
OXA-23型酶介导的鲍曼不动杆菌对碳青霉烯类抗生素耐药分子传播机制
2015-12-19 10:36程巧菊许苔希
浙江临床医学 2015年6期
程巧菊 许苔希
·实验研究·
OXA-23型酶介导的鲍曼不动杆菌对碳青霉烯类抗生素耐药分子传播机制
程巧菊 许苔希
目的 探讨OXA-23酶在介导鲍曼不动杆菌碳青霉烯类抗生素耐药过程中的传播机制。方法 对2010年9月至2012年3月58株产OXA-23型碳青霉烯酶鲍曼不动杆菌进行深入研究,所有菌株运用PCR mapping方法对blaOXA-23基因的周围结构进行扩增及测序分析;根据PFGE型别挑选不同克隆的菌株,进一步采用S1-PFGE以及ApaI-PFGE联合Southern blot杂交法对blaOXA-23基因进行定位分析。结果 58株菌中有47株菌的blaOXA-23基因周围结构为Tn2009转座结构、11株为Tn2008转座结构;58株菌属于8个不同的克隆型别(A、B、C、D、E、F1、F2、G和H克隆),其中E克隆的blaOXA-23基因存在2个拷贝,分别位于质粒及染色体上,质粒大小为78.2kb左右;其余克隆菌blaOXA-23基因定位于染色体上,所在片段大小为310.1kb左右,其中D克隆存在2个染色体拷贝。结论 转座结构是导致blaOXA-23基因迅速在鲍曼不动杆菌中转移的重要元件,导致鲍曼不动杆菌碳青霉烯类抗生素耐药传播和扩散。
OXA-23酶 碳青霉烯类 耐药传播 鲍曼不动杆菌
近年来,我国鲍曼不动杆菌临床分离率逐年升高[1~3]。多重耐药鲍曼不动杆菌(MDRAB)菌株和碳青霉烯类耐药鲍曼不动杆菌(CRAB)已呈世界性流行,在我国现已成为院内感染的重要病原菌[4]。Ambler D类碳青霉烯酶中的D组丝氨酸苯唑西林酶(OXA)是介导鲍曼不动杆菌成为CRAB的主要碳青霉烯酶,鲍曼不动杆菌中>90%的碳青霉烯酶基因为blaOXA基因,根据氨基酸序列以及酶对碳青霉烯类抗生素的水解活性[5]。OXA酶中具备水解碳青霉烯类能力的D类酶称为CHDLs,包括blaOXA-23、blaOXA-24、blaOXA-58等[5]。CRAB最主要的耐药机制就是获得了blaOXA-23基因[6]。然而,目前有关该基因的研究仍处于探索阶段。本实验拟对发现的产OXA-23酶鲍曼不动杆菌进行深入研究,探讨blaOXA-23基因的分子流行病学传播规律。报道如下。
1 材料和方法
1.1 材料 本实验菌株来源于2010年9月至2012年3月收集自浙江省临海市中医院和浙江省台州医院分离的60株 CRAB,经聚合酶链式反应(PCR)扩增blaOXA-23基因,明确为blaOXA-23基因阳性的菌株纳入本实验,共计58株CRAB[7,8]。
1.2 主要仪器及试剂 PCR扩增仪为ABI Applied公司产品;CHEF Mapper XA型脉冲场凝胶电泳仪、电泳仪及凝胶成像仪均为美国Bio-rad公司产品;恒温水平摇床为美国Thermo公司产品。PCR试剂盒购自TaKaRa公司。引物由上海生工生物工程有限公司合成;PCR产物测序由杭州铂尚测序公司完成,测序仪为美国ABI公司产品ABI Prism 3730。
1.3 blaOXA-23周围结构的研究 运用PCR mapping技术,根据文献报道blaOXA-23周围结构的序列特点设计用于扩增Tn2006、Tn2008及Tn2009的PCR mapping引物。PCR扩增条件:95℃,5min;95℃解链45s,60℃退火30s,72℃延伸1.5min,35个循环;72℃,10min。扩增产物经电泳鉴定是否有目的条带以及条带的特异性。引物的相对位置见图1,引物序列见表1。
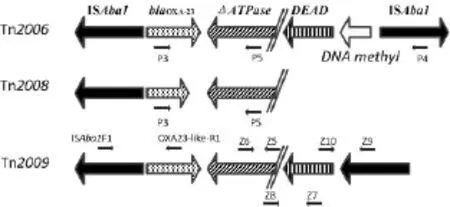
图1 blaOXA-23基因周围转座结构及引物相对位置

表1 blaOXA-23周围结构引物列表
1.4 菌株的同源性分析 根据前期PFGE实验结果,对58株菌重新进行ApaI-PFGE分析,明确菌株的同源性以便挑选菌株进行blaOXA-23基因的定位分析[9]。Apa-PFGE分析酶切条件为运用ApaI核酸内切酶37℃处理4~6h,在脉冲场凝胶电泳仪上进行电泳,PFGE条件:0.5×TBE、14℃、电压6V/cm、电场夹角120°、电流转换时间5~20s、电泳22h。电泳结束后采用GelRed染料对胶进行染色,并用凝胶成像仪拍摄结果。以肠炎沙门菌H9812菌株经Xba I酶切片段作为分子量标准。
1.5 blaOXA-23基因定位分析 挑选PFGE型别不同的菌株进行blaOXA-23基因定位实验。制备包埋有细菌基因组的小胶,以S1核酸内切酶对细菌基因组进行酶切,37℃处理40min;在脉冲场凝胶电泳仪上进行电泳,PFGE条件:0.5×TBE、14℃、电压6V/cm、电场夹角120°、电流转换时间2.13~63.6s、电泳18h。以blaOXA-23基因为探针进行Southern blot杂交试验,将PFGE胶经DNA脱嘌呤及变性处理,利用虹吸原理将胶上DNA转移至硝酸纤维素膜上,以地高辛标记的单链blaOXA-23基因DNA作为探针与膜上DNA进行杂交,明确该基因是否定位于质粒上[10]。对于S1-PFGE定位实验失败的细菌进一步进行ApaI-PFGE电泳后,blaOXA-23基因Southern blot杂交试验,确定blaOXA-23基因是否定位于染色体上。
2 结果
2.1 blaOXA-23基因周围结构结果 PCR mapping及测序比对结果显示58株blaOXA-23阳性菌株中有11株blaOXA-23周围的转座结构为Tn2008,47株为Tn2009转座结构。见表2。
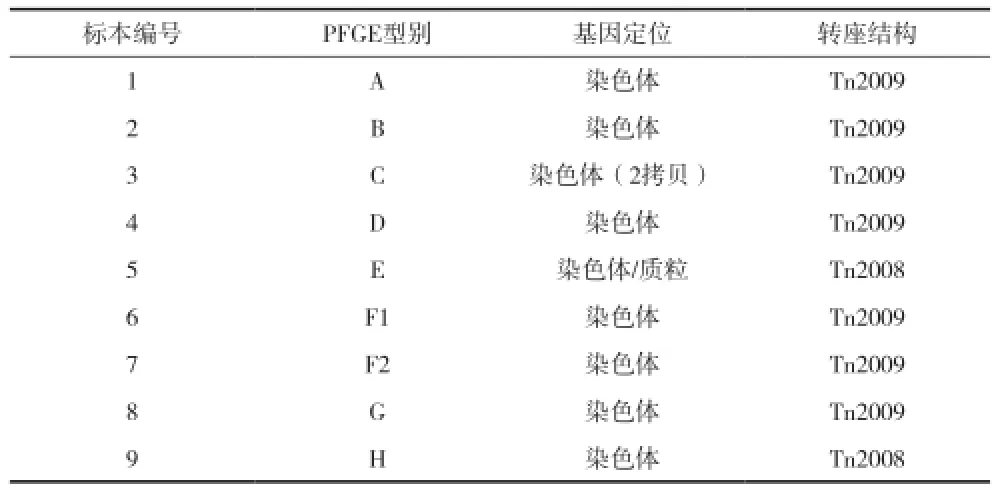
表2 blaOXA-23定位分析结果
2.2 菌株同源性分析结果 PFGE结果显示入选菌株的PFGE型别来源于8个克隆,分别为A、B、C、D、E、F1、F2、G和H克隆,见图2。
2.3 blaOXA-23基因定位分析结果 对不同克隆型的细菌进行blaOXA-23基因的Southern blotting杂交定位试验,结果显示:9种型别的菌株仅有1株存在一个拷贝blaOXA-23基因定位于质粒上,质粒大小为78.2kb左右(图3),该菌同时在染色体上亦存在一个拷贝;9株菌株blaOXA-23基因均存在定位于染色体上的拷贝,其中3号菌存在两个大小不同的染色体拷贝(图4)。
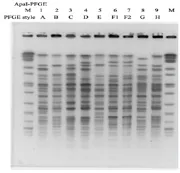
图2 菌株同源性分析结果
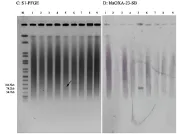
图3 菌株blaOXA-23基因质粒杂交定位结果
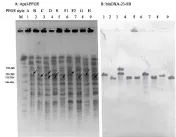
图4 菌株blaOXA-23基因染色体杂交定位结果
3 讨论
近年来,CRAB呈全球流行,临床有效抗菌药物的效果较局限,已经成为住院患者的严重威胁[11]。鲍曼不动杆菌发展成为CRAB的主要机制是细菌通过水平转移获得具有编码碳青霉烯类抗生素水解活性酶的基因,其中OXA-23碳青霉烯酶为鲍曼不动杆菌中的最主要的D类碳青霉烯酶。本文通过对收集的58株blaOXA-23基因的周围结构以及基因的定位进行研究,发现Tn2009转座结构以及质粒广泛存在于CRAB菌株中,揭示了CRAB流行及播散的规律。
碳青霉烯类抗生素是目前公认的对革兰阴性菌具有良好抗菌活性的药物,CRAB的出现及流行无疑给临床治疗带来了严重困难,亦成为院感防控的难点。OXA-23-likeβ-内酰胺酶(OXA-23 β-lactamase)是一类具有碳青霉烯类抗生素水解活性的β-内酰胺酶,包括OXA-23,OXA-27等19种不同的亚型;该酶于1985年首次在英国被报道,由鲍曼不动杆菌产生;产酶菌株对亚胺培南的MICs值高达16μg/ml,直接与耐药相关。研究显示blaOXA-23基因广泛为不动杆菌属的多种细菌携带,亦存在于不同遗传背景的鲍曼不动杆菌中,是CRAB产生最重要原因之一。blaOXA-23基因定位于质粒上,且能随质粒在不同菌种间传递,这种携带blaOXA-23基因的可转移性质粒是导致CRAB在医院范围内的广泛流行重要危险因素[12]。研究显示blaOXA-23基因的周围结构具有相似性,2011年我国学者在49株鲍曼不动杆菌中检测到blaOXA-23基因周围稳定的结构—Tn2008,该结构由ISAba1-blaOXA-23-ΔATPase三个基因组成,并通过对ISAba1周围序列进行分析提出ISAba1具有介导blaOXA-23基因转移的能力的观点[13]。研究显示pAZJ221质粒以及Tn2009转座结构是导致我国blaOXA-23基因广泛存在于不动杆菌属细菌,导致CRAB产生的主要原因,从而进一步从基因水平揭示CRAB流行的机制[10]。但本资料结果显示质粒并非承载blaOXA-23基因的主要移动元件,作者仅发现一个克隆型的细菌携带有blaOXA-23质粒(大小为78.3kb);亦发现Tn2009转座结构及Tn2008转座结果同时是blaOXA-23基因的重要携带者,较有可能参与该基因在鲍曼不动杆菌中的转移,是CRAB产生的重要威胁。ST92型鲍曼不动杆菌是全球流行的CRAB型别,而克隆播散是导致该ST型别流行的主要原因之一,本结果进一步证实除克隆播散以外,基于转座结构的blaOXA-23基因的转移元件亦参与CRAB的产生及播散,揭示了CRAB医院感染(尤其是对于ICU患者)流行及扩散的规律。
1 汪复, 朱德妹, 胡付品,等. 2008年中国CHINET细菌耐药性监测. 中国感染与化疗杂志,2009,9(5): 321~329.
2 汪复. 2006年中国CHINET细菌耐药性监测. 中国感染与化疗杂志, 2008, 8(1): 1~9.
3 朱德妹, 汪复, 胡付品. 2009年中国CHINET细菌耐药监测结果. 中国感染与化疗杂志 ,2010,10(5): 325~34.
4 胡付平,朱德妹,汪复,等. 2013年中国CHINET细菌耐药性监测. 中国感染与化疗杂志, 2014,14(5): 365~374.
5 Evans BA, Amyes SG. OXA beta-lactamases. Clinical microbiology reviews,2014,27(2): 241~263.
6 Zhou H, Pi BR, Yang Q,et al. Dissemination of imipenem-resistant Acinetobacter baumannⅡ strains carrying the ISAba1 blaOXA-23 genes in a Chinese hospital. J Med Microbiol,2007,56(8): 1076~1080.
7 余素飞,厉世笑,黄林瑶,等.碳青霉烯类耐药鲍曼不动杆菌的耐药性及碳青霉烯酶基因型研究.中华临床感染病杂志,2013,6(2): 108~110.
8 厉世笑 余素飞, 黄林瑶. 耐碳青霉烯类鲍曼不动杆菌OXA基因检测及同源性分析. 中国微生态学杂志,2013,25(3): 332~334.
9 Lee MH, Chen TL, Lee YT,et al. Dissemination of multidrug-resistant Acinetobacter baumannⅡ carrying BlaOXA-23 from hospitals in central Taiwan. Journal of microbiology, immunology and infection=Wei mian yu gan ran za zhi,2013,46(6): 419~424.
10 Liu LL, Ji SJ, Ruan Z,et al. Dissemination of blaOXA-23 in Acinetobacter spp. in China: main roles of conjugative plasmid pAZJ221 and transposon Tn2009. Antimicrobial agents and chemotherapy,2015,59(4): 1998~2005.
11 Pogue JM, Mann T, Barber KE, et al. Carbapenem-resistant Acinetobacter baumannⅡ: epidemiology, surveillance and management. Expert review of anti-infective therapy,2013,11(4): 383~393.
12 Poirel L, Nordmann P. Carbapenem resistance in Acinetobacter baumannⅡ: mechanisms and epidemiology. Clinical microbiology and infection:the official publication of the European Society of Clinical Microbiology and Infectious Diseases,2006,12(9): 826~836.
13 Wang X, Zong Z, Lu X. Tn2008 is a major vehicle carrying bla(OXA-23) in Acinetobacter baumannⅡ from China. Diagnostic microbiology and infectious disease,2011,69(2): 218~222.
Objective To illustrate the molecular transmission mechanism of carbapenem-resistance mediated by OXA-23 type carbapenemase in A. baumannⅡ. Methods Fifty-eight strains collected from September 2010 to March 2012 were involved in our study. PCR mapping method followed by sequencing was used to analysis the surrounding structure of the blaOXA-23gene. Those isolates originating from different clones according to PFGE analysis were picked out to detect the gene location of the blaOXA-23with S1-PFGE and ApaI-PFGE followed by Southern blotting method. Results 47 in 58 strains harbored the blaOXA-23gene in Tn2009 transposon structure,while 11 in Tn2008. Only one type of PFGE clone strain (Clone E) harbors the blaOXA-23gene both in plasmid (ranging around 78.2kb) and in chromosome. The blaOXA-23gene was located in chromosome in the rest types of clones,in which two copies of clones were found in Clone D. Conclusion Both transposon was the main factors mediated the blaOXA-23gene transmission among A. baumannⅡ,which was also the molecular basis leading to the epidemic of carbapenem-resistant A. baumannⅡ.
OXA-23 type carbapenemase Carbapenems Resistant transmission A. baumannⅡ
317000 浙江省临海市中医院检验科(程巧菊)
317000 浙江省台州医院检验科(许苔希)
猜你喜欢
传染病信息(2022年2期)2022-07-15
现代临床医学(2022年3期)2022-06-06
天津医科大学学报(2019年6期)2019-08-13
天津医科大学学报(2019年3期)2019-08-13
人人健康(2019年21期)2019-01-12
幼儿园(2016年4期)2016-03-14
