
遮荫对5种园林植物光合特性的影响
2015-12-21 03:25:38宋立天
草原与草坪 2015年3期
宋立天
(深圳市市政设计研究院有限公司,广东 深圳 518000)
研究植物耐荫性是选择绿地植物和立体优化配置的基础,也是充分利用荫生环境空间、改善立体绿化植物种类单一现状、提高绿化效果的有效途径[1-3]。随着我国城市化进程的加快,荫生环境面积快速增加,荫生环境的绿化问题日显突出,植物的耐荫性研究也因此逐步深入[4,5]。
光合作用是植物生长发育的基础,20世纪初,国外已开始关注植物的耐荫性和光合作用机理,并开展了一系列生理生化研究。主要包括植株形态、叶片解剖结构、叶绿体及叶绿素分布、叶绿素荧光特性光电子传递途径及相关酶的活性等[6-12]。我国对植物耐荫性的研究始于20世纪70年代,研究范围主要集中在植株形态特征、叶绿素含量和光合作用响应曲线等方面,对引进和筛选耐荫性园林植物还有缺陷,在城市园林绿化中仍然存在许多植物群落配置不科学的现象[13]。
通过引入一批耐荫植物品种,进行生物学特性观察及耐荫机理研究,筛选出观赏价值高、耐荫性好的植物品种,以期为今后开发利用耐荫植物资源、营建稳定的生态植物群落提供理论依据。
1 材料和方法
1.1 试验地概况
试验地设在深圳职业技术学院,E 114°04′12″,N 22°37′12″,属亚热带季风气候,年均气温22.4℃,年均降水量1 948mm。土壤为赤红壤。
1.2 试验材料
(1)收集具有一定耐荫性的初试植物:通过查阅文献资料[14-17],并在华南地区实地调查,共收集30种具有一定耐荫性的园林植物(表1)。
(2)试验品种的筛选:在遮荫60%的条件下对30种具有一定耐荫性的植物进行2个月的预试验,从中选出了15种易成活、观赏价值高的园林植物,再探讨遮荫对其生物量和生长发育、叶片解剖结构、叶绿素含量及景观特征等指标的影响[18,19],进而筛选出阔叶凤尾蕨、珊瑚花、鸟尾花、叉花草、江南星蕨5种园林植物,作为专门研究遮荫对光补偿点、光饱和点及光合速率等项指标影响。

表1 耐荫性的园林植物名录Table1 Pretested garden plants with shading tolerance
(3)仪器设备:LI-6400型光合仪和 BAU 光合测定系统[20,21]。
1.3 试验处理与步骤
(1)试验处理:试验设对照(全光照)和3个遮荫处理,用黑色遮荫网搭建不同规格的遮荫棚。依据对荫生环境下测定的光照强度设置60%,75%和85%3个光照处理,依次用TR1,TR2和TR3来表示,每处理9个重复[22,23]。
(2)测定方法:遮荫180d后于2013年12月选取3d(天气晴朗、光照充足)在野外直接测定试验指标。每天分别在7∶00~17∶00,每隔2h测定1次。每品种选3个样株,每样株选取上端第2轮功能叶(发育完整,无病虫害侵染),每株选3片叶重复测定。每个叶片重复3次,取平均值。测定净光合作用值、净光合速率(Pn)、蒸腾速率(Tr)、光能利用效率等指标。以3d测量平均值为叶片气体交换参数的值。水分利用效率(WUE)用光合速率与蒸腾速率之比计算。光能利用率用净光合速率与光合有效辐射(PAR)的比值计算[24]。
1.4 数据统计分析
运用Excell和SPSS进行数据统计和相关性分析。
2 结果与分析
2.1 5种园林植物净光合速率的比较
净光合速率是指植物的总光合量减去呼吸作用所消耗的量[25]。珊瑚花和江南星蕨在遮光60%的环境下净光合速率最大,鸟尾花和叉花草在75%的条件下净光合速率值最大,而珊瑚花在遮荫85%的条件下净光合速率值最大。结果表明:85%遮荫净光合速率5种园林植物耐荫性的顺序是,珊瑚花>阔叶凤尾蕨>叉花草>鸟尾花>江南星蕨(图1)。

图1 不同遮荫条件下5种园林植物净光合速率Fig.1 Variation of net photosynthetic rate of five garden plants under different shading levels
2.2 5种园林植物光合有效辐射与净光合速率比较
光合有效辐射是绿色植物在光合作用过程中,吸收太阳辐射中使叶绿素分子呈激发状态的那部分光谱能量。光合速率则反映不同植物的光合生物学特性及不同强度的有效光合辐射对植物净光合速率的影响[26]。随光源的增强,鸟尾花等5种园林植物的净光合速率增加,并在有效光辐射为50~100 μmol/(m2·s)时净光合速率迅速增加,此后增速明显降低,其中,江南星蕨和阔叶凤尾蕨2种蕨类植物的净光合速率在1 200μmol/(m2·s)时达到最大值,两者的光饱和点在1 200μmol/(m2·s),而鸟尾花、珊瑚花和叉花草的净光合速率持续上升,其光饱和点接近1 500μmol/(m2·s)。进一步分析可知,江南星蕨和阔叶凤尾蕨的光补偿点很低,分别为10.25和9.35μmol/(m2·s),为喜荫植物,其他3种光补偿点也比较低,依次为 32.66,16.95 和 18.78 μmol/(m2·s),具有一定的耐荫性(图2)。
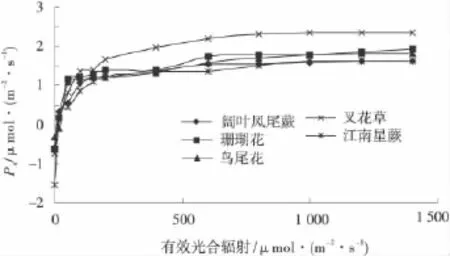
图2 5种园林植物光响应曲线Fig.2 Light-response curve of five garden plants
2.3 光能利用率日变化
2.3.1 60%遮荫下5种园林植物的光能利用率日变化 60%遮荫下9∶00~15∶000的光能利用率并无很大的差别。其中,阔叶凤尾蕨和珊瑚花在7∶00~9∶00光能利用率逐渐升高,9∶00~15∶00趋于平稳,15∶00~17∶00逐渐升高;鸟尾花在7∶00~9∶00的光能利用率逐渐升高,9∶00~15∶00趋于平稳,15∶00~17∶00再次逐渐升高;叉花草在7∶00~9∶00光能利用率逐渐升高,9∶00~13∶00有下降的趋势,13∶00~15∶00再次逐渐升高,15∶00~17∶00则大幅上升;江南星蕨7∶00~11∶00光能利用率逐渐升高,11∶00~13∶00逐渐下降,13∶00~15∶00逐渐回升,15∶00~17∶00则大幅上升。表明江南星蕨利用光的能力很强。

图3 60%遮荫下的5种园林植物的光能利用率日变化Fig.3 Daily variation of light utility efficiency of five garden plants at 60%of shading level
2.3.2 85%遮荫下5种园林植物的光能利用率日变化 85%遮荫下7∶00~13∶00光能利用率没有大的区别(图4)。5种园林植物都在13∶00~15∶00光能利用率逐渐上升,15∶00~17∶00逐渐下降,并在15∶00达到峰值;但峰值差异显著,自大至小顺序为:江南星蕨>阔叶凤尾蕨>鸟尾花>叉花草>珊瑚花,表明江南星蕨和阔叶凤尾蕨利用弱光的能力强,而其余3种园林植物也具有一定的利用弱光能力。

图4 85%遮荫下的5种园林植物的光能利用率日变化Fig.4 Daily variation of light utility efficiency of five garden plants at 85%of shading level
2.4 净光合速率的日变化
2.4.1 60%遮荫下5种园林植物的净光合速率的日变化 5种园林植物的净光合速率均呈现双峰值曲线(图5),分别在11∶00和15∶00呈现一个峰值,其中,珊瑚花、叉花草和鸟尾花11∶00的峰值远大于15∶00的峰值,而江南星蕨和阔叶凤尾蕨的峰值下午15∶00的较大,在上午9∶00~11∶00,前3种园林植物的净光合速率高于后2种园林植物,但在早晚时,江南星蕨和阔叶凤尾蕨的净光合速率则高于其他3种园林植物,表明江南星蕨和阔叶凤尾蕨2种园林植物的光利用能力高于其他植物,这与前面的结果一致。同时,5种园林植物在13∶00都会出现一个“午休”现象,此时植物的净光合速率下降到 “谷值”。

图5 60%遮荫度下5种园林植物净光合速率日变化Fig.5 Daily variation of net photosynthetic rate of five garden plants at 60%of shading level
2.4.2 85%遮荫下5种园林植物的净光合速率的日变化 85%遮荫的条件下的净光和速率变化趋势与60%遮荫相似。在8∶00以前鸟尾花、珊瑚花和叉花草的净光合速率均为负数,而阔叶凤尾蕨和江南星蕨的净光合速率均为正数,到17∶00以后2者的净光合速率高于其他3种园林植物,这也表明阔叶凤尾蕨和江南星蕨利用弱光的能力强于其他3种植物。鸟尾花、珊瑚花和叉花草在遮荫85%条件下的净光合速率明显低于遮光60%条件下的,相反,阔叶凤尾蕨和江南星蕨的净光合速率相差无几,表明阔叶凤尾蕨和江南星蕨在遮荫85%的环境下依然能很好地积累有机质。而珊瑚花、鸟尾花、叉花草在遮荫85%的环境下尽管也能积累一定的有机质,但其净光合速率明显低于遮荫60%环境下的。这些变化表明其最适宜在遮荫60%的环境下生长,这一结论与植物引种栽培观察的结果及叶片解剖结构结论相一致。
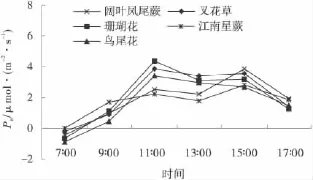
图6 85%遮阴下5种园林植物净光合速率日变化Fig.6 Daily variation of net photosynthetic rate of five garden plants at 85%of shading level
2.5 蒸腾速率(Tr)的日变化
2.5.1 60%遮荫下园林植物的蒸腾速率(Tr)的日变化 植物通过蒸腾作用运输矿物质、调节叶面温度、供应光合作用所需要的水分等,与植物净光合速率高度相关[27,28]。各种植物的蒸腾速率的变化差异较大(图7),江南星蕨呈现先降后升,最后达到一个平稳值,在13∶00的时候达到最低值,为0.111μmol/(m2·s),可以看出此时江南星蕨正处在“休眠”期,以保证自身的水分散失。阔叶凤尾蕨的蒸腾速率呈现波浪式曲线,在11∶00达到最低值,为0.244μmol/(m ·s),15∶00达到峰值0.317μmol/(m2·s)。珊瑚花的蒸腾速率变化值呈现单峰曲线,在15∶00达到了最大值0.256 μmol/(m2·s)。叉花草的蒸腾速率变化呈现单峰曲线,且在13∶00达最大值0.257μmol/(m2·s),其中,以江南星蕨蒸腾速率变化范围最大。

图7 60%遮荫度下园林植物的蒸腾速率日变化Fig.7 Daily variation of transpiration rate of five garden plants at 60%of shading level
2.5.2 85%遮荫下园林植物的蒸腾速率的日变化85%遮荫条件下,各种植物的蒸腾速率变化值差异较大(图8)。其中,阔叶凤尾蕨呈现单峰式变化曲线,并在15∶00时达到最大值0.343μmol/(m2·s);珊瑚花呈现波浪式变化曲线,在9∶00时达到最低值0.136 μmol/(m2·s),在11∶00时达到第一个峰值0.279 μmol/(m2·s),15∶00时达到第2个峰值0.231 μmol/(m2·s)。叉花草蒸腾速率变化呈现波浪式曲线,在15∶00时达到最大值0.332μmol/(m2·s),在15∶00以后蒸腾速率快速下降。江南星蕨蒸腾速率呈现“凹”形变化曲线,并在15∶00达到最低值0.122 μmol/(m2·s)。综合分析,叉花草在85%遮荫条件下蒸腾速率的变化范围最大。
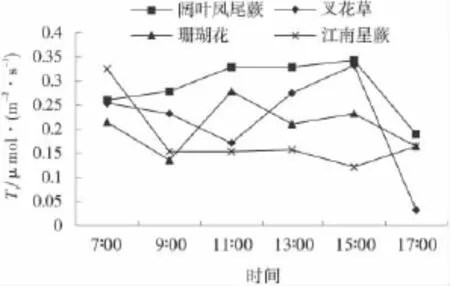
图8 85%遮荫度下园林植物蒸腾速率日变化Fig.8 Daily variation of transpiration rate of five garden plants at 85%of shading level
3 讨论与结论
光合生理指标主要有光补偿点、光饱和点、最大表观量子效率以及光饱和时的光合速率等,是影响植物光响应曲线的重要参数。利用光响应曲线来评价植物的耐荫性是世界公认的。一般来说,植物的光补偿点越低、表观量子效率越大,说明该植物在弱光下能捕获更多的光能,顺利进行光合作用和累积光合产物,因此,植物具有较强的耐荫能力。反之,耐荫能力则弱。相关研究表明,若植物的光补偿点和光饱和点越低,则该植物的耐荫能力就越强;若植物的光补偿点低,光饱和点高,则说明该种植物的光适应较强,较耐荫;若植物的光补偿点和光饱和点都高,则说明该植物属阳性植物[12,14,18,28]。
试验结果表明,江南星蕨、阔叶凤尾蕨利用光能的效率高,特别是利用低光的能力强,在遮荫85%的条件下能获得最大或接近最大的净光合作用值。其光补偿点很低,是典型的耐荫植物。鸟尾花、叉花草和珊瑚花的光饱和点和光补偿点也比较低,表明这3种植物具有一定的耐荫性,适度遮荫有利于其进行光合作用,它们光合作用的最大值出现在遮荫60%的条件下,这3种植物很适宜在遮荫60%的环境下生长,这一结论和引种栽培观察试验及叶片结构解剖分析的结果基本相同。
(1)研究不同有效光合辐射与净光合速率变化发现,江南星蕨和阔叶凤尾蕨的光补偿点分别是10.25 μmol/(m2·s)和9.35μmol/(m2·s),为喜荫植物,而鸟尾花、叉花草和珊瑚花的光补偿点依次是32.66,18.78和16.95μmol/(m2·s),表明它们也具有一定的耐荫性。
(2)比较60%和85%遮荫下光能利用率日变化表明,在60%遮荫下江南星蕨利用光的能力很强;在85%遮荫下江南星蕨和阔叶凤尾蕨利用弱光的能力也比较强,说明二者都是耐荫性很强的植物。而珊瑚花、鸟尾花和叉花草也具有不同程度的利用弱光能力。
(3)净光合速率的日变化表明,在60%和85%遮荫下江南星蕨和阔叶凤尾蕨2种植物的光利用能力高于其他植物,能很好地积累有机质,其余3种植物也能积累一定的有机质,但其净光合速率明显低于遮荫60%的净光合作用值。
(4)比较5种植物蒸腾速率的日变化,在60%遮荫下5种植物以江南星蕨的蒸腾速率值变化范围最大;在85%遮荫下叉花草蒸腾速率的变化范围最大。
[1] 周潇,毛凯,干友民.我国地被植物耐荫性研究[J].北方园艺,2007(1):51-53.
[2] 安锋,林位夫.植物耐荫性研究的意义与现状[J].热带农业科学,2005,25(2):68-72.
[3] 余森文.园林植物配置艺术的探讨[J].建筑学报,1984(1):35-40.
[4] 苏雪痕.园林植物耐荫性及其配置[J].北京林业大学学报,1981(6):63-71.
[5] Masakazu Iwai.Distinct physiological responses to a high light and low CO2environment revealed by fluorescence quenching in photo-autotrophieally grown Chlamydomonas reinhardtii[J].Photosynth Res,2007,94:307-314.
[6] 周玉迁,李永辉,李滨胜,等.卵叶玉簪耐荫性的研究[J].生态农林,2010(14):53-75.
[7] 刘欣欣,张明如,邹伶俐,等.浙江省15个树种苗期叶片解剖结构特征比较分析[J].浙江农林大学学报,2013,30(4):484-489.
[8] Paquette A,Boucharda A,Cogliastroa A.Morphological plasticity in seedlings of three deciduous species under shelterwoed under-planting management does not correspond to shade tolerance ranks[J].Forest Ecol Manag,2007,241:278-87.
[9] Oguchi R.Does the photosynthetic light acclimation need change in leaf anatomy[J].Plant,Cell and Environment,2003,26:505-512.
[10] 游文娟,张庆费,夏檑.城市绿化植物叶片结构对光强的响应[J].西北林学院学报,2008,23(5):22-25.
[11] 刘宝臣,唐伟斌.遮荫对麦冬和玉簪叶面积及叶绿素含量的影响[J].北方园艺,2012(14):77-79.
[12] 张玲慧.地被植物耐荫性研究及园林配置探讨[D].杭州:浙江大学,2004
[13] 伍世平,于志熙.11种地被植物的耐荫性研究[J].武汉植物学研究,1994,12(4):360-364.
[14] 黄笛,吴铁明.植物耐荫生理研究进展[J].湖南林业科技,2011,38(4):61-63.
[15] 吴宗萍.遮荫对头花蓼形态和生理指标的影响[J].西南师范大学学报,2010,35(3):85-89.
[16] 王忠.植物生理学[M].北京:中国农业出版社,1999:126-131.
[17] 李兰芳,包维楷.植物叶片形态结构对环境变化的影响与适应[J].植物学通报,2005,22(增刊):118-127.
[18] 杨秉建,朱敏群,黄惠青,等.不同光照强度对叉花草叶片结构和叶绿素含量的影响[J].草原与草坪,2014(4):58-62.
[19] 张志良.植物生理学实验指导(第二版)[M].北京:高等教育出版社,1990.
[20] 白伟岚,任建武,苏雪痕.八种植物耐荫性比较研究[J].北京林业大学学报,1999,21(3):46-52.
[21] 张庆费,夏檑,钱又宇.城市绿化植物耐荫性的诊断指标体系[J].中国园林,2000,16(6):93-95.
[22] 刘艳玲,倪学明.3种野生耐荫地被植物的调查与评价[J].草业科学,2004,21(9):77-79.
[23] 冯建灿,张玉沽.喜树光合速率日变化及其影响因子的研究[J].林业科学,2002,38(4):3-39.
[24] 孙磊,章铁,李宏,等.柿树光合速率日变化及其影响因子的研究[J].北方果树,2006(3):4-6.
[25] 朱万泽,王金锡,薛建辉.台湾桤木引种的光合生理特性研究[J].西北植物学报,2004,24(11):2012-2019.
[26] 张振英,段朋娜,陈昕.遮阴对石灰花楸幼苗生长和光合特性的的影响[J].甘肃农业大学学报,2015,49(6):138-143.
[27] 冷平生,杨晓红.5种园林树木的光合和蒸腾特性的研究[J].北京农学院学报,2000,15(4):13-18.
[28] 李炎林,桂克印,唐前瑞,等.地被植物耐荫性研究进展[J].现代园艺,2008(5):4-5.
猜你喜欢
东坡赤壁诗词(2023年1期)2023-05-30 13:07:54
建筑与预算(2022年6期)2022-07-08 13:59:18
节水灌溉(2021年5期)2021-05-26 06:49:38
中国农业科技导报(2020年12期)2020-03-15 10:50:08
中成药(2019年12期)2020-01-04 02:02:44
疯狂英语·初中天地(2019年4期)2019-10-17 02:07:20
丝绸(2015年7期)2015-02-28 14:56:33
建筑与预算(2014年7期)2014-10-14 13:28:52
建筑与预算(2014年5期)2014-04-11 07:29:28
建筑与预算(2014年3期)2014-04-11 07:28:44
