
放牧对三江源区高寒草甸主要植物表型特征的影响
2015-12-21 03:25石红霄侯向阳师尚礼胡宁宁
草原与草坪 2015年3期
石红霄,侯向阳,师尚礼,贺 晶,胡宁宁
(1.甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070;2.中国农业科学院草原研究所,内蒙古 呼和浩特 010010)
放牧作为人类对草地生态系统最主要的干扰方式,对生态系统产生重要的影响。家畜可以通过选择性采食行为直接影响植物种群动态,并间接地改变了群落结构[1,2]。适度放牧可以增加群落的物种多样性,进而影响草地的生产力和稳定性;过度放牧则会导致群落多样性降低,草地生产力下降[3]。而植物的形态学指标是植物生长中能够直接观察的特征指标,能够直观地反映植物在环境中的生长变化。研究植物在放牧影响下表型特征变化也是研究放牧影响的重要内容。从功能性状变化探索草原植物的放牧响应机制,是受科学家青睐的思路[4]。目前,我国放牧对草地的影响主要集中于生物多样性[5]、生物量[6]、土壤养分变化[7]、植物形态结构[8]等影响的研究,关于长期放牧下草原植物性状可塑性变化的报道仍较少[9]。高寒草甸是青藏高原的主体,对维持生态系统功能有重要的作用[10],但由于长期过度放牧,草地生态系统退化、生产力持续衰减[11,12],这些现象已成为近年来我国生态学研究中的热点问题[13]。高原早熟禾(Poacrymophila)和矮嵩草(Kobresiakansuensis)是高寒草甸主要优势植物,研究其对放牧的表型反应对于揭示草原生态系统放牧响应机制具有很强的代表性。为此,以青藏高原典型植物矮嵩草和高原早熟禾为例,探讨自由放牧和围栏封育对植物表型特征影响,旨在揭示植物茎叶性状对放牧与围栏禁牧的差异化响应特征,为深入研究放牧对植株作用机理提供科学依据。
1 材料和方法
1.1 试验地概况
试验设于农业部玉树高寒草原资源与生态环境重点野外科学观测站,地理位置N 33°24′30″,E 97°18′00″,海拔4 250m。年平均温度为-6.4~4.3℃,7月平均最高温度为11.7~21.0℃,1月平均最低气温为-27.9~14.3℃。全年可照时数4 425h,日照时数2 488~2 619h。年均降水量为374.2~721.2mm,分配不均匀,降水日数130~160d,多年一日最大降水量36.5mm。蒸发量1 430~1 615mm,属青藏高原气候系统。
1.2 试验样地
试验选取的草地类型为高山嵩草杂草类草甸,草地主要优势牧草是高山嵩草(Kobresiapygmea),其次为矮嵩草、异针茅(Stipaaliena)、高原早熟禾、垂穗披碱草(Elymusnatans)、丁柱委陵菜(Potehtillasaundersiana)、乳白香青(Anaphalislactea)、高山唐松草(Thalictrumalpinum)等[14]。
1.2.1 样地设置 观测试验站佳塘草原为一块平坦开阔的退化高寒嵩草草甸,面积20hm2,近年来,采取了围栏封育和自由放牧的利用和管理措施。试验分别设置了围栏封育样地和自由放牧2个处理。
(1)围栏封育样地:随机选择围封3,5和10年的高寒草甸样地3个,做为围栏封育样地。样地内植被盖度在80%,高度20cm,地表有枯落物覆盖层。
(2)自由放牧退化样地:围栏外一直处于自由放牧状态,为比较典型的退化高寒草甸利用方式,围栏封育样地周边随机选择3个自由放牧草地,做为自由放牧样地。植被盖度约40%,草层高度在8cm以下,地表无枯落物覆盖。
1.3 试验方法
于2013年7月中旬在围栏禁牧样地和自由放牧6个样地,均随机设置3个1m×1m的样方,每个样方随机选取6株高原早熟禾、矮嵩草,测定株高、叶片数、叶面积、茎粗、主根长和主根粗。
株高的测定采用直尺直接测定每个植株地上部分的自然高度,取其均值;测定记录每个植株现存的绿色叶片的数量,取其均值;叶面积的测定采用游标卡尺测定每个植株的叶长、叶宽(叶宽是指叶片的中间部分的宽度),然后将二者的乘积作为叶面积,取均值;茎粗、主根长、主根粗的测定,随机选取植株,然后将其连根挖出,用游标卡尺测量茎粗、主根长、主根粗的值,取均值。
1.4 数据分析
采用Excel 2007软件对数据进行整理和绘图,应用SAS 9.1软件进行方差分析。
2 结果与分析
2.1 放牧对植株高度的影响
高原早熟禾和矮嵩草的株高在放牧的影响下均有所减小,植株高度减小的程度因草种不同而有所差异,且与围栏禁牧差异显著(P<0.05)。高原早熟禾在围栏禁牧条件下植株高度为12.2cm,在自由放牧条件下为4.3cm,比围栏禁牧的植株高度减少64.62%。矮嵩草在围栏禁牧条件下植株高度为5.19cm,在自由放牧条件下为2.10cm,比围栏禁牧的植株高度减少61.66%(图1)。表明放牧抑制了高原早熟禾和矮嵩草植株的生长高度。

图1 放牧下的植株高度Fig.1 Effects of grazing on plant height
2.2 放牧对植株叶片数的影响
高原早熟禾和矮嵩草的叶片数都受到放牧影响,有一定程度的下降(图2),其中,放牧与围栏禁牧条件下高原早熟禾植株叶片数差异不显著(P>0.05),矮嵩草植株叶片数差异显著(P<0.05)。高原早熟禾在围栏禁牧条件下植株叶片数为2.5个,在自由放牧条件下为2.28个,比围栏禁牧少10.87%。矮嵩草在围栏禁牧条件下植株叶片数为3.67个,在自由放牧条件下为2.28个,比围栏禁牧条件下减少了37.88%(图2)。表明放牧在一定程度上抑制了高原早熟禾和矮嵩草叶片数量的增加。

图2 放牧下植株的叶片数Fig.2 Effects of grazing on leaf number
2.3 放牧对植株叶面积的影响
高原早熟禾和矮嵩草的植株叶面积在放牧的影响下都有所变化,变化的程度因草种的不同也有所不同(图3),放牧对高原早熟禾和矮嵩草植株叶面积的影响均极显著(P<0.05)。高原早熟禾在围栏禁牧条件下的叶面积为64.60mm2,同时自由放牧条件下为26.40mm2,比围栏禁牧条件下的叶面积减少了59.14%。矮嵩草在围栏禁牧条件下的叶面积为17.62 mm2,在自用放牧条件下为7.47mm2,比围栏禁牧条件下的叶面积减少了57.56%。由此可见,放牧在一定程度上抑制了高原早熟禾和矮嵩草叶面积的增大。

图3 放牧下植株的叶面积Fig.3 Effects of grazing on leaf area
2.4 放牧对植株茎粗的影响
放牧条件下高原早熟禾和矮嵩草植株茎粗比围栏禁牧的减小(图4),其中,高原早熟禾茎粗的差异显著(P<0.05),矮嵩草茎粗不显著(P>0.05)。高原早熟禾在围栏禁牧条件下茎粗为0.69mm,在自由放牧为0.52mm,比围栏禁牧条件下减少了24.76%。矮嵩草围栏禁牧下茎粗为0.52mm,自由放牧下为0.47mm,比围栏禁牧条件下减少了9.98%。表明放牧在一定程度上抑制了高原早熟禾和矮嵩草的茎粗。
2.5 放牧对植株主根长的影响

图4 放牧下植株的茎粗Fig.4 Effects of grazing on stem diameter

图5 放牧下植株的主根长Fig.5 Effects of grazing on length of primary root
放牧条件下高原早熟禾和矮嵩草主根长较围栏禁牧下有所降低(图5),但差异均不显著(P>0.05)。高原早熟禾在围栏禁牧条件下植株主根长为48.25mm,自由放牧为42.44mm,比围栏禁牧减少12.03%。矮嵩草在围栏禁牧条件下植株主根长为36.96mm,在自由放牧条件下为30.89mm,比围栏禁牧条件下减少了16.43%。表明放牧在一定程度上抑制了高原早熟禾和矮嵩草的主根长。
2.6 放牧对植株主根粗的影响
高原早熟禾和矮嵩草在放牧下植株主根粗比不放牧的都小(图6),其中,放牧对高原早熟禾植株主根粗的影响显著(P<0.05),对矮嵩草植株主根粗的影响不显著(P>0.05)。高原早熟禾围栏禁牧植株主根粗为0.513mm,在自由放牧下为0.34mm,比围栏禁牧减少34.17%。矮嵩草在围栏禁牧下主根粗为0.48mm,自由放牧为0.39mm,比围栏禁牧减少17.07%。表明放牧在一定程度上抑制了高原早熟禾和矮嵩草茎的主根粗度增加。
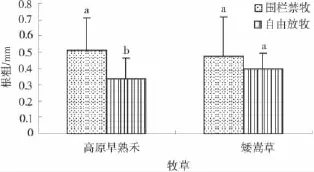
图6 放牧下植株的主根长Fig.6 Effects of grazing on length primary root
3 讨论与结论
3.1 放牧对植物表型特征的影响
过度放牧下植物茎叶性状功能发生变化,是植物对过度放牧的适应性形成的避牧机制[15]。研究结果表明[16,17],叶片形态对环境的响应和适应特征直接影响着植物与周围环境的物质和能量交换、植物的生存能力和发展、变化方向。此次研究证实,与围栏禁牧相比较,放牧抑制了高原早熟禾和矮嵩草的生长,导致了2种植物株高、叶面积的显著下降(P<0.05),植物个体表现出明显的矮小化现象,这与周寿荣等[18]高载畜率(放牧率)、高放牧强度下,放牧草地草丛矮小的结论相一致。而长期放牧后,短期围封对植物叶片数、根长、根粗等性状的恢复效果并不显著(P>0.05),这与王炜等[19]报道的放牧退化草原植物性状具有保守性的结论一致。植物随着放牧胁迫增强,往往先采取高度和生物量降低的适应策略[20]。因此,植物的矮化型变是草原生态系统结构与功能变化的重要触发机制。
高原早熟禾在放牧下茎粗显著下降,而矮嵩草的茎粗也出现了一定程度的减少,但减少的不显著;高原早熟禾的叶片数下降不显著,而矮嵩草的下降较为显著。这与董世魁等[21]的试验结果相符。放牧对不同的植物有不同的影响,可能与植物自身的适应能力以及家畜的选择性采食有关。根系是植物体受放牧活动机械干扰(如采食)较小的部位。高原早熟禾在放牧的影响下主根长的下降不显著,主根粗的下降比较明显;矮嵩草的主根长和主根粗都下降不明显。与放牧会使牧草根系总量减少的结论相符[22]。根量减少会对土壤生态过程产生消极作用,同时吸收矿质营养的面积减少,也会抑制地上部分生长[23]。
3.2 放牧对植物表型特征影响的可能机制
试验仅从植物形态学的6个指标研究放牧对植物形态学的影响,比较了围栏禁牧和放牧条件下植物表形变化,植物所能承受的最佳放牧压以及植物发生变化机理的确定还需要进行不同放牧压下植物形态学以及微观形态等方面的研究。在未来研究中,解析放牧干扰下植物型变机理,不仅应考虑种群、群落、生态系统、景观等宏观尺度,而且应更侧重于分子生态学机理。分子生物学理论与技术近年来得到迅猛发展,它以巨大的解释能力,为探究放牧下草原植物矮化型变生态现象背后的机理提供了新的方法,展现了诱人前景。目前,草原植物分子生态学研究才刚刚起步[24],未来需利用基因组学、转录组学、蛋白组学、代谢组学,以及表型组学的高通量分析技术,发现从放牧等诱导信号到植物代谢调控的过程,解析环境应答基因,从而构建起草原植物矮化型变的信号调控途径与代谢网络。
[1] Diaz S,Lavorel S,Mcintyre S,et al.Plant trait responses to grazing-aglobal synthesis[J].Global Change Biol,2006(12):1-29.
[2] 付伟,赵俊权,杜国祯.青藏高原高寒草地放牧生态系统可持续发展研究[J].草原与草坪,2013,33(1):84-88.
[3] 袁建立,江小蕾,黄文冰,等.放牧季节及放牧强度对高寒草地植物多样性的影响[J].草业学报,2004,13(3):16-21.
[4] Louault F,Pillar V D,Aufrère J,et al.Plant traits and functional types in response to reduceddisturbance in a semi-natural grassland[J].Journal of Vegetation Science,2005,16:151-160.
[5] Wu G L,Du G Z,Liu Z H,et al.Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau[J].Plant Soil,2009,319(1):115-126.
[6] 仁青吉,武高林,任国华.放牧强度对青藏高原东部高寒草甸植物群落特征的影响[J].草业学报,2009,18(5):256-261.
[7] Sun D S,Wesche K,Chen D D,et al.Grazing depresses soil carbon storage through changing plant biomass and composition in a Tibetan alpine meadow[J].Plant Soil Environ,2011,57(6):271-278.
[8] 于向芝.过度放牧对内蒙古典型草原11种植物茎叶结构的影响[D].呼和浩特:内蒙古农业大学,2007.
[9] Stahlheber K A,D’Antonio C M.Using livestock to manage plant composition:A meta-analysis of grazing in California Mediterranean grasslands[J].Biological Conservation,2013,157:300-308.
[10] 王宝山,尕玛加,张玉.青藏高原“黑土滩”退化高寒草甸草原的形成机制和治理方法的研究进展[J].草原与草坪,2007,2:72-77.
[11] Li W J,Ali S H,Zhang Q.Property rights and grassland degradation:a study of the Xilingol Pasture,Inner Mongolia,China[J].Journal of Environmental Management,2007,85:461-470.
[12] 侯向阳.中国草原科学[M].北京:科学出版社,2013:424-470.
[13] Li S Y,Verburg P H,LüS H,et al.Spatial analysis of the driving factors of grassland degradation under conditions of climate change and intensive use inInner Mongolia,China[J].Regional Environmental Change,2012,12,:461-474.
[14] 范月君,侯向阳,石红霄,等.封育与放牧对三江源区高寒草甸植物和土壤碳储量的影响[J].草原与草坪,2012,32(5),41-46)
[15] Suzuki R O,Suzuki S N.Facilitative and competitive effects of a large species with defensive traits on a grazing-adapted,small species in a long-term deer grazinghabitat[J].Plant Ecology,2011,212:343-351.
[16] Schuepp P H.Tansley review No.59.Leaf boundary layers[J].New Phytologist,1993,125:477-507.
[17] Yu G R,Wang Q F.Ecophysiology of Plant Phytosynthesis,Transpiration,and Water Use[M].Beijing:Science Press,2010.
[18] 周寿荣.草地生态学[M].北京:中国农业出版社,1998.
[19] 王炜,梁存柱,刘钟龄,等.草原群落退化与恢复演替中的植物个体行为分析[J].植物生态学报,2000,24:268-274.
[20] 汪诗平,王艳芬,陈佐忠.放牧生态系统管理[M].北京:科学出版社,2003:113-132.
[21] 董世魁,丁路明,徐敏云,等.放牧强度对高寒地区多年生混播禾草叶片特征及草地初级生产力的影响[J].中国农业科学,2004,37(1):136-142.
[22] Ford M A,Grace J B.Effects of vertebrate herbivores on soil processes,plant biom ass,litter a c cum ulat ion and soil elevation changes in a coastal m arsh[J].Journal of Ecology,1998,86(6):974-982.
[23] 侯扶江,杨中艺.放牧对草地的作用[J].生态学报,2006,26(1):244-264.
[24] 韩冰,赵萌莉,珊丹.针茅属植物分子生态学[M].北京:科学出版社,2011:1-32.
猜你喜欢
农业环境科学学报(2022年9期)2022-10-14
广西植物(2022年5期)2022-06-18
文山学院学报(2022年2期)2022-05-27
课外生活·趣知识(2022年5期)2022-05-14
今日农业(2021年2期)2021-11-27
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
红蜻蜓·低年级(2020年12期)2020-06-20
新疆农业科学(2020年1期)2020-02-14
好孩子画报(2019年8期)2019-09-19
发明与创新·大科技(2018年2期)2018-03-17
