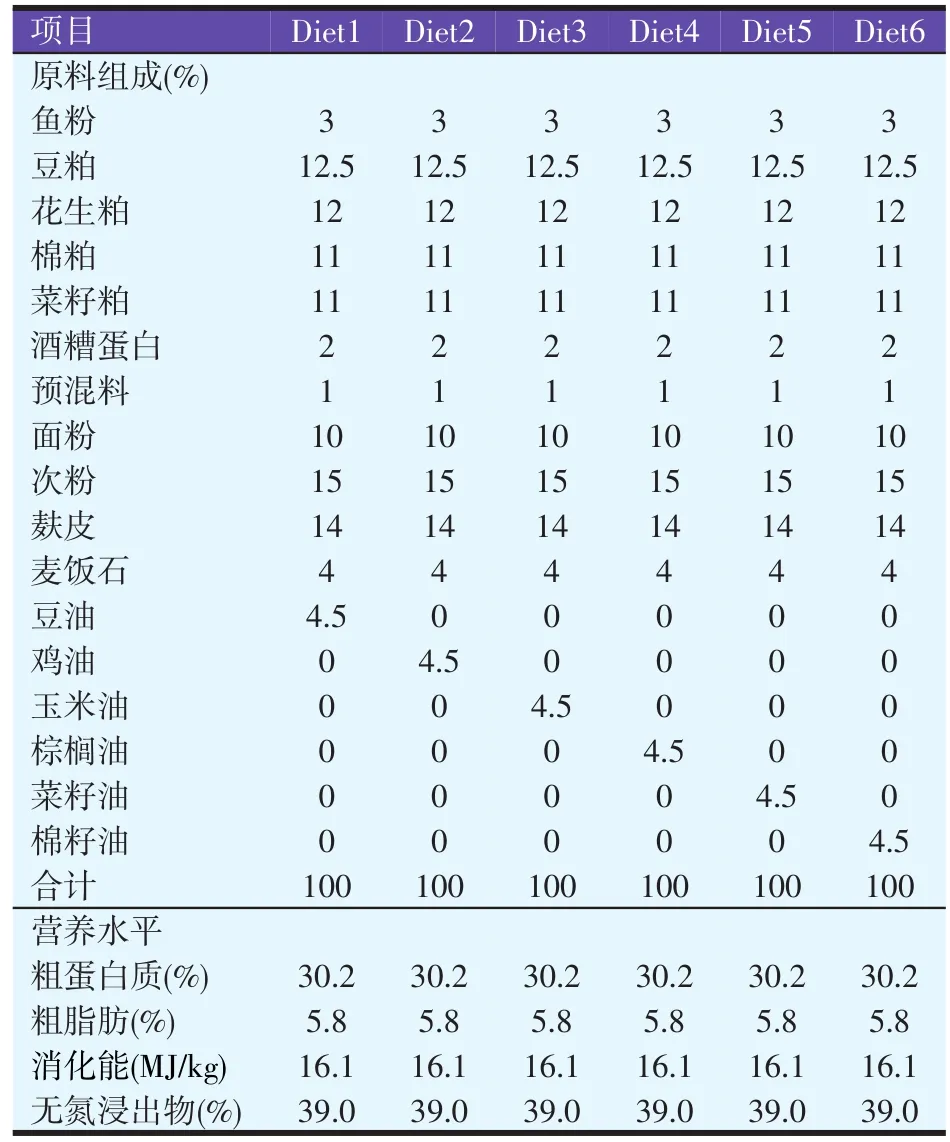
表1 试验饲料组成及营养水平(风干基础)
1.3 样本采集和分析
饲养结束后,禁食48 h,逐箱称重,每个重复中取10尾鱼解剖取出肝胰脏及肾脏并称肝胰脏重量,然后转入-20℃低温冰箱中保存,待测脂肪代谢酶活性。剩余的鱼再投喂后0、2、4、6、8 h,取每个重复中的9尾鱼的肝胰脏和肾脏在液氮中速冻,然后转入-80℃低温冰箱中保存,待测脂肪代谢酶基因表达。
生长试验开始和结束时,测定试验鱼体长和体重,计算存活率、增重率、特定生长率、饵料系数、蛋白质效率、肝体指数及肥满度。
存活率(%)=100 ×Nt/N0;
相对增重率(WGR,%)=100×(Wt-W0)/W0;
特定生长率(SGR,%/d)=100×[ln(Wt)-ln(W0)]/t;
饵料系数(FCR)=F/(Wt-W0);
蛋白质效率(PER,%)=100×(Wt-W0)/(F×P);
肝体比(HIS,%)=100×Wg/Wt;
肥满度(CF,%)=100×Wt/Lt3。
式中:Nt、N0——试验末期和初期鱼体总数;
W0——试验开始时鱼体重(g);
Wt——试验结束时鱼体重(g);
F——饲料摄入量干重(g);
P——饲料中粗蛋白含量(%);
Wg——肝胰脏质量(g);
Lt——试验末鱼体长(cm);
t——饲养天数(d)。
肝脂酶(HL)、脂蛋白脂酶(LPL)、总脂酶(GE)(其中总脂酶活性=脂蛋白脂酶活性+肝脂酶活性)和脂肪酸合成酶(FAS),其测定所需试剂盒均购自南京建成生物工程研究所。
1.4 数据统计与分析
数据均以“X±SD”表示,采用SPSS18.0对所得数据进行单因素方差分析(one-way ANOVA),若差异达到显著水平,则采用Duncan's法进行多重比较,显著性水平P<0.05。
1.5 组织中FAS及LPL mRNA表达的测定
1.5.1 引物设计
根据Genbank中现有的其他鱼类的FAS和LPL保守序列设计引物,根据鱼类β-actin保守序列,设计津新鲤β-actin引物(见表 2)。
1.5.2 总RNA的提取和cDNA的第一链合成
总RNA提取采用RNAiso Plus试剂,根据说明书进行RNA抽提。使用微分光光度计检测总RNA纯度和定量OD260/280值。使用PrimeScriptTM1stStand cDNA Synthesis Kit(大连宝生物工程有限公司)将提取的肝胰脏及肾脏总RNA反转录成 cDNA,以Oligo(dT)为引物,反应条件为:50 ℃,45 min,70 ℃,15 min,合成的cDNA第一链保存于-20℃冰箱中。

表2 本研究所用的引物序列
1.5.3 PCR扩增
PCR反应每一样品扩增体系为25 μl,其中10×PCR Buffer 2.5 μl,dNTP Mixture 1.5 μl,TaKaRa TaqE 0.5 μl,引物 F 0.5 μl,引物R 0.5 μl,双蒸水 18.5 μl,cDNA模板1 μl。
FAS cDNA片段扩增反应条件为94℃预变性10 min;94℃变性30 s,52℃退火45 s,72℃延伸1 min,35个循环;最后72℃延伸10 min。LPL cDNA片段扩增反应条件为94℃预变性10 min;94℃变性40 s,53 ℃退火40 s,72 ℃延伸1 min,30个循环;最后72℃延伸10 min。β-actin cDNA片段扩增反应条件为94℃预变性10 min;94℃变性45 s,59℃退火30 s,72℃延伸1 min,30个循环;最后72℃延伸10 min。
1.5.4 组织中FAS mRNA及LPL mRNA相对表达量的数据统计分析
利用Gel-Pro analyzer软件分析电泳图像中各条带的信号强度,用FAS及LPL信号强度分别与相对应 β-actin信号强度的比值表示FAS mRNA及LPL mRNA的相对表达量,并根据FAS/β-actin及LPL/βactin比值,对FAS mRNA及LPL mRNA的相对表达量进行统计分析。
2 结果
2.1 不同脂肪源饲料对津新鲤生长和饲料利用的影响(见表3、表4)

表3 不同脂肪源饲料对津新鲤生长和饲料利用的影响

表4 不同脂肪源饲料对津新鲤生长的影响(%)
由表3可知,饲养8周后,各组津新鲤存活率组间差异不明显(P>0.05)。相对增重率与特定生长率均以Diet 3组为最高,显著高于Diet 5 组(P<0.05),但与其他4组相比变化不明显(P>0.05)。Diet 3组饵料系数最低,显著低于Diet 4 组,但与Diet 1、2、5、6处理组相比变化不大(P>0.05)。从蛋白质效率上看,Diet 3组最高,达2.76%,显著高于Diet 2 组(P<0.05)。
由表4可知,添加豆油、鸡油、玉米油、棕榈油、菜籽油、棉籽油6种脂肪源对津新鲤肥满度影响不大(P>0.05)。但对津新鲤的肝体比影响较大,Diet 5组肝体比最高,显著高于其他各组(P<0.05);其次是Diet 3和Diet 4组,这两组间差别不大(P>0.05);Diet 2组肝体比最低,显著低于Diet 3~5组(P<0.05)。
可见,从生长指标来看,添加玉米油的饲料组效果最好。
2.2 不同脂肪源饲料对津新鲤相关脂代谢酶活性的影响
2.2.1 不同脂肪源饲料对津新鲤肝胰脏中脂代谢酶活性的影响(见表5)

表5 不同脂肪源饲料对津新鲤肝胰脏中脂代谢酶活性的影响(U/g)
由表5可知,添加玉米油组津新鲤肝胰脏中脂蛋白脂酶、肝脂酶和总脂酶活性显著高于其余各组(P<0.05)。脂肪酸合成酶活性Diet 1、Diet 2、Diet 3各组间差异不明显(P>0.05);Diet 4、Diet 5、Diet 6 组间差异不显著(P>0.05)。
2.2.2 不同脂肪源饲料对津新鲤肾脏中脂代谢酶活性的影响(见表6)
由表6可知,肾脏脂蛋白脂酶、肝脂酶、总脂酶活性因添加不同脂肪源而不同,Diet 3组显著高于其他各组(P<0.05),而Diet 4、Diet 5、Diet 6组的3种酶活性间无显著差异(P>0.05)。津新鲤肾脏中脂肪酸合成酶活性 Diet 1、Diet 2、Diet 3各组间差异并不显著(P>0.05);Diet 4、与Diet 5与 Diet 6组间脂肪酸合成酶活性差异不显著(P>0.05)。

表6 不同脂肪源饲料对津新鲤肾脏中脂代谢酶活性的影响(U/g)
2.3 不同脂肪源饲料对津新鲤肝胰脏及肾脏中FAS及LPL mRNA表达的影响
2.3.1 津新鲤总RNA的提取及检测结果
本试验提取的RNA经过微分光光度计检测总RNA纯度,其定量OD260/280值均在1.8~2.0之间,表明纯度高、污染低。提取的总RNA样品经1%琼脂糖凝胶电泳检测呈现三条带,结果见图1。2.3.2 津新鲤FAS、LPL和β-actin在不同组织中的表达(见图2、图3)

图1 津新鲤总RNA的琼脂糖凝胶电泳图
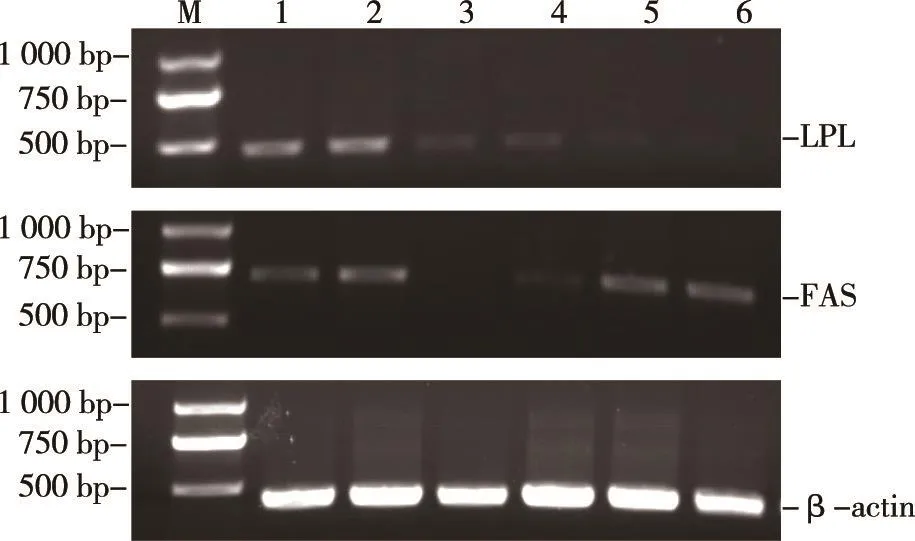
图2 津新鲤各组织中FAS及LPL基因的表达
采用半定量RT-PCR方法,对津新鲤各组织的FAS及LPL mRNA表达情况进行检测,由图2和图3可知,FAS在肾脏、肝胰脏中表达丰度相对较高,在脾脏中表达丰度较低;LPL在肾脏、肝胰脏中表达丰度相对较高,在肠道中表达丰度较低。
2.3.3 不同脂肪源饲料对津新鲤肝胰脏和肾脏中FAS及LPL mRNA表达丰度的影响
本试验根据相关脂代谢酶活性因饲料中添加不同脂肪源呈现的变化趋势,检测了禁食48 h及再投喂不同脂肪源饲料后的2、4、6、8 h时津新鲤肝胰脏及肾脏中FAS及LPL mRNA表达水平,并作了显著性分析。

图3 津新鲤各组织中FAS和LPL基因的RT-PCR扩增产物
2.3.3.1 不同脂肪源饲料对津新鲤肝胰脏LPL mRNA表达丰度的影响(见表7)

表7 禁食与投喂不同脂肪源饲料后津新鲤肝胰脏LPL mRNA水平的变化
由表7可知,投喂不同脂肪源饲料之后,津新鲤肝胰脏LPL mRNA表达丰度呈现先上升后下降的趋势,在禁食水平下最低,6 h达到峰值,并且高于其它时间段,随后开始下降,趋于稳定,达到禁食水平。总体来看,Diet 3组LPL mRNA表达丰度显著高于其它各组(P<0.05)。
2.3.3.2 不同脂肪源饲料对津新鲤肝胰脏FAS mRNA表达丰度的影响(见表8)
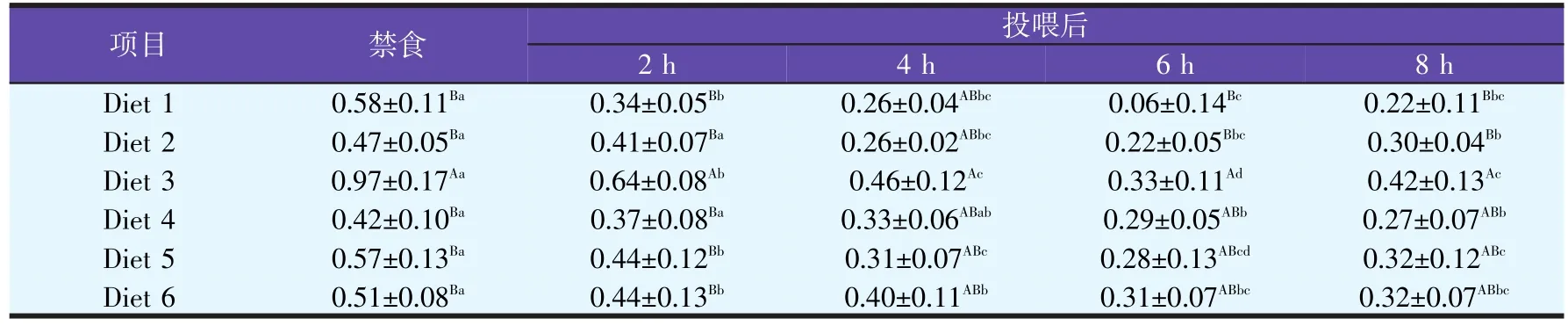
表8 禁食与投喂不同脂肪源饲料后津新鲤肝胰脏FAS mRNA水平的变化
由表8可知,投喂不同脂肪源饲料之后,津新鲤肝胰脏FAS mRNA表达丰度呈现先下降后上升的趋势,6 h达到最低,并且低于其它时间段,随后开始上升,趋于稳定,达到禁食水平。总体来看,Diet 3组FAS mRNA表达丰度高于其它各组,但无显著性差异(P>0.05)。2.3.3.3 不同脂肪源饲料对津新鲤肾脏LPL mRNA表达丰度的影响(见表9)
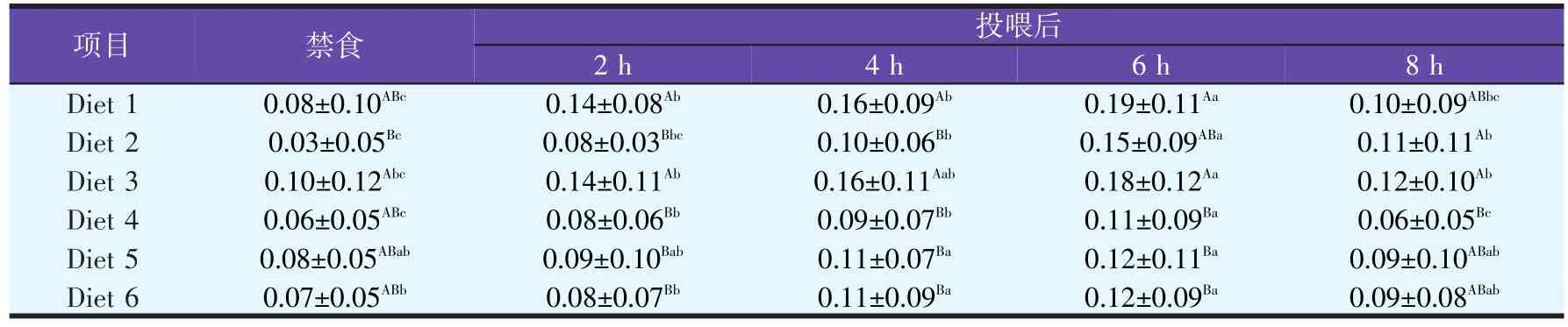
表9 禁食与投喂不同脂肪源饲料后津新鲤肾脏LPL mRNA水平的变化
由表9可知,投喂不同脂肪源饲料之后,津新鲤肾脏LPL mRNA表达丰度呈现先上升后下降的趋势,在禁食水平下最低,6 h达到峰值,并且高于其它时间段,随后开始下降,趋于稳定,达到禁食水平。总体来看,Diet 3组LPL mRNA表达丰度高于其它各组,但无显著性差异(P>0.05)。
2.3.3.4 不同脂肪源饲料对津新鲤肾脏FAS mRNA表达丰度的影响(见表10)

表10 禁食与投喂不同脂肪源饲料后津新鲤肾脏FAS mRNA水平的变化
由表10可知,投喂不同脂肪源饲料之后,津新鲤肾脏FAS mRNA表达丰度呈现逐渐下降的趋势,禁食水平下FAS基因表达上调到最高。总体来看,虽然Diet 3组FAS mRNA表达丰度高于其它各组,但变化不大(P>0.05)。
3 讨论
3.1 不同脂肪源饲料对津新鲤生长性能的影响
Arzel等研究鱼油和玉米油的添加量对褐鳟生长、机体营养组成的影响,结果显示:玉米油组褐鳟成活率比鱼油组偏高。Rainuzzo等认为,不同的水产动物对饲料中不同脂肪原料的利用有较大差异;雍文岳等研究发现:大豆油为脂肪源的饲料对草鱼的生长作用明显好于猪油;不同鱼类对日粮中不同脂肪原料的利用也有较大的差异。Kamarudin等对吉罗鱼研究表明,与葵花籽油和亚麻籽油相比,棕榈油的生长效果最好。对鲤、罗非鱼、大口鲈等鱼研究发现不同脂肪源对鱼类生长及饲料利用没有显著影响。不同脂肪源对鱼类的生长没有影响,究其原因可能是由于试验条件或鱼种类不同,如蛋白源鱼粉保证了一定量的HUFA来满足鱼类快速生长,而本试验饲料蛋白源主要由豆粕、花生粕和棉粕等提供,仅含少量的鱼粉,结果显示,添加玉米油的饲料组的相对增重率、特定生长率和蛋白质效率较高,饵料系数最低,津新鲤生长效果最佳。这可能是由于不同脂肪源的脂肪酸组成不同,被鱼类所利用的方式和能力不同,对鱼类生长、代谢等的影响因而也有所差别。
冯健等研究发现,太平洋鲑玉米油组的肥满度显著高于大豆油组和鱼油组;杜震宇等发现添加大豆油、玉米油对鲈鱼肥满度的影响不显著。本试验中不同脂肪源对津新鲤肥满度影响差异不显著。不同脂肪源对鱼体肥满度的影响不完全相同,可能是与脂肪源的种类、添加量、鱼的种类和大小有关。鱼类肝体比为对长期和短期营养方式都很敏感的指标。Bell等研究发现,菜籽油组大西洋鲑的肝体比显著高于鱼油组。本试验中津新鲤的肝体比以添加玉米油的饲料组最大,但并未达到明显脂肪肝的程度。
3.2 不同脂肪源饲料对津新鲤相关脂代谢酶活性的影响
肝脏是鱼类进行脂肪酸代谢的主要场所,肝脂酶(HL)和脂蛋白脂酶(LPL)是鱼类脂肪酸分解代谢过程中的两个关键酶,脂肪酸合成酶(FAS)在脂肪合成过程中起关键作用。LPL、FAS和HL三者在脂类代谢过程中发挥着重要作用。已有研究发现,饲料中适宜的脂肪含量可提高饲料利用率和脂代谢效率。饲料营养因素影响FAS活性高低,如Dias等研究发现,高豆粕饲料能够使欧洲鲈鱼肝脏中FAS活性下降;许多研究表明,饲料脂肪水平的升高会抑制FAS的活力及FAS mRNA的表达丰度,但对LPL的活力和基因表达有促进作用。津新鲤摄食高脂饲料后,甘油三酯含量上升,机体代谢旺盛,需要大量的LPL,从而促进了肝胰脏LPL的分泌。LPL的活性和基因表达受多方面因素影响,例如鱼的种类、水温、胰岛素、季节等。当前,对于不同脂肪源对鱼类肝胰脏LPL活性影响的研究包括舌齿鲈和异育银鲫等鱼类,研究结果也因养殖品种不同而有所差异。
在本试验中,随着不同脂肪源的添加,津新鲤肝胰脏中LPL、HL和GE活性呈显著变化,且均为添加玉米油的饲料组最大,显著高于其余各试验组(P<0.05)。肾脏中LPL、HL、GE活性随着不同脂肪源的添加也有变化趋势,同样添加玉米油饲料组显著高于其他各组(P<0.05)。津新鲤肝胰脏及肾脏中FAS活性有些组之间的差异并不显著(P>0.05),这可能是由于脂肪的来源不同,其饱和脂肪酸、单多不饱和脂肪酸含量也有所不同,对鱼体的消化吸收也有显著不同。目前,关于鱼类肾脏与脂质代谢之间关系的研究相对较少,其具体调节机制尚待进一步研究。
3.3 不同脂肪源饲料对津新鲤组织中FAS及LPL mRNA表达的影响
脂肪酸合成酶(FAS)和脂蛋白脂酶(LPL)是动物体内脂肪代谢的关键酶。FAS作为脂肪酸合成酶系,其含量的多寡、活性的高低对控制动物体脂的生成、沉积发挥着重要作用。饲料脂肪水平提高了黑鲶鱼和红鲷肝脏中LPL表达,而降低了吉富罗非鱼肝胰脏中FAS表达。与本试验中不同脂肪源饲料诱导和调节了津新鲤肝胰脏中LPL表达并抑制和调节FAS表达的结果相呼应。然而Liang等的研究发现,饲料脂肪水平并没有显著提高红鲷肝胰脏中LPL mRNA表达,这可能与鱼的品种、大小及投喂时间长短有关。
采用半定量RT-PCR方法,检测了津新鲤FAS及LPL mRNA在各组织中表达分布,结果发现FAS在肾脏、肝胰脏中表达丰度较高,在脾脏中表达丰度较低;LPL在肾脏、肝胰脏中表达丰度较高,在肠道中表达丰度较低。采用同样方法对禁食后48 h及再投喂后的2、4、6 h和8 h肝胰脏、肾脏中FAS及LPL mRNA表达量进行测定分析,以表明禁食后再投喂不同脂肪源对津新鲤肝胰脏及肾脏中FAS及LPL mRNA表达水平的影响。
投喂不同脂肪源饲料之后,津新鲤肝胰脏和肾脏LPL mRNA表达丰度均呈现先上升后下降的趋势,在禁食水平下最低,6 h达到峰值,并且高于其它时间段,随后开始下降,趋于稳定,达到禁食水平。王煜恒等的研究结果也证实了不同脂肪源对异育银鲫肝胰脏LPL活性产生不同的影响。这可能是由于随着脂肪酸不饱和度的增加脂肪酸促进LPL的基因mRNA表达的作用加强,进而诱导肝胰脏LPL合成增加所致。投喂不同脂肪源饲料之后,津新鲤肝胰脏FAS mRNA表达丰度呈现先下降后上升的趋势,6 h达到最低,并且低于其它时间段,随后开始上升,趋于稳定,达到禁食水平。而肾脏FAS mRNA表达丰度呈现逐渐下降的趋势,禁食水平下最高。这些结果表明肝胰脏中FAS及LPL mRNA表达量受到禁食或者再投喂的调节。以上试验结果与吉红等的试验结果一致,与王爱民等和Oku等部分结果一致。造成这种差异可能的原因是由于禁食时间和再投喂时间的不同或者试验鱼种不同。至于分子调节机制,还需进一步研究。
4 结论
①不同脂肪源饲料会影响津新鲤的生长性能、脂肪酶活性及LPL、FAS基因的表达。
②通过添加等量豆油、鸡油、玉米油、棕榈油、菜籽油和棉籽油作为脂肪源,得出添加玉米油的饲料组的相对增重率、特定生长率和蛋白效率较高,饵料系数最低,津新鲤生长效果最佳,说明玉米油作为单一脂肪源可以提高饲料营养物质的利用率。
③通过不同脂肪源饲料对津新鲤相关脂代谢酶活性的影响分析,津新鲤肝胰脏及肾脏的脂蛋白脂酶、肝脂酶和总脂酶活性以添加玉米油的饲料组最佳。
④饲料中添加玉米油更容易诱导津新鲤肝胰脏及肾脏LPL活性及LPL mRNA的表达,而对FAS活性及FAS mRNA表达的影响恰恰相反。
(参考文献41篇,刊略,需者可函索)
猜你喜欢
中国食品(2021年11期)2021-06-23
现代临床医学(2021年2期)2021-03-29
医学与法学(2020年3期)2020-09-18
科学(2020年2期)2020-08-24
科学(2020年2期)2020-01-08
意林·全彩Color(2019年9期)2019-10-17
科学与财富(2018年8期)2018-05-09
广东饲料(2016年2期)2016-12-01
现代食品(2016年14期)2016-04-28
中国粮油学报(2016年1期)2016-02-06
